
Hatchability of Fascioloides magna Eggs in Cervids


Abstract
:
1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Parasitological Procedure
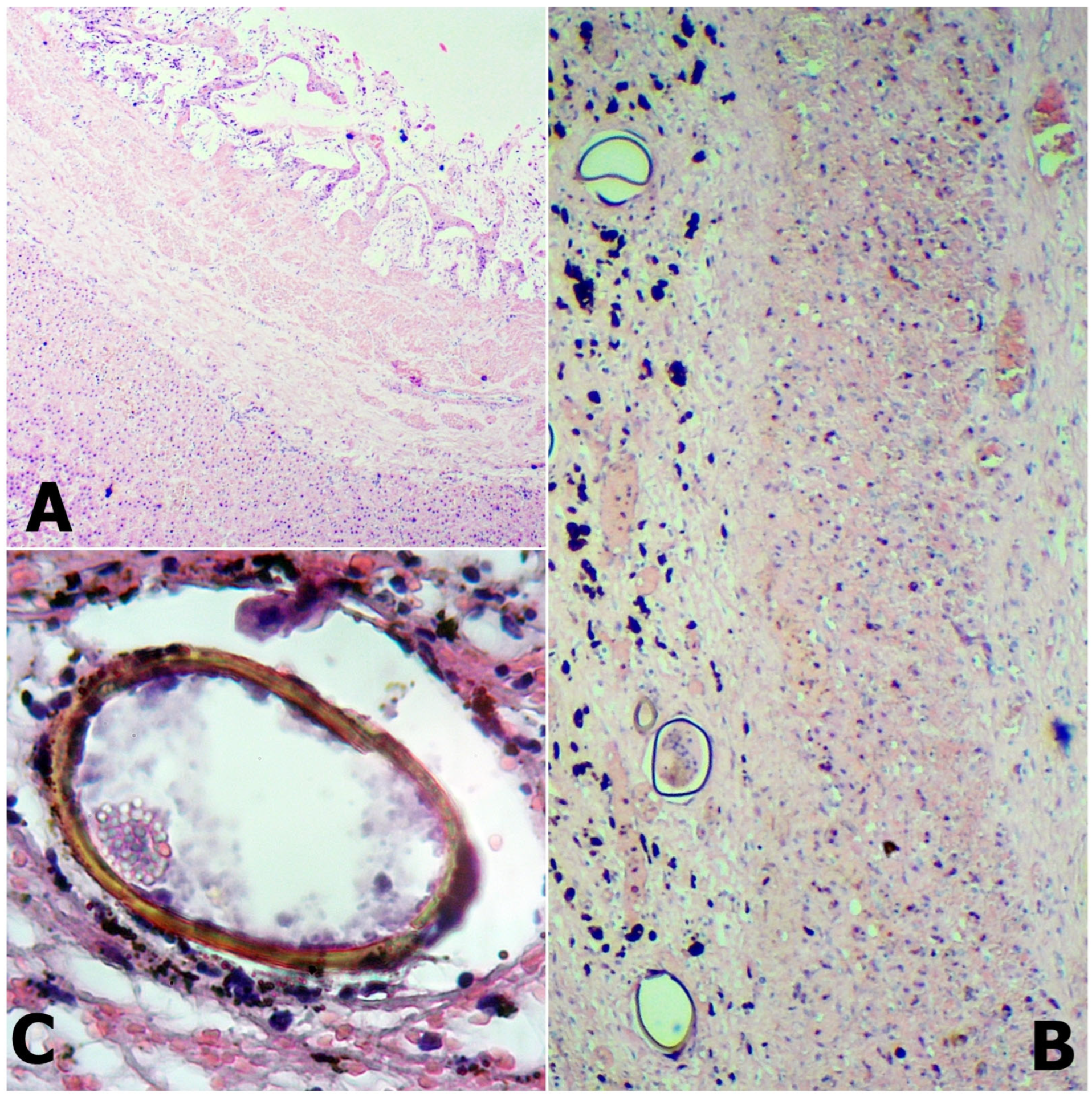
2.3. Histopathology
2.4. Hatching Procedure
2.5. Statistical Analysis
3. Results
3.1. Parasitological Findings
3.2. Histopathology
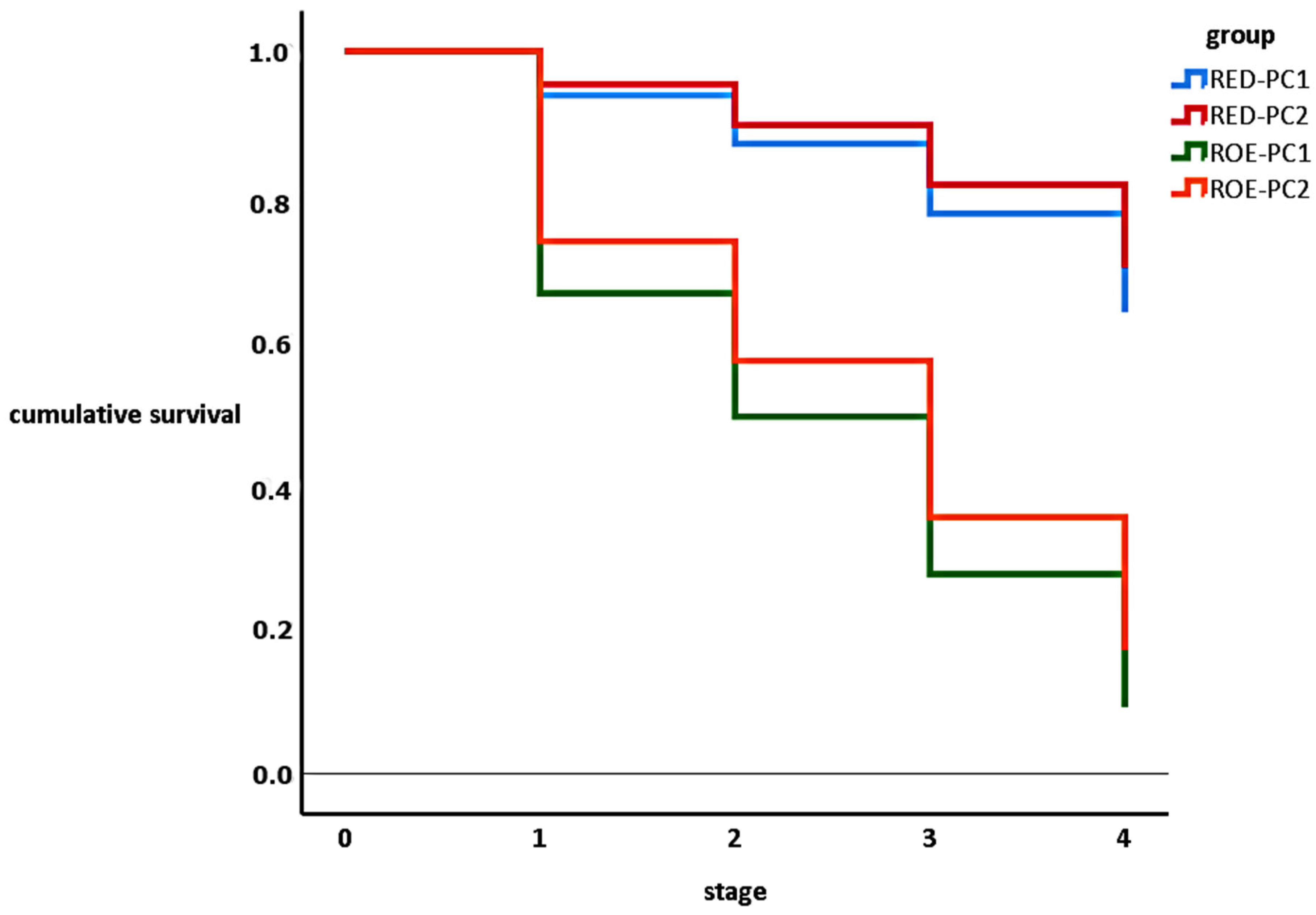
3.3. Egg Hatching
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pybus, M.J. Liver flukes. In Parasitic Diseases of Wild Mammals; Samuel, W.M., Pybus, M.J., Kocan, A.A., Eds.; Iowa State Press: Ames, IA, USA, 2021; pp. 121–149. [Google Scholar]
- Juhász, A.; Stothard, J.R. The giant liver fluke in Europe: A review of Fascioloides magna within cervids and livestock with considerations on an expanding snail-fluke transmission risk. Adv. Parasitol. 2023, 119, 223–257. [Google Scholar] [PubMed]
- Králová-Hromadová, I.; Juhásová, L.; Bazsalovicsová, E. The Giant Liver Fluke, Fascioloides magna: Past, Present and Future Research; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Malcicka, M. Life history and biology of Fascioloides magna (Trematoda) and its native and exotic hosts. Ecol. Evol. 2015, 5, 1381–1397. [Google Scholar] [CrossRef]
- Konjević, D.; Bujanić, M.; Beck, A.; Beck, R.; Martinković, F.; Janicki, Z. First record of chronic Fascioloides magna infection in roe deer (Capreolus capreolus). Int. J. Parasitol. Parasites Wildl. 