
Differential Analysis of Key Proteins Related to Fibrosis and Inflammation in Soluble Egg Antigen of Schistosoma mansoni at Different Infection Times



Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasites and Animals
2.2. Preparation of SEA
2.3. Protein Digestion and LC-MS/MS Analysis
2.4. Bioinformatics Analysis
2.5. LX-2 Cell Treatment by SEA of Different Infection Weeks
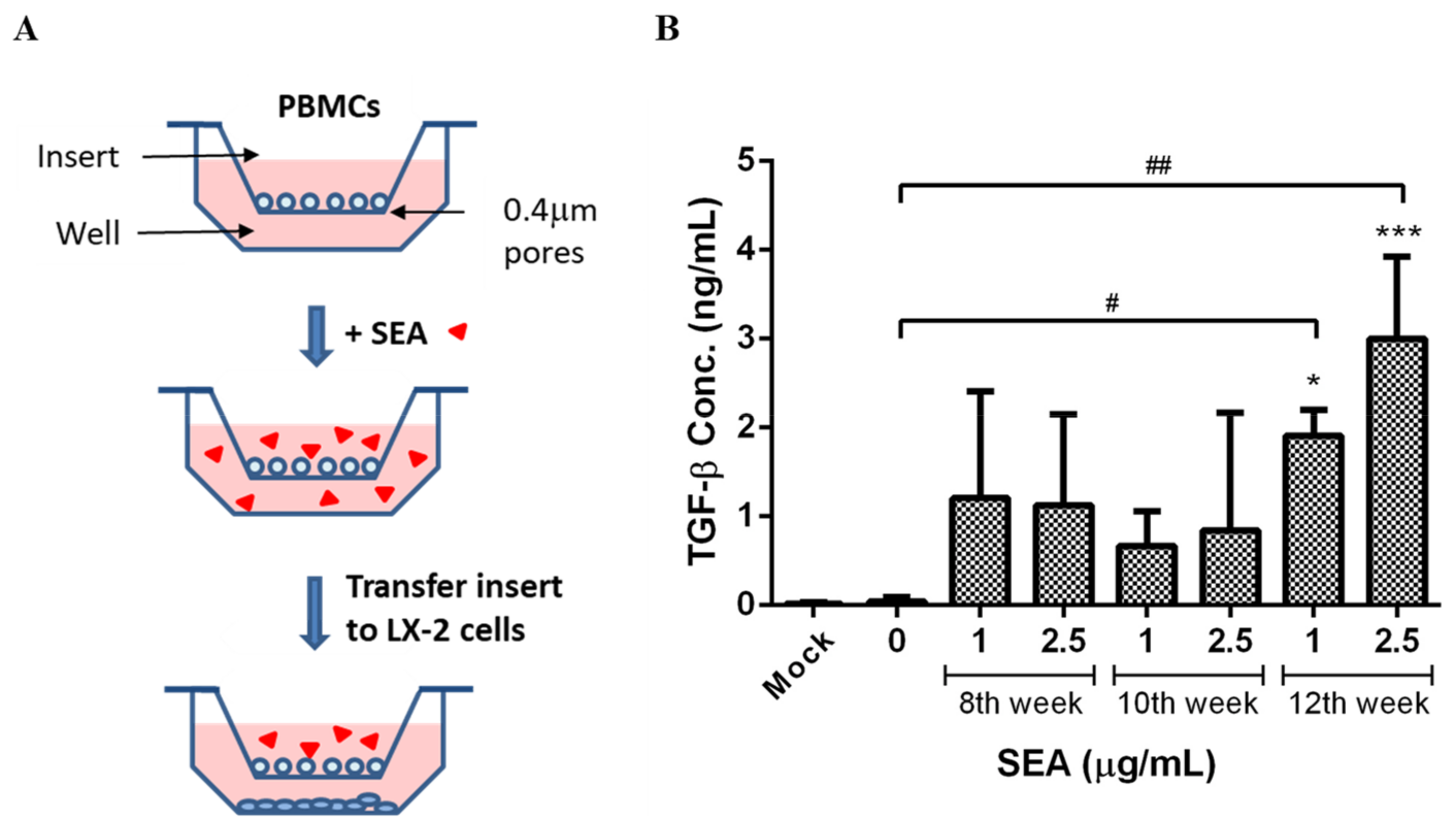
2.6. Evaluation of Liver Fibrosis Using In Vitro PBMC/LX-2 Co-Culture Model
2.7. Enzyme-Linked Immunosorbent Assays (ELISA)
2.8. Real-Time PCR Analysis
2.9. Statistical Analysis
3. Results
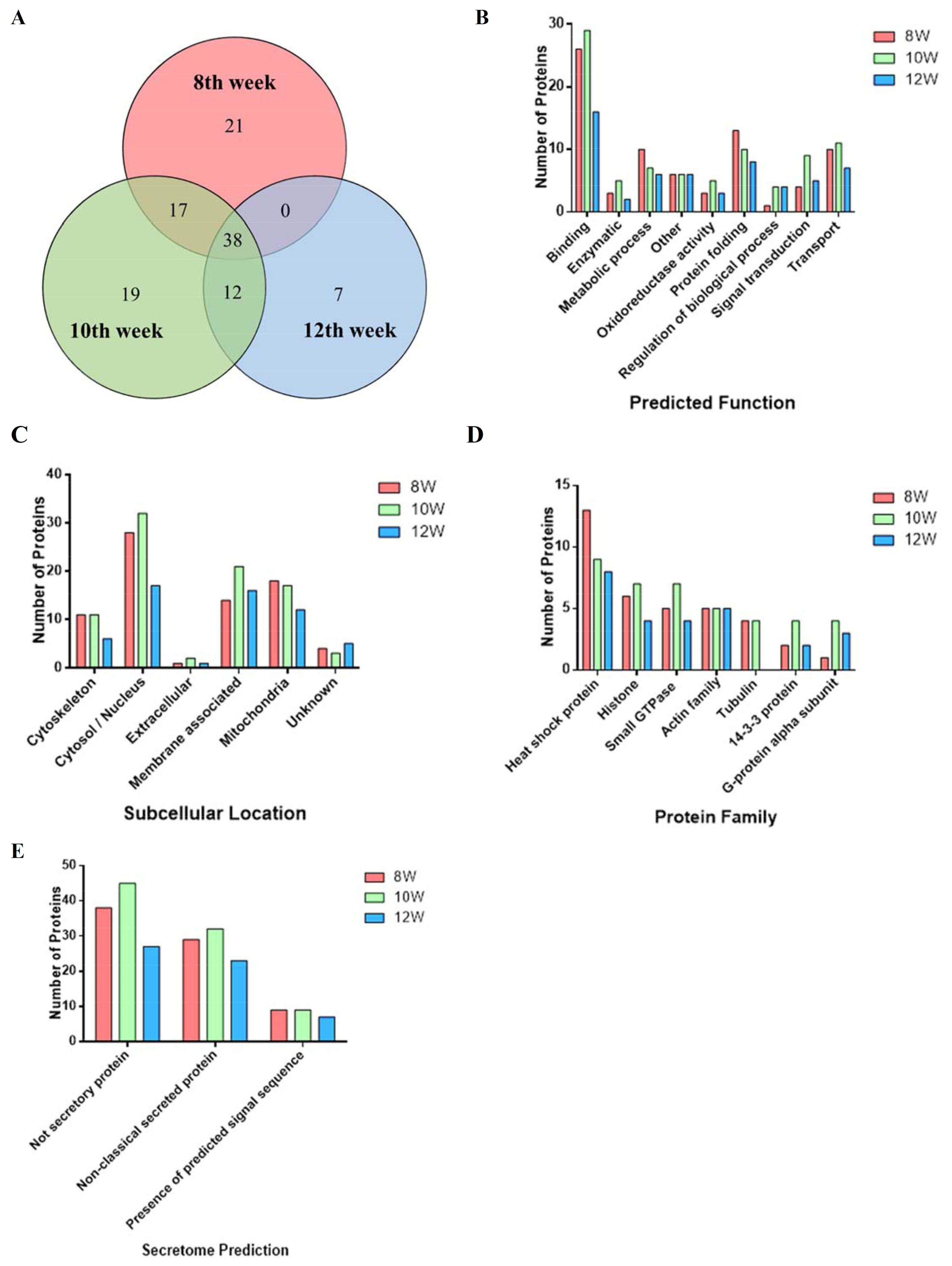
3.1. Proteomic Analysis of SEA from Different Infected Weeks
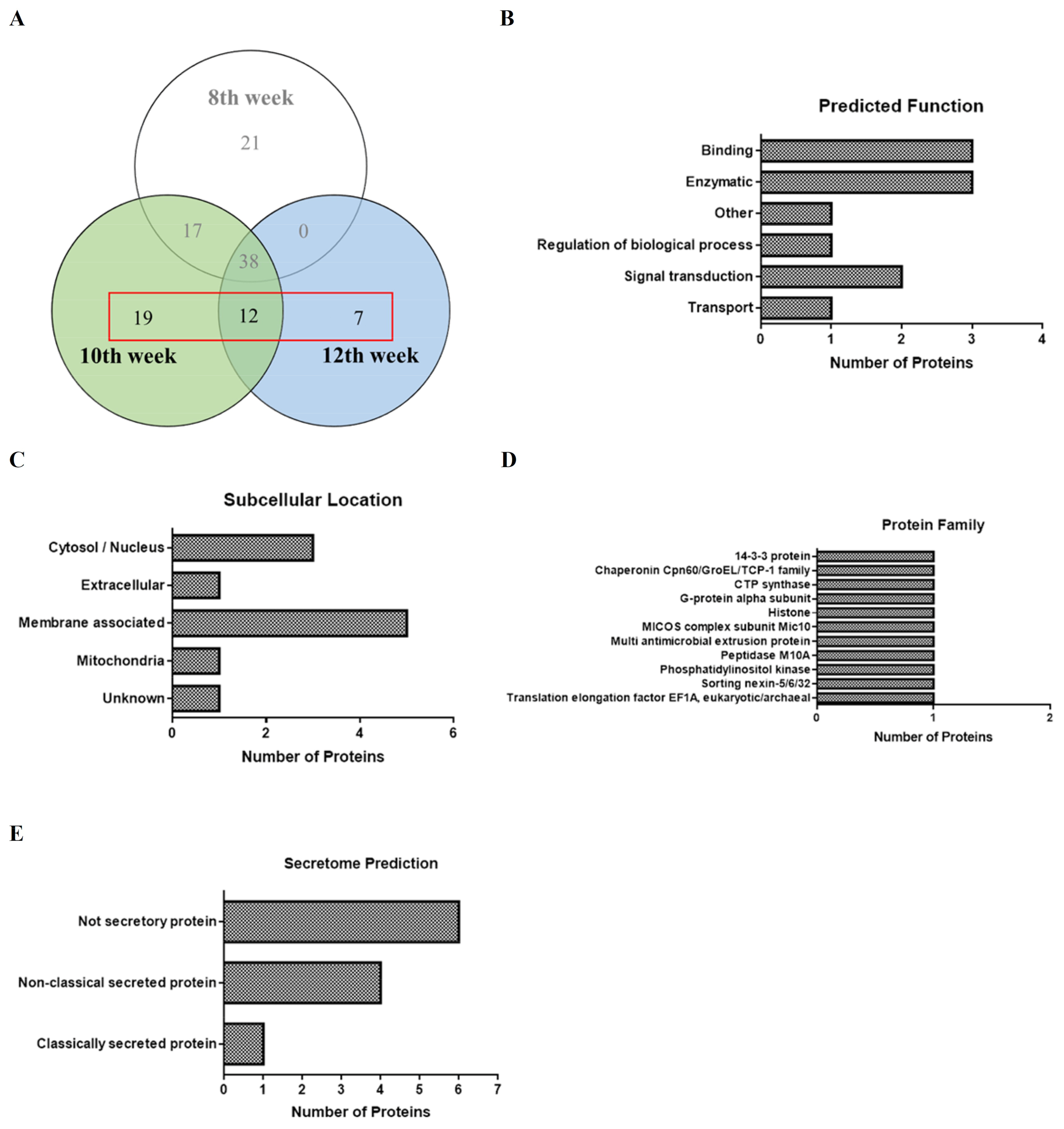
3.2. Proteins Identified in SEA Associated with Hepatic Fibrosis of Schistosomiasis
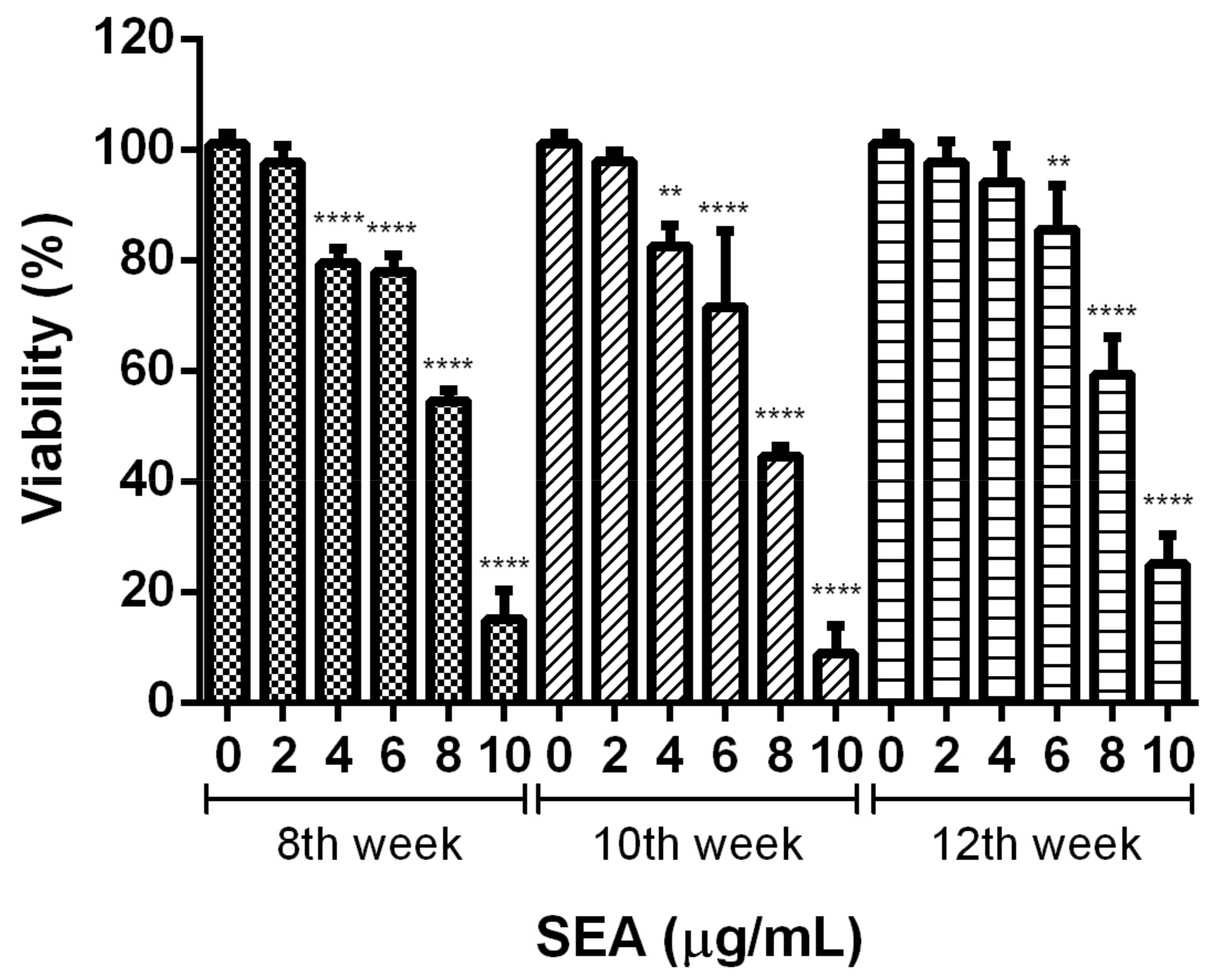
3.3. Effect of Different Infection Weeks on Activation Induction and Cell Viability of Human LX-2 Hepatic Stellate Cells Induced by SEA
3.4. Effect of SEA on TGF-β Expression in PBMCs Co-Cultured with LX-2
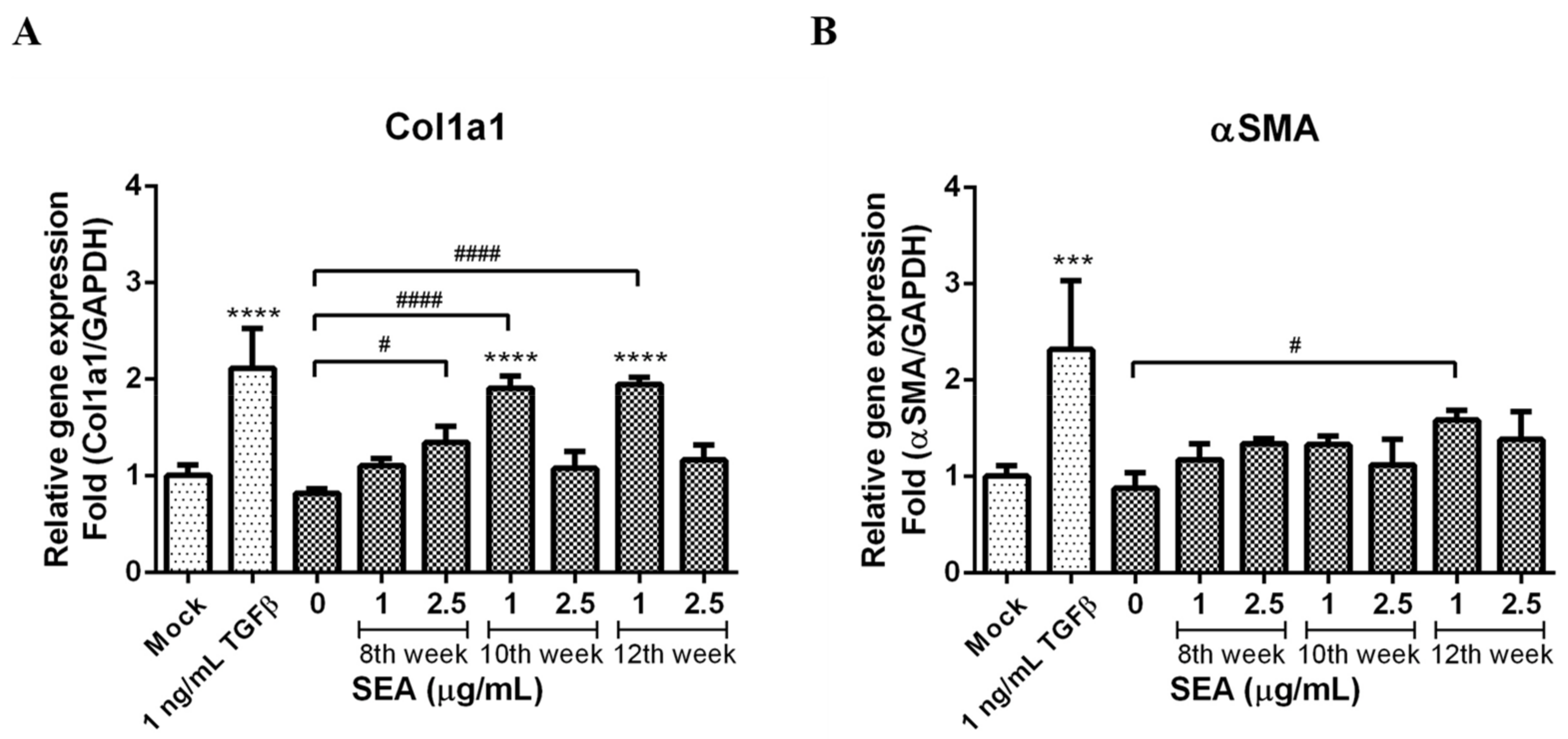
3.5. Effect of SEA on Col1a1 and α-SMA Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chitsulo, L.; Engels, D.; Montresor, A.; Savioli, L. The global status of schistosomiasis and its control. Acta Trop. 2000, 77, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Gryseels, B.; Polman, K.; Clerinx, J.; Kestens, L. Human schistosomiasis. Lancet 2006, 368, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.B.; Zheng, H.M.; Jiang, Q.W. A diagnostic challenge for Schistosomiasis japonica in China: Consequences on praziquantel-based morbidity control. Parasites Vectors 2011, 4, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, E.J.; MacDonald, A.S. The immunobiology of schistosomiasis. Nat. Rev. 2002, 2, 499–511. [Google Scholar] [CrossRef]
- Stadecker, M.J.; Asahi, H.; Finger, E.; Hernandez, H.J.; Rutitzky, L.I.; Sun, J. The immunobiology of Th1 polarization in high-pathology schistosomiasis. Immunol. Rev. 2004, 201, 168–179. [Google Scholar] [CrossRef]
- Cheever, A.W.; Hoffmann, K.F.; Wynn, T.A. Immunopathology of schistosomiasis mansoni in mice and men. Immunol. Today 2000, 21, 465–466. [Google Scholar] [CrossRef]
- Andrade, Z.A. Schistosomiasis and liver fibrosis. Parasite Immunol. 2009, 31, 656–663. [Google Scholar] [CrossRef]
- El-Sayed, N.M.; Fathy, G.M.; Abdel-Rahman, S.A.-R.; El-Shafei, M.A.-A. Cytokine patterns in experimental schistosomiasis mansoni infected mice treated with silymarin. J. Parasit. Dis. 2016, 40, 922–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuah, C.; Jones, M.K.; Burke, M.L.; McManus, D.P.; Gobert, G.N. Cellular and chemokine-mediated regulation in schistosome-induced hepatic pathology. Trends Parasitol. 2014, 30, 141–150. [Google Scholar] [CrossRef]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.-K. Hepatic Fibrosis Inhibitory Effect of Peptides Isolated from Navicula incerta on TGF-β1 Induced Activation of LX-2 Human Hepatic Stellate Cells. Prev. Nutr. Food. Sci. 2013, 18, 124–132. [Google Scholar] [CrossRef] [Green Version]
