
Combined Toxicity of the Most Common Indoor Aspergilli




Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Media
2.2. Cytotoxicity, Genotoxicity and Immunomodulation of Combined Extracted Aspergilli
2.2.1. Cell Culture and Treatment
2.2.2. MTT Proliferation Assay
2.2.3. Alkaline Comet Assay
2.2.4. Determination of Cytokine Levels
3. Statistics
4. Results
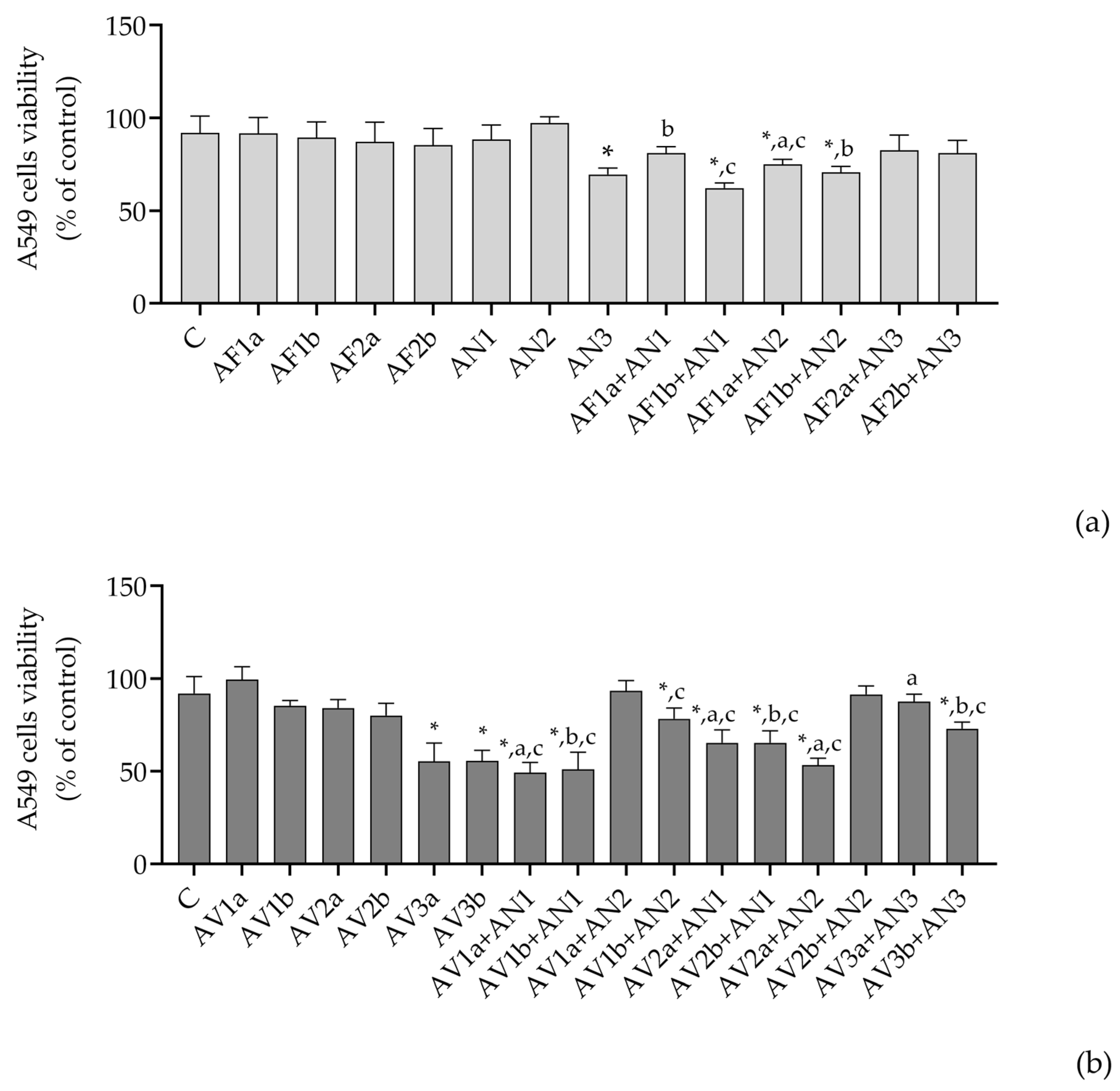
4.1. Cytotoxicity
4.2. Genotoxicity
4.3. Cytokines IL-1β, IL-5, IL-6, IL-8, IL-17, TNF-α Production
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- WHO. Guidelines for Indoor Air Quality and Dampness; World Health Organization: Geneva, Switzerland, 2010; ISBN 9789289002134.