2021, 15, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Filip-Hutsch, K.; Pyziel-Serafin, A.M.; Hutsch, T.; Bulak, K.; Czopowicz, M.; Merta, D.; Kobielski, J.; Demiaszkiewicz, A.W. The occurrence of Fascioloides Magna (Bassi, 1875) in the wild cervid population in the Lower Silesian Wilderness—Epidemiological and pathological aspects. J. Vet. Res. 2022, 66, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Nagy, E.; Jócsák, I.; Csivincsik, Á.; Zsolnai, A.; Halász, T.; Nyúl, A.; Plucsinszki, Z.; Simon, T.; Szabó, S.; Turbók, J.; et al. Establishment of Fascioloides magna in a new region of Hungary: Case report. Parasitol. Res. 2018, 117, 3683–3687. [Google Scholar] [CrossRef]
- Swales, W.E. The life cycle of Fascioloides magna (Bassi, 1875), the large liver fluke of ruminants, in Canada: With observations on the bionomics of the larval stages and the intermediate hosts, pathology of Fascioloidiasis magna, and control measures. Can. J. Res. 1935, 12, 177–215. [Google Scholar] [CrossRef]
- Campbell, W.C. Notes on the egg and miracidium of Fascioloides magna, (Trematoda). Trans. Am. Micros. Soc. 1961, 28, 308–319. [Google Scholar] [CrossRef]
- Reiczigel, J.; Marozzi, M.; Fábián, I.; Rózsa, L. Biostatistics for parasitologists—A primer to Quantitative Parasitology. Trends Parasitol. 2019, 35, 277–281. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 27.0; IBM Corp: Armonk, NY, USA, 2020. [Google Scholar]