- Kamdem, S.D.; Moyou-Somo, R.; Brombacher, F.; Nono, J.K. Host Regulators of Liver Fibrosis During Human Schistosomiasis. Front. Immunol. 2018, 9, 2781. [Google Scholar] [CrossRef]
- Koyama, Y.; Brenner, D.A. Liver inflammation and fibrosis. J. Clin. Investig. 2017, 127, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Liang, Y.B.; Chen, Z.B.; Du, L.L.; Zeng, L.J.; Wu, J.G.; Yang, W.; Liang, H.P.; Ma, Z.F. Soluble Egg Antigen Activates M2 Macrophages via the STAT6 and PI3K Pathways, and Schistosoma Japonicum Alternatively Activates Macrophage Polarization to Improve the Survival Rate of Septic Mice. J. Cell. Biochem. 2017, 118, 4230–4239. [Google Scholar] [CrossRef]
- Masamba, P.; Kappo, A.P. Immunological and Biochemical Interplay between Cytokines, Oxidative Stress and Schistosomiasis. Int. J. Mol. Sci. 2021, 22, 7216. [Google Scholar] [CrossRef]
- Wang, L.; Liao, Y.; Yang, R.; Yu, Z.; Zhang, L.; Zhu, Z.; Wu, X.; Shen, J.; Liu, J.; Xu, L.; et al. Sja-miR-71a in Schistosome egg-derived extracellular vesicles suppresses liver fibrosis caused by schistosomiasis via targeting semaphorin 4D. J. Extracell. Vesicles. 2020, 9, 1785738. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Xu, Q.; Wang, Q.; Wu, H.; Hua, J. Effect of modulation of PPAR-γ activity on Kupffer cells M1/M2 polarization in the development of non-alcoholic fatty liver disease. Sci. Rep. 2017, 7, 44612. [Google Scholar] [CrossRef] [Green Version]
- Lau, A.O.; Sacci, J.B., Jr.; Azad, A.F. Host responses to Plasmodium yoelii hepatic stages: A paradigm in host-parasite interaction. J. Immunol. 2001, 166, 1945–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkaid, Y.; Sun, C.M.; Bouladoux, N. Parasites and immunoregulatory T cells. Curr. Opin. Immunol. 2006, 18, 406–412. [Google Scholar] [CrossRef]
- Cheng, P.C.; Lin, C.N.; Chen, Y.J.; Chang, F.S.; Tsaihong, J.C.; Lee, K.M. Triggering receptor expressed on myeloid cells (TREM)-1 participates in Schistosoma mansoni inflammatory responses. Parasite Immunol. 2011, 33, 276–286. [Google Scholar] [CrossRef]
- Tang, G.X.; Zhou, H.J.; Xu, J.W.; Xu, J.M.; Ji, M.J.; Wu, H.W.; Wu, G.L. Schistosoma japonicum soluble egg antigens attenuate IFN-gamma-induced MHC class II expression in RAW 264.7 macrophages. PLoS ONE 2012, 7, e49234. [Google Scholar] [CrossRef]
- Herbert, D.R.; Holscher, C.; Mohrs, M.; Arendse, B.; Schwegmann, A.; Radwanska, M.; Leeto, M.; Kirsch, R.; Hall, P.; Mossmann, H.; et al. Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity 2004, 20, 623–635. [Google Scholar] [CrossRef]
- Barron, L.; Smith, A.M.; El Kasmi, K.C.; Qualls, J.E.; Huang, X.; Cheever, A.; Borthwick, L.A.; Wilson, M.S.; Murray, P.J.; Wynn, T.A. Role of arginase 1 from myeloid cells in th2-dominated lung inflammation. PLoS ONE 2013, 8, e61961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiemessen, M.M.; Jagger, A.L.; Evans, H.G.; van Herwijnen, M.J.; John, S.; Taams, L.S. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 19446–19451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, M.G.; Du, Y.; Perrigoue, J.G.; Zaph, C.; Taylor, J.J.; Goldschmidt, M.; Swain, G.P.; Yancopoulos, G.D.; Valenzuela, D.M.; Murphy, A.; et al. Alternatively activated macrophage-derived RELM-{alpha} is a negative regulator of type 2 inflammation in the lung. J. Exp. Med. 2009, 206, 937–952. [Google Scholar] [CrossRef]