- Cabral, J.P.S. Can We Use Indoor Fungi as Bioindicators of Indoor Air Quality? Historical Perspectives and Open Questions. Sci. Total Environ. 2010, 408, 4285–4295. [Google Scholar] [CrossRef] [PubMed]
- Schulz, T.; Senkpiel, K.; Ohgke, H. Comparison of the Toxicity of Reference Mycotoxins and Spore Extracts of Common Indoor Moulds. Int. J. Hyg. Environ. Health 2004, 207, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Piecková, E.; Jesenská, Z. Microscopic Fungi in Dwellings and Their Health Implications in Humans. Ann. Agric. Environ. Med. 1999, 6, 1–11. [Google Scholar] [PubMed]
- Nevalainen, A.; Täubel, M.; Hyvärinen, A. Indoor Fungi: Companions and Contaminants. Indoor Air 2015, 25. [Google Scholar] [CrossRef]
- Méheust, D.; Le Cann, P.; Reboux, G.; Millon, L.; Gangneux, J.-P. Indoor Fungal Contamination: Health Risks and Measurement Methods in Hospitals, Homes and Workplaces. Crit. Rev. Microbiol. 2014, 40, 248–260. [Google Scholar] [CrossRef]
- Redlich, C.A.; Sparer, J.; Cullen, M.R. Sick-Building Syndrome. Lancet 1997, 349, 1013–1016. [Google Scholar] [CrossRef]
- Piecková, E. Domestic Environment: Indoor Mycobiota As a Public Health Risk Factor. Environ. Mycol. Public Health 2016, 129–146. [Google Scholar] [CrossRef]
- Fog Nielsen, K. Mycotoxin Production by Indoor Molds. Fungal Genet. Biol. 2003, 39, 103–117. [Google Scholar] [CrossRef]
- Tuomi, T.; Reijula, K.; Johnsson, T.; Hemminki, K.; Hintikka, E.L.; Lindroos, O.; Kalso, S.; Koukila-Kähkölä, P.; Mussalo-Rauhamaa, H.; Haahtela, T. Mycotoxins in Crude Building Materials from Water-Damaged Buildings. Appl. Environ. Microbiol. 2000, 66, 1899–1904. [Google Scholar] [CrossRef]
- Horn, B.W. Biodiversity of Aspergillus Section Flavi in the United States: A Review. In Proceedings of the Food Additives and Contaminants. Food Addit. Contam. 2007, 24, 1088–1101. [Google Scholar] [CrossRef]
- Jakšić, D.; Kocsubé, S.; Bencsik, O.; Kecskeméti, A.; Szekeres, A.; Jelić, D.; Kopjar, N.; Vágvölgyi, C.; Varga, J.; Šegvić Klarić, M. Aflatoxin Production and in Vitro Toxicity of Aspergilli Section Flavi Isolated from Air Samples Collected from Different Environments. Mycotoxin Res. 2019, 35, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Jakšić, D.; Sertić, M.; Kocsubé, S.; Kovačević, I.; Kifer, D.; Mornar, A.; Nigović, B.; Klarić, M.Š. Post-Flood Impacts on Occurrence and Distribution of Mycotoxin-Producing Aspergilli from the Sections Circumdati, Flavi, and Nigri in Indoor Environment. J. Fungi 2020, 6, 282. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Hubka, V.; Ezekiel, C.N.; Hong, S.-B.; Nováková, A.; Chen, A.J.; Arzanlou, M.; Larsen, T.O.; Sklenář, F.; Mahakarnchanakul, W.; et al. Taxonomy of Aspergillus Section Flavi and Their Production of Aflatoxins, Ochratoxins and Other Mycotoxins. Stud. Mycol. 2019, 93, 1–63. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kocsubé, S.; Szigeti, G.; Baranyi, N.; Vágvölgyi, C.; Jakšić Despot, D.; Magyar, D.; Meijer, M.; Samson, R.A.; Šegvić Klarić, M.; et al. Occurrence of Black Aspergilli in Indoor Environments of Six Countries. Arh. Za Hig. Rada I Toksikol. Ind. Hyg. Toxicol. 2014, 65, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Jurjevic, Z.; Peterson, S.W.; Horn, B.W. Aspergillus Section Versicolores: Nine New Species and Multilocus DNA Sequence Based Phylogeny. IMA Fungus 2012, 3, 59–79. [Google Scholar] [CrossRef] [PubMed]