- Trouvé, S.; Renaud, F.; Durand, P.; Jourdane, J. Selfing and outcrossing in a parasitic hermaphrodite helminth (Trematoda, Echinostomatidae). Heredity 1996, 77, 1–8. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Dalton, J.P.; Dufresne, P.J.; La Course, J.; Williams, D.J.L.; Hodkgkinson, J.; Paterson, S. The Fasciola hepatica genome: Gene duplication and polymorphism reveals adaptation to the host environment and the capacity for rapid evolution. Genome Biol. 2015, 16, 71. [Google Scholar] [CrossRef]
- Shury, T.K.; Pybus, M.J.; Nation, N.; Cool, N.L.; Rettie, W.J. Fascioloides magna in moose (Alces alces) from Elk Island National Park, Alberta. Vet. Pathol. 2019, 56, 476–485. [Google Scholar] [CrossRef]
- Houszka, M.; Piekarska, J.; Podkowik, M.; Gorczykowski, M.; Bania, J. Morphology and molecular study of Fascioloides magna—A growing threat to cervids (Cervidae) in Poland. J. Ver. Res. 2016, 60, 435–439. [Google Scholar] [CrossRef]
- Beesley, N.J.; Williams, D.J.; Paterson, S.; Hodgkinson, J. Fasciola hepatica demonstrates high levels of genetic diversity, a lack of population structure and high gene flow: Possible implications for drug resistance. Int. J. Parasitol. 2017, 47, 11–20. [Google Scholar] [CrossRef]
- Gowler, C.D.; Essington, H.; O’Brien, B.; Shaw, C.L.; Bilich, R.W.; Clay, P.A.; Duffy, M.A. Virulence evolution during a naturally occurring parasite outbreak. Evol. Ecol. 2023, 37, 113–129. [Google Scholar] [CrossRef]
- Leggett, H.C.; Buckling, A.; Long, G.H.; Boots, M. Generalism and the evolution of parasite virulence. Trends Ecol. Ecol. 2013, 28, 592–596. [Google Scholar] [CrossRef]
