
Dynamics of Cardicola spp. Infection in Ranched Southern Bluefin Tuna: First Observation of C. orientalis at Transfer

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Sample Analysis
2.3. Statistical Analysis
3. Results
3.1. 2021 Season
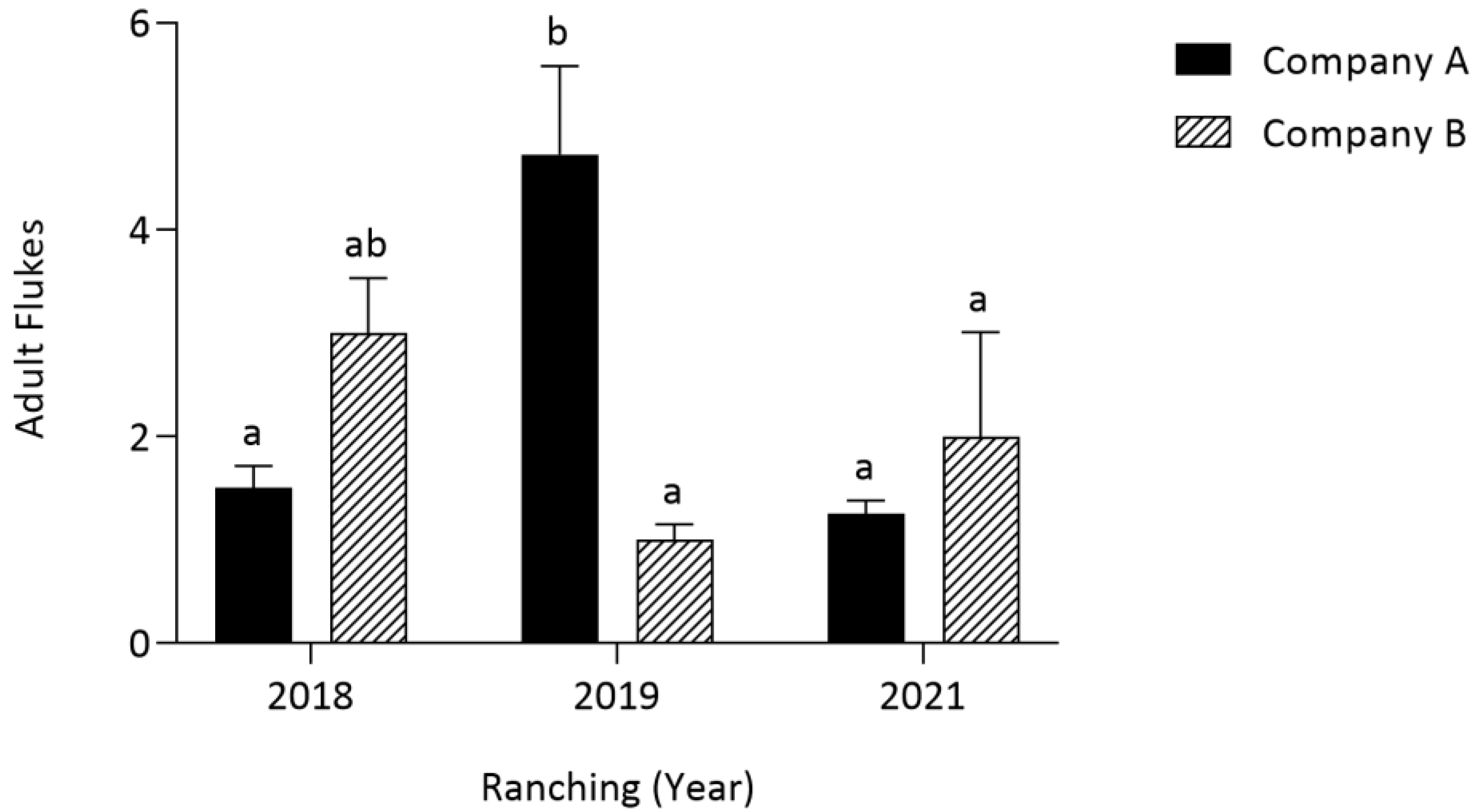
3.2. Comparison of Harvest Year
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, D.; Kiessling, I. Ranching of Southern bluefin tuna in Australia. In Advances in Tuna Aquaculture: From Hatchery to Market; Elsevier: Oxford, UK, 2016; pp. 217–232. [Google Scholar] [CrossRef]
- Patterson, T.A.; Eveson, J.P.; Hartog, J.R.; Evans, K.; Cooper, S.; Lansdell, M.; Hobday, A.J.; Davies, C.R. Migration Dynamics of Juvenile Southern Bluefin Tuna. Sci. Rep. 2018, 8, 14553. [Google Scholar] [CrossRef]
- South Australian Aquaculture: A Summary of Its Diversity, Production and Innovation. Available online: https://www.pir.sa.gov.au/__data/assets/pdf_file/0008/221993/Aquaculture_IndustryBooklet.pdf (accessed on 27 June 2023).