- Jakšić Despot, D.; Kocsubé, S.; Bencsik, O.; Kecskeméti, A.; Szekeres, A.; Vágvölgyi, C.; Varga, J.; Šegvić Klarić, M. Species Diversity and Cytotoxic Potency of Airborne Sterigmatocystin-Producing Aspergilli from the Section Versicolores. Sci. Total Environ. 2016, 562, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.A. Ecology and general characteristics of indoor fungi. In Fundamentals of Mold Growth in Indoor Environments and Strategies for Healthy Living, 1st ed.; Adan, C.G.O., Ed.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2011; pp. 101–116. [Google Scholar]
- Frisvad, J. Taxonomy, Chemodiversity, and Chemoconsistency of Aspergillus, Penicillium, and Talaromyces Species. Front. Microbiol. 2015, 5, 773. [Google Scholar] [CrossRef]
- Mousavi, B.; Hedayati, M.T.; Hedayati, N.; Ilkit, M.; Syedmousavi, S. Aspergillus Species in Indoor Environments and Their Possible Occupational and Public Health Hazards. Curr. Med. Mycol. 2016, 2, 36. [Google Scholar] [CrossRef]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, Identification and Nomenclature of the Genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef]
- Kocsubé, S.; Perrone, G.; Magistà, D.; Houbraken, J.; Varga, J.; Szigeti, G.; Hubka, V.; Hong, S.B.; Frisvad, J.C.; Samson, R.A. Aspergillus Is Monophyletic: Evidence from Multiple Gene Phylogenies and Extrolites Profiles. Stud. Mycol. 2016, 85, 199–213. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Samson, R.A.; Frisvad, J.C. Classification of Aspergillus, Penicillium, Talaromyces and Related Genera (Eurotiales): An Overview of Families, Genera, Subgenera, Sections, Series and Species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef] [PubMed]
- Sklenář, F.; Glässnerová, K.; Jurjević, Ž.; Houbraken, J.; Samson, R.A.; Visagie, C.M.; Yilmaz, N.; Gené, J.; Cano, J.; Chen, A.J.; et al. Taxonomy of Aspergillus Series Versicolores: Species Reduction and Lessons Learned about Intraspecific Variability. Stud. Mycol. 2022, 102, 53–93. [Google Scholar] [CrossRef] [PubMed]
- Bian, C.; Kusuya, Y.; Sklenář, F.; D’hooge, E.; Yaguchi, T.; Ban, S.; Visagie, C.M.; Houbraken, J.; Takahashi, H.; Hubka, V. Reducing the Number of Accepted Species in Aspergillus Series Nigri. Stud. Mycol. 2022, 102, 95–132. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Larsen, T.O. Chemodiversity in the Genus Aspergillus. Appl. Microbiol. Biotechnol. 2015, 99, 7859–7877. [Google Scholar] [CrossRef]