- Woolhouse, M.E.; Taylor, L.H.; Haydon, D.T. Population biology of multihost pathogens. Science 2001, 292, 1109–1112. [Google Scholar] [CrossRef]
- Martcheva, M.; Tuncer, N.; Kim, Y. On the principle of host evolution in host-pathogen interactions. J. Biol. Dyn. 2017, 11, 102–119. [Google Scholar] [CrossRef]
- Živković, D.; John, S.; Verin, M.; Stephan, W.; Tellier, A. neutral genomic signatures of host-parasite coevolution. BMC Evol. Biol. 2019, 19, 230. [Google Scholar] [CrossRef]
- Buckingham, L.J.; Ashby, B. Coevolutionary theory of hosts and parasites. J. Evol. Biol. 2022, 35, 205–224. [Google Scholar] [CrossRef]
- Regoes, R.R.; Nowak, M.A.; Bonhoeffer, S. Evolution of virulence in a heterogeneous host population. Evolution 2000, 54, 64–71. [Google Scholar]
- White, P.S.; Choi, A.; Pandey, R.; Menezes, A.; Penley, M.; Gibson, A.K.; de Roode, J.; Morran, L. Host heterogeneity mitigates virulence evolution. Biol. Lett. 2020, 16, 20190744. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Category | Main Characteristics |
|---|---|
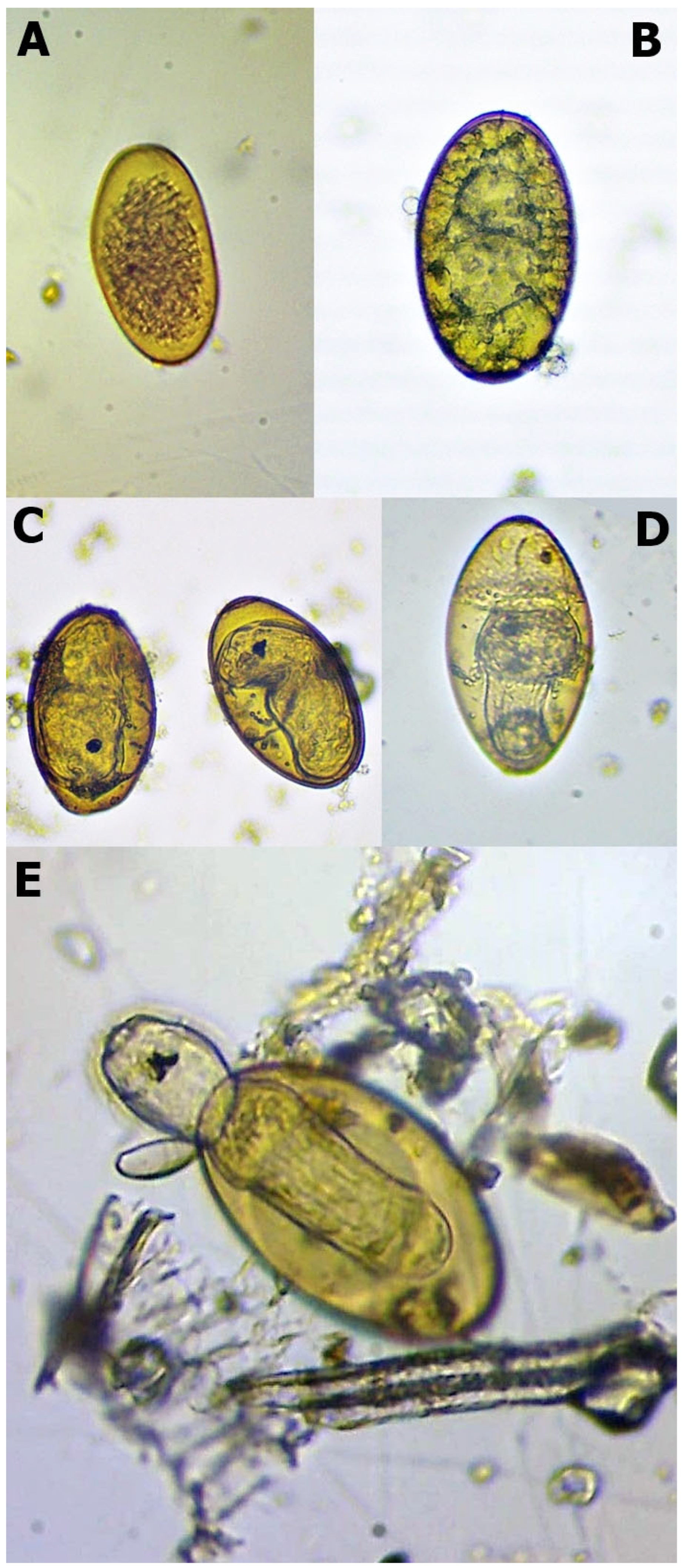
| Unembryonated egg | No developmental processing, the eggs usually seem empty or damaged (Figure 1A) |
| Embryonated egg | Embryo position in the egg’s centre, embryo formation visible (Figure 1B) |
| Eye-spot development | Movement often observable eye-spots appeared (Figure 1C) |
| Pre-hatching miracidium | Intensive movement increased mucoid plug (Figure 1D) |
| Hatching | Opened operculum miracidia outside the egg (Figure 1E) |