- The Economic Contribution of Aquaculture in the South Australian State and Regional Economies, 2020/21: A Report for PIRSA Fisheries and Aquaculture. Available online: https://www.pir.sa.gov.au/__data/assets/pdf_file/0006/434067/economic-contribution-aquaculture-2021-22.pdf (accessed on 27 June 2023).
- Polinski, M.; Hamilton, D.B.; Nowak, B.F.; Bridle, A.R. SYBR, TaqMan, or both: Highly sensitive, non-invasive detection of Cardicola blood fluke species in Southern Bluefin Tuna (Thunnus maccoyii). Mol. Biochem. Parasitol. 2013, 191, 7–15. [Google Scholar] [CrossRef]
- Shirakashi, S.; Tsunemoto, K.; Webber, C.; Rough, K.; Ellis, D.; Ogawa, K. Two species of Cardicola (Trematoda: Aporocotylidae) found in Southern Bluefin Tuna Thunnus maccoyii ranched in South Australia. Fish Pathol. 2013, 48, 1–4. [Google Scholar] [CrossRef]
- Neumann, L.; Bridle, A.R.; Leef, M.J.; Nowak, B.F. Annual variability of infection with Cardicola forsteri and Cardicola orientalis in ranched and wild Southern Bluefin Tuna (Thunnus maccoyii). Aquaculture 2018, 487, 1–6. [Google Scholar] [CrossRef]
- Power, C.; Nowak, B.F.; Cribb, T.H.; Bott, N.J. Bloody flukes: A review of Aporocotylids as parasites of cultured marine fishes. Int. J. Parasitol. 2020, 50, 743–753. [Google Scholar] [CrossRef]
- Power, C.; Evenden, S.; Rough, K.; Webber, C.; Widdicombe, M.; Nowak, B.F.; Bott, N.J. Prevalence and intensity of Cardicola spp. infection in ranched Southern Bluefin Tuna and a comparison of diagnostic methods. Pathogens 2021, 10, 1248. [Google Scholar] [CrossRef]
- Colquitt, S.E.; Munday, B.L.; Daintith, M. Pathological findings in Southern Bluefin Tuna, Thunnus maccoyii (Castelnau). infected with Cardicola forsteri (Cribb, Daintith & Munday, 2000) (Digenea: Sanguinicolidae), a Blood Fluke. J. Fish Dis. 2001, 24, 225–229. [Google Scholar] [CrossRef]
- Dennis, M.M.; Landos, M.; Antignana, D.T. Case-control study of epidemic mortality and Cardicola forsteri–Associated disease in farmed Southern Bluefin Tuna (Thunnus maccoyii) of South Australia. Vet. Pathol. 2011, 48, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Tanaka, S.; Sugihara, Y.; Takami, I. A new blood fluke of the genus Cardicola (Trematoda: Sanguinicolidae) from Pacific Bluefin Tuna Thunnus orientalis (Temminck & Schlegel, 1844) cultured in Japan. Parasitol. Int. 2010, 59, 44–48. [Google Scholar] [CrossRef]
- Shirakashi, S.; Kishimoto, Y.; Kinami, R.; Katano, H.; Ishimaru, K.; Murata, O.; Itoh, N.; Ogawa, K. Morphology and distribution of blood fluke eggs and associated pathology in the gills of cultured Pacific Bluefin Tuna, Thunnus orientalis. Parasitol. Int. 2012, 61, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Cribb, T.H.; Chick, R.C.; O’Connor, W.; O’Connor, S.; Johnson, D.; Sewell, K.B.; Cutmore, S.C. Evidence that blood flukes (Trematoda: Aporocotylidae) of chondrichthyans infect bivalves as intermediate hosts: Indications of an ancient diversification of the Schistosomatoidea. Int. J. Parasitol. 2017, 47, 885–891. [Google Scholar] [CrossRef] [PubMed]