- Larsen, T.O.; Smedsgaard, J.; Nielsen, K.F.; Hansen, M.E.; Frisvad, J.C. Phenotypic Taxonomy and Metabolite Profiling in Microbial Drug Discovery. Nat. Prod. Rep. 2005, 22, 672–695. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The Use of Secondary Metabolite Profiling in Chemotaxonomy of Filamentous Fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Täubel, M.; Hyvärinen, A. Occurrence of Mycotoxins in Indoor Environments. Environ. Mycol. Public Health 2016, 299–323. [Google Scholar] [CrossRef]
- Visagie, C.M.; Hirooka, Y.; Tanney, J.B.; Whitfield, E.; Mwange, K.; Meijer, M.; Amend, A.S.; Seifert, K.A.; Samson, R.A. Aspergillus, Penicillium and Talaromyces Isolated from House Dust Samples Collected around the World. Stud. Mycol. 2014, 78, 63–139. [Google Scholar] [CrossRef]
- Engelhart, S.; Loock, A.; Skutlarek, D.; Sagunski, H.; Lommel, A.; Färber, H.; Exner, M. Occurrence of Toxigenic Aspergillus Versicolor Isolates and Sterigmatocystin in Carpet Dust from Damp Indoor Environments. Appl. Environ. Microbiol. 2002, 68, 3886–3890. [Google Scholar] [CrossRef]
- Viegas, C.; Almeida, B.; Monteiro, A.; Paciência, I.; Rufo, J.C.; Carolino, E.; Quintal-Gomes, A.; Twarużek, M.; Kosicki, R.; Marchand, G.; et al. Settled Dust Assessment in Clinical Environment: Useful for the Evaluation of a Wider Bioburden Spectrum. Int. J. Environ. Health Res. 2019, 31, 160–178. [Google Scholar] [CrossRef]
- Jakšić, D.; Sertić, M.; Kifer, D.; Kocsubè, S.; Mornar Turk, A.; Nigović, B.; Šarkanj, B.; Krska, R.; Sulyok, M.; Šegvić Klarić, M. Fungi and Their Secondary Metabolites in Water-Damaged Indoors after a Major Flood Event in Eastern Croatia. Indoor Air 2021, 31, 730–744. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Sulyok, M.; Lemons, A.R.; Green, B.J.; Cox-Ganser, J.M. Characterization of Fungi in Office Dust: Comparing Results of Microbial Secondary Metabolites, Fungal Internal Transcribed Spacer Region Sequencing, Viable Culture and Other Microbial Indices. Indoor Air 2018, 28, 708–720. [Google Scholar] [CrossRef]
- Peitzsch, M.; Sulyok, M.; Täubel, M.; Vishwanath, V.; Krop, E.; Borràs-Santos, A.; Hyvärinen, A.; Nevalainen, A.; Krska, R.; Larsson, L. Microbial Secondary Metabolites in School Buildings Inspected for Moisture Damage in Finland, The Netherlands and Spain. J. Environ. Monit. 2012, 14, 2044. [Google Scholar] [CrossRef] [PubMed]
- Korkalainen, M.; Täubel, M.; Naarala, J.; Kirjavainen, P.; Koistinen, A.; Hyvärinen, A.; Komulainen, H.; Viluksela, M. Synergistic Proinflammatory Interactions of Microbial Toxins and Structural Components Characteristic to Moisture-Damaged Buildings. Indoor Air 2017, 27, 13–23. [Google Scholar] [CrossRef]
- Jakšić, D.; Kocsubé, S.; Bencsik, O.; Kecskeméti, A.; Szekeres, A.; Jelić, D.; Kopjar, N.; Vágvölgyi, C.; Varga, J.; Šegvić Klarić, M. Fumonisin Production and Toxic Capacity in Airborne Black Aspergilli. Toxicol. Vitr. 2018, 53, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Smedsgaard, J. Micro-Scale Extraction Procedure for Standardized Screening of Fungal Metabolite Production in Cultures. J. Chromatogr. A 1997, 760, 264–270. [Google Scholar] [CrossRef]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A Simple Technique for Quantitation of Low Levels of DNA Damage in Individual Cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Collins, A.R. The Comet Assay for DNA Damage and Repair: Principles, Applications, and Limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Kifer, D.; Jakšić, D.; Šegvić Klarić, M. Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate? Toxins 2020, 12, 153. [Google Scholar] [CrossRef]