| One Fluke | Two Flukes | Three Flukes | |
|---|---|---|---|
| red deer | 17 (4) * | 46 (45) | 7 (7) |
| roe deer | 1 (1) | 2 (1) | 0 |
| Total Egg | Embryonated | Eye-Spotted | Pre-Hatching | Hatching | |
|---|---|---|---|---|---|
| RED-PC1 | 212 | 80.2% (170) * | 64.2% (136) | 64.2% (136) | 52.8% (1112) |
| RED-PC2 | 232 | 84.5% (196) | 70.3% (163) | 70.3% (163) | 60.3% (140) |
| ROE-PC1 | 83 | 36.9% (31) | 14.3% (12) | 14.3% (12) | 4.8% (4) |
| ROE-PC2 | 278 | 44.8% (60) | 20.1% (27) | 20.1% (27) | 9.7% (13) |
| RED-PC2 | ROE-PC1 | ROE-PC2 | |
|---|---|---|---|
| RED-PC1 * | 0.059 ** | <0.0001 | <0.0001 |
| RED-PC2 | <0.0001 | <0.0001 | |
| ROE-PC1 | 0.056 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halász, T.; Tari, T.; Nagy, E.; Nagy, G.; Csivincsik, Á. Hatchability of Fascioloides magna Eggs in Cervids. Pathogens 2023, 12, 741. https://doi.org/10.3390/pathogens12050741
Halász T, Tari T, Nagy E, Nagy G, Csivincsik Á. Hatchability of Fascioloides magna Eggs in Cervids. Pathogens. 2023; 12(5):741. https://doi.org/10.3390/pathogens12050741
Chicago/Turabian StyleHalász, Tibor, Tamás Tari, Eszter Nagy, Gábor Nagy, and Ágnes Csivincsik. 2023. "Hatchability of Fascioloides magna Eggs in Cervids" Pathogens 12, no. 5: 741. https://doi.org/10.3390/pathogens12050741
APA StyleHalász, T., Tari, T., Nagy, E., Nagy, G., & Csivincsik, Á. (2023). Hatchability of Fascioloides magna Eggs in Cervids. Pathogens, 12(5), 741. https://doi.org/10.3390/pathogens12050741