- de Ybáñez, R.R.; Peñalver, J.; Martínez-Carrasco Pleite, C.; Del Rio, L.; Dolores, E.; Berriatua, E.; Muñoz, P. Blood fluke infection (Digenea: Aporocotylidae) of cage reared Atlantic Bluefin Tuna Thunnus thynnus (Scombridae) in west Mediterranean. Fish Pathol. 2011, 46, 87–90. [Google Scholar] [CrossRef]
- Cribb, T.H.; Adlard, R.D.; Hayward, C.J.; Bott, N.J.; Ellis, D.; Evans, D.; Nowak, B.F. The Life Cycle of Cardicola forsteri (Trematoda: Aporocotylidae), a pathogen of ranched Southern Bluefin Tuna, Thunnus maccoyii. Int. J. Parasitol. 2011, 41, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Cribb, T.H.; Daintith, M.; Munday, B.L. A New Blood-Fluke, Cardicola forsteri (Digenea: Sanguinicolidae) of Southern Bluefin Tuna (Thunnus maccoyii) in aquaculture. Trans. R. Soc. South Aust. 2000, 124, 117–120. [Google Scholar] [CrossRef]
- Palacios-Abella, J.F.; Rodriguez-Lianos, J.; Mele, S.; Montero, F.E. Morphological Characterisation and Identification of Four Species of Cardicola Short, 1953 (Trematoda: Aporocotylidae) Infecting the Atlantic Bluefin Tuna Thunnus thynnus (L.) in the Mediterranean Sea. Syst. Parasitol. 2015, 91, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Ishimaru, K.; Shirakashi, S.; Takami, I.; Grabner, D. Cardicola opisthorchis n. sp. (Trematoda: Aporocotylidae) from the Pacific Bluefin Tuna, Thunnus orientalis (Temminck & Schlegel, 1844), cultured in Japan. Parasitol. Int. 2011, 60, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, N.T.; Leef, M.; Valdenegro, V.; Hayward, C.; Nowak, B.F. Correlation of humoral immune response in Southern Bluefin Tuna, T. maccoyii, with infection stage of the blood fluke, Cardicola forsteri. PLoS ONE 2012, 7, e45742. [Google Scholar] [CrossRef]
- Shirakashi, S.; Andrews, M.; Kishimoto, Y.; Ishimaru, K.; Okada, T.; Sawada, Y.; Ogawa, K. Oral treatment of praziquantel as an effective control measure against blood fluke infection in Pacific Bluefin Tuna (Thunnus orientalis). Aquaculture 2012, 326–329, 15–19. [Google Scholar] [CrossRef]
- Hardy-Smith, P.; Ellis, D.; Humphrey, J.; Evans, M.; Evans, D.; Rough, K.; Valdenegro, V.; Nowak, B.F. In vitro and in vivo efficacy of anthelmintic compounds against blood fluke (Cardicola forsteri). Aquaculture 2012, 334–337, 39–44. [Google Scholar] [CrossRef]
- Norbury, L.J.; Shirakashi, S.; Power, C.; Nowak, B.F.; Bott, N.J. Praziquantel use in aquaculture–Current status and emerging issues. Int. J. Parasitol. Drugs Drug Resist. 2022, 18, 87–102. [Google Scholar] [CrossRef]
- Permit to Allow Minor Use of an Unregistered Agvet Chemical Product for Control of Blood Fluke in Southern Bluefin Tuna. APVMA, Permit 85738. Available online: https://permits.apvma.gov.au/PER85738.PDF (accessed on 27 June 2023).