- Klas, K.R.; Kato, H.; Frisvad, J.C.; Yu, F.; Newmister, S.A.; Fraley, A.E.; Sherman, D.H.; Tsukamoto, S.; Williams, R.M. Structural and Stereochemical Diversity in Prenylated Indole Alkaloids Containing the Bicyclo[2.2.2]Diazaoctane Ring System from Marine and Terrestrial Fungi. Nat. Prod. Rep. 2018, 35, 532–558. [Google Scholar] [CrossRef]
- Kato, H.; Yoshida, T.; Tokue, T.; Nojiri, Y.; Hirota, H.; Ohta, T.; Williams, R.M.; Tsukamoto, S. Notoamides A-D: Prenylated Indole Alkaloids Isolated from a Marine-Derived Fungus, Aspergillus sp. Angew. Chem. Int. Ed. Engl. 2007, 46, 2254–2256. [Google Scholar] [CrossRef] [PubMed]
- Bünger, J.; Westphal, G.; Mönnich, A.; Hinnendahl, B.; Hallier, E.; Müller, M.; BUNGER, J. Cytotoxicity of Occupationally and Environmentally Relevant Mycotoxins. Toxicology 2004, 202, 199–211. [Google Scholar] [CrossRef]
- Pei, R.; Gunsch, C.K. Inflammatory Cytokine Gene Expression in THP-1 Cells Exposed to Stachybotrys Chartarum and Aspergillus Versicolor. Environ. Toxicol. 2013, 28, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Rakkestad, K.E.; Skaar, I.; Ansteinsson, V.E.; Solhaug, A.; Holme, J.A.; Pestka, J.J.; Samuelsen, J.T.; Dahlman, H.J.; Hongslo, J.K.; Becher, R. DNA Damage and DNA Damage Responses in THP-1 Monocytes after Exposure to Spores of Either Stachybotrys Chartarum or Aspergillus Versicolor or to T-2 Toxin. Toxicol. Sci. 2010, 115, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Roilides, E.; Gil Lamaignere, C.; Farmaki, E. Cytokines in Immunodeficient Patients with Invasive Fungal Infections: An Emerging Therapy. Int. J. Infect. Dis. 2002, 6, 154–163. [Google Scholar] [CrossRef]
- Suganuma, M.; Okabe, S.; Marino, M.; Sakai, A.; Sueoka, E.; Fujiki, H. Essential Role of Tumor Necrosis Factor α (TNF-α) in Tumor Promotion as Revealed by TNF-α-Deficient Mice. Cancer Res. 1999, 59, 4516–4518. [Google Scholar]
- Eder, C. Mechanisms of Interleukin-1beta Release. Immunobiology 2009, 214, 543–553. [Google Scholar] [CrossRef]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and Chemokines: At the Crossroads of Cell Signalling and Inflammatory Disease. Biochim. Biophys. Acta 2014, 1843, 2563–2582. [Google Scholar] [CrossRef]
- Carre, P.C.; Mortenson, R.L.; King, T.E.; Noble, P.W.; Sable, C.L.; Riches, D.W.H. Increased Expression of the Interleukin-8 Gene by Alveolar Macrophages in Idiopathic Pulmonary Fibrosis. A Potential Mechanism for the Recruitment and Activation of Neutrophils in Lung Fibrosis. J. Clin. Investig. 1991, 88, 1802–1810. [Google Scholar] [CrossRef]