- Ito, T.; Schaller, M.; Raymond, T.; Joshi, A.D.; Coelho, A.L.; Frantz, F.G.; Carson, W.F.t.; Hogaboam, C.M.; Lukacs, N.W.; Standiford, T.J.; et al. Toll-like receptor 9 activation is a key mechanism for the maintenance of chronic lung inflammation. Am. J. Respir. Crit. Care Med. 2009, 180, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Scheer, S.; Krempl, C.; Kallfass, C.; Frey, S.; Jakob, T.; Mouahid, G.; Mone, H.; Schmitt-Graff, A.; Staeheli, P.; Lamers, M.C. S. mansoni bolsters anti-viral immunity in the murine respiratory tract. PLoS ONE 2014, 9, e112469. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.C.; Lin, C.N.; Peng, S.Y.; Li, L.L.; Luo, T.Y.; Fan, C.K.; Lee, K.M. A study of immunomodulatory genes responses to macrophages of Schistosoma japonicum infection during different stages by microarray analysis. Acta Trop. 2013, 127, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.H.; Cheng, C.H.; Huang, T.W.; Peng, S.Y.; Lee, K.M.; Cheng, P.C. Switched phenotypes of macrophages during the different stages of Schistosoma japonicum infection influenced the subsequent trends of immune responses. J. Microbiol. Immunol. Infect. 2021, 55, 503–526. [Google Scholar] [CrossRef]
- Fallon, P.G.; Dunne, D.W. Tolerization of mice to Schistosoma mansoni egg antigens causes elevated type 1 and diminished type 2 cytokine responses and increased mortality in acute infection. J. Immunol. 1999, 162, 4122–4132. [Google Scholar] [CrossRef]
- Smith, P.; Fallon, R.E.; Mangan, N.E.; Walsh, C.M.; Saraiva, M.; Sayers, J.R.; McKenzie, A.N.; Alcami, A.; Fallon, P.G. Schistosoma mansoni secretes a chemokine binding protein with antiinflammatory activity. J. Exp. Med. 2005, 202, 1319–1325. [Google Scholar] [CrossRef]
- Knuhr, K.; Langhans, K.; Nyenhuis, S.; Viertmann, K.; Kildemoes, A.M.O.; Doenhoff, M.J.; Haas, H.; Schramm, G. Schistosoma mansoni Egg-Released IPSE/alpha-1 Dampens Inflammatory Cytokine Responses via Basophil Interleukin (IL)-4 and IL-13. Front. Immunol. 2018, 9, 2293. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.Y.; Lee, Y.L.; Chiang, M.H.; Wang, H.Y.; Chen, C.Y.; Lin, C.H.; Chen, Y.C.; Fan, C.K.; Cheng, P.C. Schistosoma egg antigens suppress LPS-induced inflammation in human IMR-90 cells by modulation of JAK/STAT1 signaling. J. Microbiol. Immunol. Infect. 2020, 54, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, X.; Chen, Y.; Zhang, L.S.; Zhang, Y.R.; Ji, D.R.; Liu, S.M.; Jia, M.Z.; Zhu, Y.H.; Qi, Y.F.; et al. STAT3 Promotes Schistosome-Induced Liver Injury by Inflammation, Oxidative Stress, Proliferation, and Apoptosis Signal Pathway. Infect. Immun. 2021, 89, e00309-20. [Google Scholar] [CrossRef]
- Lam, H.Y.P.; Cheng, P.C.; Peng, S.Y. Resolution of systemic complications in Schistosoma mansoni-infected mice by concomitant treatment with praziquantel and Schisandrin B. Int. J. Parasitol. 2021, 52, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.Y.P.; Liang, T.R.; Lan, Y.C.; Chang, K.C.; Cheng, P.C.; Peng, S.Y. Antifibrotic and anthelminthic effect of casticin on Schistosoma mansoni-infected BALB/c mice. J. Microbiol. Immunol. Infect. 2021, 55, 314–322. [Google Scholar] [CrossRef]