- Power, C.; Webber, C.; Rough, K.; Staunton, R.; Nowak, B.F.; Bott, N.J. The effect of different treatment strategies on Cardicola spp. (Trematoda: Aporocotylidae) infection in ranched Southern Bluefin Tuna (Thunnus maccoyii) from Port Lincoln, South Australia. Aquaculture 2019, 513, 734401. [Google Scholar] [CrossRef]
- Power, C.; Carabott, M.; Widdicombe, M.; Coff, L.; Rough, K.; Nowak, B.F.; Bott, N.J. Effects of company and season on blood fluke (Cardicola spp.) infection in ranched Southern Bluefin Tuna: Preliminary evidence infection has a negative effect on fish growth. Peer J 2023, 11, e15763. [Google Scholar] [CrossRef] [PubMed]
- Kirchhoff, N.T.; Rough, K.; Nowak, B.F. Moving cages further offshore: Effects on Southern Bluefin Tuna, T. maccoyii, parasites, health and performance. PLoS ONE 2011, 6, e23705. [Google Scholar] [CrossRef] [PubMed]
- Aiken, H.M.; Hayward, C.J.; Nowak, B.F. An epizootic and its decline of a blood fluke, Cardicola forsteri, in farmed Southern Bluefin Tuna, Thunnus maccoyii. Aquaculture 2006, 254, 40–45. [Google Scholar] [CrossRef]
- NanoDrop Lite: Interpretation of Nucleic Acid 260/280 Ratios, Thermo Scientific. Available online: https://assets.thermofisher.com/TFS-Assets/CAD/Product-Bulletins/T123-NanoDrop-Lite-Interpretation-of-Nucleic-Acid-260-280-Ratios.pdf (accessed on 27 June 2023).
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Underwood, A.J. Techniques of analysis of variance in experimental marine biology and ecology. Oceanogr. Mar. Biol. Annu. Rev. 1981, 19, 513–605. [Google Scholar]
- Cleophas, T.J.; Zwinderman, A.H. McNemar’s Tests. In Statistical Analysis of Clinical Data on a Pocket Calculator; Springer: Dordrecht, The Netherlands, 2011; pp. 39–40. [Google Scholar] [CrossRef]
- Aiken, H.; Hayward, C.; Cameron, A.; Nowak, B.F. Simulating blood fluke, Cardicola forsteri, infection in farmed Southern Bluefin Tuna, Thunnus maccoyii, using stochastic models. Aquaculture 2009, 293, 204–210. [Google Scholar] [CrossRef]
- Yong, R.Q.-Y.; Cutmore, S.C.; Miller, T.L.; Adlard, R.D.; Cribb, T.H. The ghost of parasites past: Eggs of the blood fluke Cardicola chaetodontis (Aporocotylidae) trapped in the heart and gills of butterflyfishes (Perciformes: Chaetodontidae) of the Great Barrier Reef. Parasitology. 2013, 140, 1186–1194. [Google Scholar] [CrossRef]
- Rohr, C.M.; Sprague, D.J.; Park, S.; Malcolm, N.J.; Marchant, J.S. Natural variation in the binding pocket of a parasitic flatworm TRPM channel resolves the basis for praziquantel sensitivity. Proc. Natl. Acad. Sci. USA 2023, 120, e2217732120. [Google Scholar] [CrossRef]
- Aiken, H.M.; Bott, N.J.; Mladineo, I.; Montero, F.E.; Nowak, B.F.; Hayward, C.J. Molecular evidence for cosmopolitan distribution of platyhelminth parasites of Tunas (Thunnus spp.). Fish Fish. 2007, 8, 167–180. [Google Scholar] [CrossRef]