- Duque, G.A.; Descoteaux, A. Macrophage Cytokines: Involvement in Immunity and Infectious Diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef]
- Adachi, T.; Alam, R. The Mechanism of IL-5 Signal Transduction. Am. J. Physiol. 1998, 275, C623–C633. [Google Scholar] [CrossRef] [PubMed]
- Kouro, T.; Takatsu, K. IL-5- and Eosinophil-Mediated Inflammation: From Discovery to Therapy. Int. Immunol. 2009, 21, 1303–1309. [Google Scholar] [CrossRef]
- Xu, X.; Xu, J.F.; Zheng, G.; Lu, H.W.; Duan, J.L.; Rui, W.; Guan, J.H.; Cheng, L.Q.; Yang, D.D.; Wang, M.C.; et al. CARD9S12N Facilitates the Production of IL-5 by Alveolar Macrophages for the Induction of Type 2 Immune Responses. Nat. Immunol. 2018, 19, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.L.; Li, D.S.; Wang, Z.Y.; Liu, Y.; Yang, J.M.; Li, C.Z.; Li, X.D.; Ma, J.D.; Zhang, M.M.; Lu, Y.J.; et al. Interleukin-17 Upregulation Participates in the Pathogenesis of Heart Failure in Mice via NF-ΚB-Dependent Suppression of SERCA2a and Cav1.2 Expression. Acta Pharmacol. Sin. 2021, 42, 1780–1789. [Google Scholar] [CrossRef]
- Kawaguchi, M.; Kokubu, F.; Fujita, J.; Huang, S.K.; Hizawa, N. Role of Interleukin-17F in Asthma. Inflamm. Allergy Drug Targets 2009, 8, 383–389. [Google Scholar] [CrossRef]
- Jolink, H.; de Boer, R.; Hombrink, P.; Jonkers, R.E.; van Dissel, J.T.; Falkenburg, J.H.F.; Heemskerk, M.H.M. Pulmonary Immune Responses against Aspergillus Fumigatus Are Characterized by High Frequencies of IL-17 Producing T-Cells. J. Infect. 2017, 74, 81–88. [Google Scholar] [CrossRef]
- Murdock, B.J.; Falkowski, N.R.; Shreiner, A.B.; Sadighi Akha, A.A.; McDonald, R.A.; White, E.S.; Toews, G.B.; Huffnagle, G.B. Interleukin-17 Drives Pulmonary Eosinophilia Following Repeated Exposure to Aspergillus Fumigatus Conidia. Infect. Immun. 2012, 80, 1424–1436. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Payne, G.A. Genetic Regulation of Aflatoxin Biosynthesis: From Gene to Genome. Fungal Genet. Biol. 2009, 46, 113–125. [Google Scholar] [CrossRef]
- Samson, R.A. Cellular Constitution, Water and Nutritional Needs, and Secondary Metabolites. Environ. Mycol. Public Health 2016, 5–15. [Google Scholar] [CrossRef]
- Samson, R.A.; Houbraken, J.; Thrane, U.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi, 2nd ed.; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010; ISBN 978-90-70351-82-3. [Google Scholar]
- Kelman, B.J.; Robbins, C.A.; Swenson, L.J.; Hardin, B.D. Risk from Inhaled Mycotoxins in Indoor Office and Residential Environments. Int. J. Toxicol. 2004, 23, 3–10. [Google Scholar] [CrossRef]
- Górny, R.L.; Reponen, T.; Willeke, K.; Schmechel, D.; Robine, E.; Boissier, M.; Grinshpun, S.A. Fungal Fragments as Indoor Air Biocontaminants. Appl. Environ. Microbiol. 2002, 68, 3522–3531. [Google Scholar] [CrossRef]
- Täubel, M.; Sulyok, M.; Vishwanath, V.; Bloom, E.; Turunen, M.; Järvi, K.; Kauhanen, E.; Krska, R.; Hyvärinen, A.; Larsson, L.; et al. Co-Occurrence of Toxic Bacterial and Fungal Secondary Metabolites in Moisture-Damaged Indoor Environments. Indoor Air 2011, 21, 368–375. [Google Scholar] [CrossRef]