- Lin, M.H.; Lee, K.M.; Hsu, C.Y.; Peng, S.Y.; Lin, C.N.; Chen, C.C.; Fan, C.K.; Cheng, P.C. Immunopathological effects of Agaricus blazei Murill polysaccharides against Schistosoma mansoni infection by Th1 and NK1 cells differentiation. Int. Immunopharmacol. 2019, 73, 502–514. [Google Scholar] [CrossRef] [PubMed]
- Obieglo, K.; Schuijs, M.J.; Ozir-Fazalalikhan, A.; Otto, F.; van Wijck, Y.; Boon, L.; Lambrecht, B.N.; Taube, C.; Smits, H.H. Isolated Schistosoma mansoni eggs prevent allergic airway inflammation. Parasite Immunol. 2018, 40, e12579. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.; Fan, X.; Yang, Y.; Dong, P.; Zhou, W.; Xu, Y.; Zhou, Y.; Guo, F.; Zheng, Y.; Yang, J.Q. Schistosoma japonicum infection downregulates house dust mite-induced allergic airway inflammation in mice. PLoS ONE 2017, 12, e0179565. [Google Scholar] [CrossRef] [Green Version]
- van der Vlugt, L.; Obieglo, K.; Ozir-Fazalalikhan, A.; Sparwasser, T.; Haeberlein, S.; Smits, H.H. Schistosome-induced pulmonary B cells inhibit allergic airway inflammation and display a reduced Th2-driving function. Int. J. Parasitol. 2017, 47, 545–554. [Google Scholar] [CrossRef]
- Morais, S.B.; Figueiredo, B.C.; Assis, N.R.G.; Alvarenga, D.M.; de Magalhaes, M.T.Q.; Ferreira, R.S.; Vieira, A.T.; Menezes, G.B.; Oliveira, S.C. Schistosoma mansoni SmKI-1 serine protease inhibitor binds to elastase and impairs neutrophil function and inflammation. PLoS Pathog. 2018, 14, e1006870. [Google Scholar] [CrossRef] [Green Version]
- De Marco Verissimo, C.; Potriquet, J.; You, H.; McManus, D.P.; Mulvenna, J.; Jones, M.K. Qualitative and quantitative proteomic analyses of Schistosoma japonicum eggs and egg-derived secretory-excretory proteins. Parasites Vectors 2019, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Ashton, P.D.; Curwen, R.S.; Wilson, R.A. Linking proteome and genome: How to identify parasite proteins. Trends Parasitol. 2001, 17, 198–202. [Google Scholar] [CrossRef]
- Cass, C.L.; Johnson, J.R.; Califf, L.L.; Xu, T.; Hernandez, H.J.; Stadecker, M.J.; Yates, J.R., 3rd; Williams, D.L. Proteomic analysis of Schistosoma mansoni egg secretions. Mol. Biochem. Parasitol. 2007, 155, 84–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewalick, S.; Bexkens, M.L.; van Balkom, B.W.; Wu, Y.P.; Smit, C.H.; Hokke, C.H.; de Groot, P.G.; Heck, A.J.; Tielens, A.G.; van Hellemond, J.J. The proteome of the insoluble Schistosoma mansoni eggshell skeleton. Int. J. Parasitol. 2011, 41, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Carson, J.P.; Robinson, M.W.; Hsieh, M.H.; Cody, J.; Le, L.; You, H.; McManus, D.P.; Gobert, G.N. A comparative proteomics analysis of the egg secretions of three major schistosome species. Mol. Biochem. Parasitol. 2020, 240, 111322. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Ivashkiv, L.B. Cross-regulation of signaling pathways by interferon-gamma: Implications for immune responses and autoimmune diseases. Immunity 2009, 31, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Speziali, E.; Aranha, C.H.; Teixeira-Carvalho, A.; Santiago, A.F.; Oliveira, R.P.; Rezende, M.C.; Carneiro, C.M.; Negrao-Correa, D.; Coelho, P.M.; Faria, A.M. Ageing down-modulates liver inflammatory immune responses to schistosome infection in mice. Scand. J. Immunol. 2010, 71, 240–248. [Google Scholar] [CrossRef]
