
Blocking BAFF Alleviates Hepatic Fibrosis in Schistosoma japonicum-Infected Mice
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Schistosome and Animals
2.2. Mice Infected with S. japonicum
2.3. S. japonicum Infected Mice Treated with Anti-BAFF Antibody
2.4. Determination Hydroxyproline Content in the Liver of Mice
2.5. HE and Masson Collagen Staining
2.6. Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Flow Cytometry
2.8. Statistical Analysis
3. Results
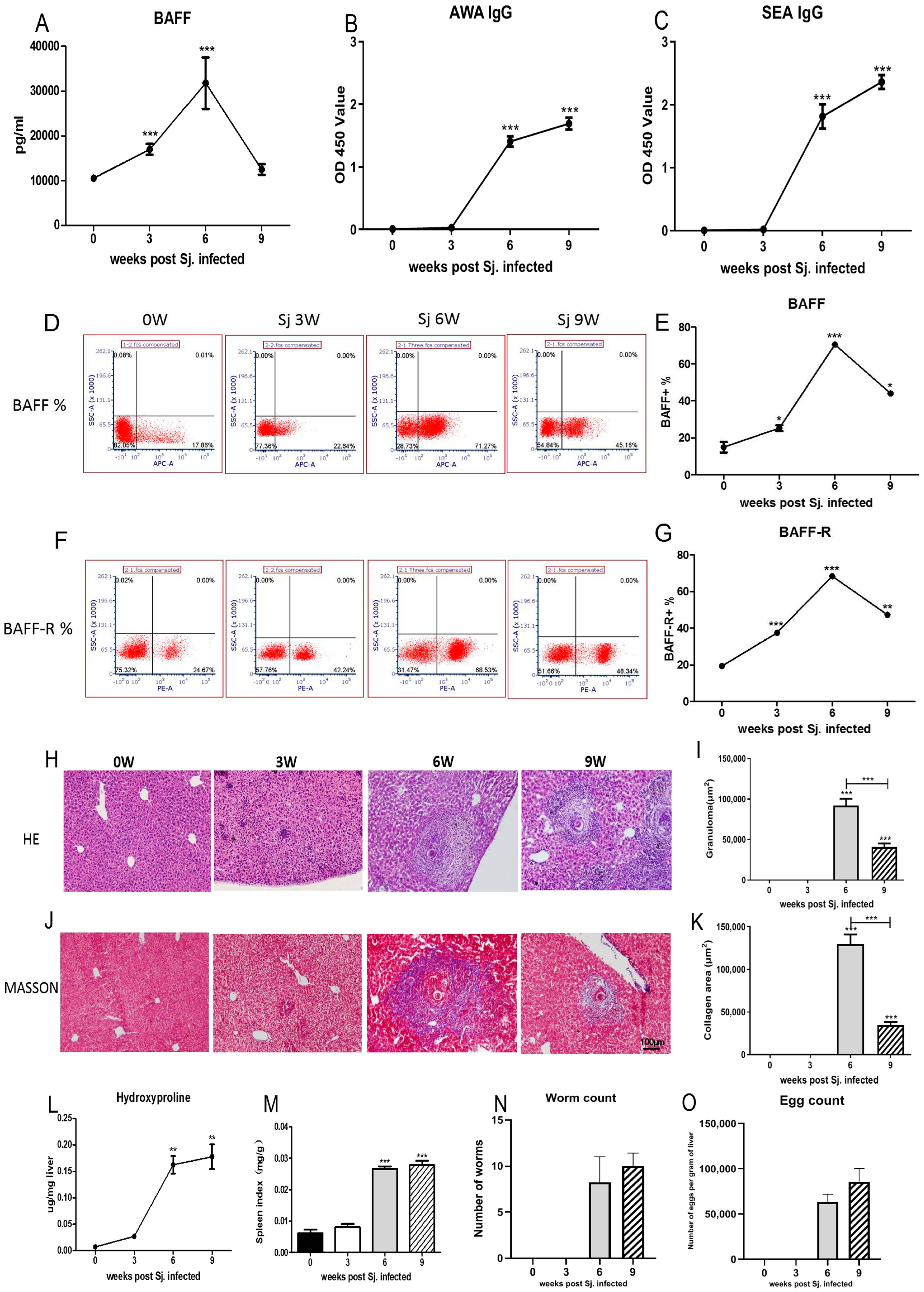
3.1. The Dynamic Changes in BAFF, BAFF-R Were Related to the Pathological Course of Schistosomiasis
3.2. Anti-BAFF Treatment Inhibited Hepatic Fibrosis of Mice Infected with S. japonicum
3.3. Anti-BAFF Treatment Decreased the Specific IgG against S. japonicum Antigens and Fibrosis-Related Cytokines Level in Serum
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.-J.; Xu, Y.-X.; Hu, Y.; Shen, Y.-J.; Li, P.; Zhou, H.-J.; Cao, J.-P. Destructive effect of eggs of Schistosoma japonicum on spleens in C57BL/6 mice. Zhongguo Xue Xi Chong Bing Fang Zhi Za Zhi 2011, 23, 285–287. [Google Scholar] [PubMed]
- Song, L.-J.; Yin, X.-R.; Mu, S.-S.; Li, J.-H.; Gao, H.; Zhang, Y.; Dong, P.-P.; Mei, C.-J.; Hua, Z.-C. The Differential and Dynamic Progression of Hepatic Inflammation and Immune Responses During Liver Fibrosis Induced by Schistosoma japonicum or Carbon Tetrachloride in Mice. Front. Immunol. 2020, 11, 570524. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.D.; Zhou, J.; Chen, E.Q. Molecular Mechanisms and Potential New Therapeutic Drugs for Liver Fibrosis. Front. Pharmacol. 2022, 13, 787748. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Bohdziewicz, A.; Pawlik, K.K.; Maciejewska, M.; Sikora, M.; Alda-Malicka, R.; Czuwara, J.; Rudnicka, L. Future Treatment Options in Systemic Sclerosis—Potential Targets and Ongoing Clinical Trials. J. Clin. Med. 2022, 11, 1310. [Google Scholar] [CrossRef]