- Ballí Garza, J. Identification of Parasitic Diseases Affecting ranched Southern Bluefin Tuna (SBT), Using Conventional and Molecular Methods. Ph.D. Thesis, University of Tasmania, Launceston, Australia, 2018. [Google Scholar]
- Saksvik, M.; Nilsen, F.; Nylud, A.; Berland, B. Effect of marine Eubothrium sp. (Cestoda: Pseudophyllidea) on the growth of Atlantic salmon, Salmo salar L. J. Fish Dis. 2001, 24, 111–119. [Google Scholar] [CrossRef]
- FHF (The Norwegian Fishery and Aquaculture Industry Research Fund). Bendelorm (Eubothrium sp.) hos laks (Salmo salar): Utprøving av nye Behandlingsmidler og Utvikling av et In Vitro Bioassay for Måling av Resistens Overfor Praziquantel–Rapport fra et Pilotprosjekt [Tapeworm (Eubothrium sp.) in salmon (Salmo salar): Trial of New Therapeutics and Development of an In Vitro Bioassay to Measure Praziquantel Resistance–Report]. Available online: https://www.fhf.no/prosjekter/prosjektbasen/551024/ (accessed on 24 November 2023).
- FHF (The Norwegian Fishery and Aquaculture Industry Research Fund). Resistens hos Bendelorm (Eubothrium sp.) Sluttrapport 2008 [Resistance in tapeworm (Eubothrium sp.) Final Report 2008]. Available online: https://www.fhf.no/prosjekter/prosjektbasen/552024/ (accessed on 24 November 2023).
- Shirakashi, S.; Tani, K.; Ishimaru, K.; Shin, S.P.; Honryo, T.; Uchida, H.; Ogawa, K. Discovery of intermediate hosts for two species of blood flukes Cardicola orientalis and Cardicola forsteri (Trematoda: Aporocotylidae) infecting Pacific Bluefin Tuna in Japan. Parasitol. Int. 2016, 65, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Shirakashi, S.; Tani, K.; Shin, S.P.; Ishimaru, K.; Honryo, T.; Sugihara, Y.; Uchida, H. Developmental stages of fish blood flukes, Cardicola forsteri and Cardicola opisthorchis (Trematoda: Aporocotylidae), in their polychaete intermediate hosts collected at Pacific Bluefin Tuna culture sites in Japan. Parasitol. Int. 2017, 66, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Hayward, C.J.; Ellis, D.; Foote, D.; Wilkinson, R.J.; Crosbie, P.B.B.; Bott, N.J.; Nowak, B.F. Concurrent Eepizootic hyper infections of sea lice (Predominantly Caligus chiastos) and blood flukes (Cardicola forsteri) in ranched Southern Bluefin Tuna. Vet. Parasitol. 2010, 173, 107–115. [Google Scholar] [CrossRef]
- Power, C. Southern Bluefin Tuna and Their Blood Flukes. Ph.D. Thesis, RMIT University, Melbourne, Australia, 2022. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pre-Treatment (Week 4) | Post-Treatment (Week 10) | Harvest (Week 16) | Cumulative Mortality (%) | |
|---|---|---|---|---|
| Company A | ||||
| Pontoon 1 | 6 | 0 | 15 | 0.54 |
| Pontoon 2 | 0 | 12 | 0 | 0.35 |
| Pontoon 3 | 6 | 0 | 0 | 0.60 |
| Company B | ||||
| Pontoon 1 | 4 | 11 | 0 | 3.00 |