- Bernardes, W.P.d.O.S.; de Araújo, J.M.; Carvalho, G.B.; Alves, C.C.; de Moura Coelho, A.T.; Dutra, I.T.S.; Teixeira, S.S.F.; de Azambuja Ribeiro, R.I.M.; de Moraes Mourão, M.; da Silva-Pereira, R.A.; et al. Sm16, A Schistosoma mansoni Immunomodulatory Protein, Fails to Elicit a Protective Immune Response and Does Not Have an Essential Role in Parasite Survival in the Definitive Host. J. Immunol. Res. 2019, 2019, 6793596. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Madungwe, N.B.; Bopassa, J.C. Mitochondrial inner membrane protein, Mic60/mitofilin in mammalian organ protection. J. Cell. Physiol. 2019, 234, 3383–3393. [Google Scholar] [CrossRef]
- Chen, H.; Gan, Q.; Yang, C.; Peng, X.; Qin, J.; Qiu, S.; Jiang, Y.; Tu, S.; He, Y.; Li, S.; et al. A novel role of glutathione S-transferase A3 in inhibiting hepatic stellate cell activation and rat hepatic fibrosis. J. Transl. Med. 2019, 17, 280. [Google Scholar] [CrossRef] [Green Version]
- Tlatelpa-Romero, B.; Cázares-Ordoñez, V.; Oyarzábal, L.F.; Vázquez-de-Lara, L.G. The Role of Pulmonary Surfactant Phospholipids in Fibrotic Lung Diseases. Int. J. Mol. Sci. 2022, 24, 326. [Google Scholar] [CrossRef]
- Retra, K.; deWalick, S.; Schmitz, M.; Yazdanbakhsh, M.; Tielens, A.G.; Brouwers, J.F.; van Hellemond, J.J. The tegumental surface membranes of Schistosoma mansoni are enriched in parasite-specific phospholipid species. Int. J. Parasitol. 2015, 45, 629–636. [Google Scholar] [CrossRef]
- Wigén, J.; Löfdahl, A.; Bjermer, L.; Elowsson Rendin, L.; Westergren-Thorsson, G. Converging pathways in pulmonary fibrosis and Covid-19-The fibrotic link to disease severity. Respir. Med. X 2020, 2, 100023. [Google Scholar] [CrossRef]
- Gobert, G.N.; Burke, M.L.; McManus, D.P.; Ellis, M.K.; Chuah, C.; Ramm, G.A.; Wang, Y.; Li, Y. Transcriptional profiling of chronic clinical hepatic schistosomiasis japonica indicates reduced metabolism and immune responses. Parasitology 2015, 142, 1453–1468. [Google Scholar] [CrossRef]
- Keppler-Noreuil, K.M.; Rios, J.J.; Parker, V.E.R.; Semple, R.K.; Lindhurst, M.J.; Sapp, J.C.; Alomari, A.; Ezaki, M.; Dobyns, W.; Biesecker, L.G. PIK3CA-related overgrowth spectrum (PROS): Diagnostic and testing eligibility criteria, differential diagnosis, and evaluation. Am. J. Med. Genet. Part A 2015, 167, 287–295. [Google Scholar] [CrossRef] [Green Version]
- Abou-El-Naga, I.F. Review: Schistosoma mansoni phosphatidylinositol 3 kinase (PI3K)/Akt/mechanistic target of rapamycin (mTOR) signaling pathway. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2021, 256, 110632. [Google Scholar] [CrossRef] [PubMed]