- Richard, J.L.; Plattner, R.D.; May, J.; Liska, S.L. The Occurrence of Ochratoxin A in Dust Collected from a Problem Household. Mycopathologia 1999, 146, 99–103. [Google Scholar] [CrossRef]
- Mayer, S.; Mota, I.A.; Borrego, L.M.; Brera, C.; Debegnach, F.; Gregori, E.; Colicchia, S.; Soricelli, S.; Miano, B.; Magri, M.C.; et al. Fungi and Mycotoxins Risk Assessment and Management. In Environmental Mycology in Public Health, 1st ed.; Viegas, C., Pinheiro, C., Sabino, R., Viegas, S., Brandão, J., Veríssimo, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; ISBN 9780124114715. [Google Scholar]
- Cox, B.; Barton, P.; Class, R.; Coxhead, H.; Delatour, C.; Gillent, E.; Henshall, J.; Isin, E.M.; King, L.; Valentin, J.P. Setup of Human Liver-Chips Integrating 3D Models, Microwells and a Standardized Microfluidic Platform as Proof-of-Concept Study to Support Drug Evaluation. Biomater. Biosyst. 2022, 7, 100054. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Description | Short Code | Reference | |
|---|---|---|---|
| Control | 0.1% DMSO in RPMI medium | C | |
| section Flavi | A. flavus 0.1 mg/mL (0.75 µM AFB1) | AF1a | [12] |
| A. flavus 0.2 mg/mL (1.5 µM AFB1) | AF1b | ||
| A. flavus 0.05 mg/mL (AFB1 negative) | AF2a | ||
| A. flavus 0.1 mg/mL (AFB1 negative) | AF2b | ||
| series Nigri | A. niger 0.1 mg/mL (0.00009 µM FB2) | AN1 | [37] |
| A. welwitschiae 0.1 mg/mL (0.095 µM FB2) | AN2 | ||
| A. luchuensis 0.1 mg/mL (FB2 negative) | AN3 | ||
| series Versicolores | A. jensenii 0.005 mg/mL (0.1 µM STC) | AV1a | [17] |
| A. jensenii 0.05 mg/mL (1 µM STC) | AV1b | ||
| A. creber 0.006 mg/mL (0.1 µM STC) | AV2a | ||
| A. creber 0.06 mg/mL (1 µM STC) | AV2b | ||
| A. protuberus 0.05 mg/mL (0.0016 µM STC) | AV3a | ||
| A. protuberus 0.1 mg/mL (0.0024 µM STC) | AV3b |
| Abbreviations for Each Combination | |
|---|---|
| Flavi + Nigri | AF1a + AN1 |
| AF1b + AN1 | |
| AF1a + AN2 | |
| AF1b + AN2 | |
| AF2a + AN3 | |
| AF2b + AN3 | |
| Versicolores + Nigri | AV1a + AN1 |
| AV1b + AN1 | |
| AV1a + AN2 | |
| AV1b + AN2 | |
| AV2a + AN1 | |
| AV2b + AN1 | |
| AV2a + AN2 | |
| AV2b + AN2 | |
| AV3a + AN3 | |
| AV3b + AN3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakšić, D.; Jelić, D.; Kopjar, N.; Šegvić Klarić, M. Combined Toxicity of the Most Common Indoor Aspergilli. Pathogens 2023, 12, 459. https://doi.org/10.3390/pathogens12030459
Jakšić D, Jelić D, Kopjar N, Šegvić Klarić M. Combined Toxicity of the Most Common Indoor Aspergilli. Pathogens. 2023; 12(3):459. https://doi.org/10.3390/pathogens12030459
Chicago/Turabian StyleJakšić, Daniela, Dubravko Jelić, Nevenka Kopjar, and Maja Šegvić Klarić. 2023. "Combined Toxicity of the Most Common Indoor Aspergilli" Pathogens 12, no. 3: 459. https://doi.org/10.3390/pathogens12030459
APA StyleJakšić, D., Jelić, D., Kopjar, N., & Šegvić Klarić, M. (2023). Combined Toxicity of the Most Common Indoor Aspergilli. Pathogens, 12(3), 459. https://doi.org/10.3390/pathogens12030459