| Pontoon 2 | 4 | 1 | 15 | 3.00 |
| Pontoon 3 | 4 | 0 | 0 | 2.00 |
| Transfer Date | Praziquantel Treatment Dose (mg/kg) | Weeks in Ranching | Cumulative Mortality (%) | |
|---|---|---|---|---|
| Company A | ||||
| 2021 | 24 March | 30 | 16 | 0.54 |
| 2019 | 1 March | 30 | 21 | 0.52 |
| 2018 | 27 February | 30 | 20 | 0.08 |
| Company B | ||||
| 2021 | 12 March | 30 | 17 | 3.00 |
| 2019 | 23 March | 18 | 17 | 0.27 |
| 2018 | 8 March | 42 | 18 | 2.33 |
| Adult C. forsteri Heart | C. forsteri (ITS-2) Heart | C. forsteri (ITS-2) Gills | C. orientalis (ITS-2) Gills | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OR | Z | p | OR | Z | p | OR | Z | p | OR | Z | p | |
| SBT Condition Index | 1.129 | 1.265 | 0.206 | 0.970 | 0.327 | 0.744 | 0.886 | 1.519 | 0.129 | 0.834 | 1.218 | 0.223 |
| Company | 1.330 | 0.588 | 0.556 | 0.196 | 2.927 | 0.003 | 0.406 | 2.186 | 0.029 | 1.000 | <0.001 | >0.999 |
| Time (weeks) | 1.118 | 2.665 | 0.008 | 1.043 | 1.082 | 0.279 | 0.940 | 1.907 | 0.057 | 0.779 | 2.347 | 0.019 |
| Adult C. forsteri Heart | C. forsteri (ITS-2) Gills | C. orientalis (ITS-2) Gills | |||||||
|---|---|---|---|---|---|---|---|---|---|
| OR | Z | p | OR | Z | p | OR | Z | p | |
| SBT Condition Index | 1.042 | 0.390 | 0.696 | 1.032 | 0.337 | 0.736 | 1.325 | 1.089 | 0.276 |
| Company | n.a. | n.a. | n.a. | 0.427 | 1.979 | 0.048 | n.a. | n.a. | n.a. |
| Time (weeks) | 1.113 | 2.465 | 0.014 | 1.057 | 1.528 | 0.126 | 0.659 | 1.890 | 0.059 |
| qPCR: Cardicola forsteri (ITS-2) | ||||
|---|---|---|---|---|
| + | − | Total | ||
| Heart Flush: Adult Cardicola forsteri | + | 19 | 3 | 22 |
| − | 56 | 20 | 76 | |
| Total | 75 | 23 | 98 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carabott, M.J.; Power, C.; Widdicombe, M.; Rough, K.; Nowak, B.F.; Bott, N.J. Dynamics of Cardicola spp. Infection in Ranched Southern Bluefin Tuna: First Observation of C. orientalis at Transfer. Pathogens 2023, 12, 1443. https://doi.org/10.3390/pathogens12121443
Carabott MJ, Power C, Widdicombe M, Rough K, Nowak BF, Bott NJ. Dynamics of Cardicola spp. Infection in Ranched Southern Bluefin Tuna: First Observation of C. orientalis at Transfer. Pathogens. 2023; 12(12):1443. https://doi.org/10.3390/pathogens12121443
Chicago/Turabian StyleCarabott, Melissa J., Cecilia Power, Maree Widdicombe, Kirsten Rough, Barbara F. Nowak, and Nathan J. Bott. 2023. "Dynamics of Cardicola spp. Infection in Ranched Southern Bluefin Tuna: First Observation of C. orientalis at Transfer" Pathogens 12, no. 12: 1443. https://doi.org/10.3390/pathogens12121443
APA StyleCarabott, M. J., Power, C., Widdicombe, M., Rough, K., Nowak, B. F., & Bott, N. J. (2023). Dynamics of Cardicola spp. Infection in Ranched Southern Bluefin Tuna: First Observation of C. orientalis at Transfer. Pathogens, 12(12), 1443. https://doi.org/10.3390/pathogens12121443