- Gessner, A.; Konig, J.; Fromm, M.F. Contribution of multidrug and toxin extrusion protein 1 (MATE1) to renal secretion of trimethylamine-N-oxide (TMAO). Sci. Rep. 2018, 8, 6659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, A.; Bueno, M.; Fournier, A.E. Extracellular functions of 14-3-3 adaptor proteins. Cell. Signal. 2017, 31, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Hamrita, B.; Nasr, H.B.; Kabbage, M.; Hamann, P.; Kuhn, L.; Guillier, C.; Remadi, S.; Chaieb, A.; Khairi, H.; Chouchane, L.; et al. L’apport thérapeutique des approches protéomiques dans les cancers. Gynécologie Obs. Fertil. 2011, 39, 87–93. [Google Scholar] [CrossRef]
- Garcia-Marcos, M.; Ghosh, P.; Farquhar, M.G. GIV/Girdin Transmits Signals from Multiple Receptors by Triggering Trimeric G Protein Activation. J. Biol. Chem. 2015, 290, 6697–6704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Peng, D.; Zhang, F.; Ruan, L.; Sun, M. The Caenorhabditis elegans CUB-like-domain containing protein RBT-1 functions as a receptor for Bacillus thuringiensis Cry6Aa toxin. PLoS Pathog. 2020, 16, e1008501. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, K.L.; Cajigas-Du Ross, C.K.; Chaney, J.K.; Nagy, L.E. Role of the chemokine system in liver fibrosis: A narrative review. Dig. Med. Res. 2022, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, L.; Liang, Y.; Lu, L. Pathology and molecular mechanisms of Schistosoma japonicum-associated liver fibrosis. Front. Cell. Infect. Microbiol. 2022, 12, 1035765. [Google Scholar] [CrossRef] [PubMed]
- Fung, M.C.; Lau, M.T.; Chen, X.G. Expressed sequence tag (EST) analysis of a Schistosoma japonicum cercariae cDNA library. Acta Trop. 2002, 82, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Stavitsky, A.B. Regulation of granulomatous inflammation in experimental models of schistosomiasis. Infect. Immun. 2004, 72, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Cetre-Sossah, C.B.; Montesano, M.A.; Freeman, G.L., Jr.; Willard, M.T.; Colley, D.G.; Secor, W.E. Early responses associated with chronic pathology in murine schistosomiasis. Parasite Immunol. 2007, 29, 241–249. [Google Scholar] [CrossRef]
- Coutinho, H.M.; Acosta, L.P.; Wu, H.W.; McGarvey, S.T.; Su, L.; Langdon, G.C.; Jiz, M.A.; Jarilla, B.; Olveda, R.M.; Friedman, J.F.; et al. Th2 cytokines are associated with persistent hepatic fibrosis in human Schistosoma japonicum infection. J. Infect. Dis. 2007, 195, 288–295. [Google Scholar] [CrossRef] [Green Version]
- Pagan, A.J.; Ramakrishnan, L. The Formation and Function of Granulomas. Annu. Rev. Immunol. 2018, 36, 639–665. [Google Scholar] [CrossRef]
- Meevissen, M.H.; Balog, C.I.; Koeleman, C.A.; Doenhoff, M.J.; Schramm, G.; Haas, H.; Deelder, A.M.; Wuhrer, M.; Hokke, C.H. Targeted glycoproteomic analysis reveals that kappa-5 is a major, uniquely glycosylated component of Schistosoma mansoni egg antigens. Mol. Cell. Proteom. 2011, 10, M110.005710. [Google Scholar] [CrossRef] [Green Version]
- Abdulla, M.H.; Lim, K.C.; McKerrow, J.H.; Caffrey, C.R. Proteomic identification of IPSE/alpha-1 as a major hepatotoxin secreted by Schistosoma mansoni eggs. PLoS Negl. Trop. Dis. 2011, 5, e1368. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, B.J.; Newland, S.A.; Gibbs, S.E.; Tourlomousis, P.; Fernandes dos Santos, P.; Patel, M.N.; Hall, S.W.; Walczak, H.; Schramm, G.; Haas, H.; et al. The Schistosoma mansoni T2 ribonuclease omega-1 modulates inflammasome-dependent IL-1beta secretion in macrophages. Int. J. Parasitol. 2015, 45, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Chen, J.; Zhu, D.; Chen, L.; Wang, J.; Sun, X.; Hu, B.; Duan, Y. Egg antigen p40 of Schistosoma japonicum promotes senescence in activated hepatic stellate cells via SKP2/P27 signaling pathway. Sci. Rep. 2017, 7, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Yang, X.; Tian, J.; Wang, X.; Ji, Y.; El-Ashram, S.; Ren, C.; Shen, J.; Liu, M. JQ-1 ameliorates schistosomiasis liver fibrosis by suppressing JAK2 and STAT3 activation. Biomed. Pharmacother. 2021, 144, 112281. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; Balic, A.; Gomez-Escobar, N.; Nair, M.; Taylor, M.D.; Allen, J.E. Helminth parasites--masters of regulation. Immunol. Rev. 2004, 201, 89–116. [Google Scholar] [CrossRef]
- Huang, P.; Ma, H.; Cao, Y.; Zhan, T.; Zhang, T.; Wang, X.; Zhang, Y.; Xu, J.; Xia, C. Activation of primary hepatic stellate cells and liver fibrosis induced by targeting TGF-beta1/Smad signaling in schistosomiasis in mice. Parasites Vectors 2022, 15, 456. [Google Scholar] [CrossRef]
- Ren, L.L.; Li, X.J.; Duan, T.T.; Li, Z.H.; Yang, J.Z.; Zhang, Y.M.; Zou, L.; Miao, H.; Zhao, Y.Y. Transforming growth factor-beta signaling: From tissue fibrosis to therapeutic opportunities. Chem. Biol. Interact. 2023, 369, 110289. [Google Scholar] [CrossRef]
- Liu, B.; Tian, Y.; He, J.; Gu, Q.; Jin, B.; Shen, H.; Li, W.; Shi, L.; Yu, H.; Shan, G.; et al. The potential of mecciRNA in hepatic stellate cell to regulate progression of nonalcoholic hepatitis. J. Transl. Med. 2022, 20, 393. [Google Scholar] [CrossRef]
- Scherl-Mostageer, M.; Sommergruber, W.; Abseher, R.; Hauptmann, R.; Ambros, P.; Schweifer, N. Identification of a novel gene, CDCP1, overexpressed in human colorectal cancer. Oncogene 2001, 20, 4402–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.; Szabolcs, A.; Dutta, S.K.; Yaqoob, U.; Jagavelu, K.; Wang, L.; Leof, E.B.; Urrutia, R.A.; Shah, V.H.; Mukhopadhyay, D. Neuropilin-1 Mediates Divergent R-Smad Signaling and the Myofibroblast Phenotype*. J. Biol. Chem. 2010, 285, 31840–31848. [Google Scholar] [CrossRef] [Green Version]
- Predes, D.; Cruz, J.V.R.; Abreu, J.G.; Mendes, F.A. CUB domain-containing protein 1 (CDCP1) binds transforming growth factor beta family members and increase TGF-β1 signaling pathway. Exp. Cell Res. 2019, 383, 111499. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infection Weeks of SEA Protein Sequence | Accession Number | Identified Protein | Effect on Fibrotic Pathology | References |
|---|---|---|---|---|
| Protein sequences that appear only at the 10th week | AF109181.1 | Sorting nexin (Sm16) | Sm16 inhibits the activation of macrophages upon invasion, thereby inhibiting the activation of the host adaptive immune response. | [48] |
| A0A146MJ03 | MICOS complex subunit MIC10 | MISCO subunit Mic60 causes mitochondrial dysfunction and cell death, resulting in neurodegeneration, cardiac hypertrophy, and fibrosis. | [49] | |
| P09792 | Glutathione S-transferase class-mu 28 kDa isozyme | GSTA3 is a special target of AKF-PD and is at least partially responsible for its function of reducing ROS accumulation in HSCs and decreasing LPO levels in vivo. | [50] | |
| A0A5K4F3N0 | REVERSED CTP synthase | This enzyme is related to phosphatidylcholine biosynthesis. The ratio of phosphatidylinositol is consistently increased for the more chronic injuries of pulmonary fibrotic diseases. | [51,52] | |
| A0A3Q0KP46 | Matrix metallopeptidase-7 (M10 family) | Host MMPs are necessary for Coronaviruses regarding cell infiltration, survival, and replication, further implicating converging pathways in pulmonary fibrosis. | [53,54] | |
| A0A3Q0KRX5 | Phosphatidylinositol-4,5-bisphosphate 3-kinase | PI3K is related to the overgrowth of human tissues, including visceral fibrous tissue, lipoma overgrowth, and vascular malformations. | [55,56] | |
| A0A3Q0KMD5 | Multidrug and toxin extrusion protein | MATE1 transports TMAO across cell membranes in conjunction with OCT2 and thereby may contribute to its renal secretion. TMAO is associated with kidney fibrosis. | [57] | |
| Protein sequences co-occurring at the 10th and 12th weeks | Q9U491 | 14-3-3 epsilon | 14-3-3 epsilon protein plays a role in signal transduction. Involved in cell proliferation, differentiation, survival, and apoptosis. It is regarded as a potential biomarker of pulmonary fibrosis. | [58] |
| G4VAD2 | Elongation factor 1-α (EF1-α) | EF1-α acts as a membrane receptor for the cryptic anti-adhesion site of fibronectin and is involved in fibronectin inhibiting cell anchoring and promoting apoptosis or fibrosis. | [59] | |
| A0A3Q0KCR4 | Trimeric G-protein alpha o subunit | It acts as a molecular switch controlling extracellular signals sensed from G protein-coupled receptors (GPCRs) and is associated with fibrosis in humans. | [60] | |
| Protein sequences that appear only at the 12th week | A0A5K4F1G2 | CUB domain-containing protein | Also known as CDCP1, is a transmembrane glycoprotein that has been found to function in lung fibroblasts. | [61] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.-C.; Chen, I.-A.; Peng, S.-Y.; Cheng, P.-C. Differential Analysis of Key Proteins Related to Fibrosis and Inflammation in Soluble Egg Antigen of Schistosoma mansoni at Different Infection Times. Pathogens 2023, 12, 441. https://doi.org/10.3390/pathogens12030441
Chen Y-C, Chen I-A, Peng S-Y, Cheng P-C. Differential Analysis of Key Proteins Related to Fibrosis and Inflammation in Soluble Egg Antigen of Schistosoma mansoni at Different Infection Times. Pathogens. 2023; 12(3):441. https://doi.org/10.3390/pathogens12030441
Chicago/Turabian StyleChen, Ying-Chou, I-An Chen, Shih-Yi Peng, and Po-Ching Cheng. 2023. "Differential Analysis of Key Proteins Related to Fibrosis and Inflammation in Soluble Egg Antigen of Schistosoma mansoni at Different Infection Times" Pathogens 12, no. 3: 441. https://doi.org/10.3390/pathogens12030441
APA StyleChen, Y.-C., Chen, I.-A., Peng, S.-Y., & Cheng, P.-C. (2023). Differential Analysis of Key Proteins Related to Fibrosis and Inflammation in Soluble Egg Antigen of Schistosoma mansoni at Different Infection Times. Pathogens, 12(3), 441. https://doi.org/10.3390/pathogens12030441