- Zheng, B.; Zhang, J.; Chen, H.; Nie, H.; Miller, H.; Gong, Q.; Liu, C. T Lymphocyte-Mediated Liver Immunopathology of Schistosomiasis. Front. Immunol. 2020, 11, 61. [Google Scholar] [CrossRef]
- Sun, Y.J.; Li, Z.Q.; Lu, F.L. Research progress on the immunopathological mechanism of Schistosoma japonicum egg-induced granuloma. Chin. J. Parasit. Dis. Dec. 2019, 37, 713–717+722. [Google Scholar]
- Ji, F.; Liu, Z.; Cao, J.; Li, N.; Liu, Z.; Zuo, J.; Chen, Y.; Wang, X.; Sun, J. B cell response is required for granuloma formation in the early infection of Schistosoma japonicum. PLoS ONE 2008, 3, e1724. [Google Scholar] [CrossRef]
- Xiao, J.; Guan, F.; Sun, L.; Zhang, Y.; Zhang, X.; Lu, S.; Liu, W. B cells induced by Schistosoma japonicum infection display diverse regulatory phenotypes and modulate CD4+ T cell response. Parasites Vectors 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Yong, L.; Tang, Y.; Ren, C.; Liu, M.; Shen, J.; Hou, X. B1 cells protect against Schistosoma japonicum–induced liver inflammation and fibrosis by controlling monocyte infiltration. PLoS Negl. Trop. Dis. 2019, 13, e0007474. [Google Scholar] [CrossRef]
- Brown, M.; O’Reilly, S. The immunopathogenesis of fibrosis in systemic sclerosis. Clin. Exp. Immunol. 2019, 195, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.C.; Zhong, J.; Xu, J.F. Regulatory B cells in infectious disease (Review). Mol. Med. Rep. 2017, 16, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Yehudai-Ofir, D.; Henig, I.; Zuckerman, T. Aberrant B cells, autoimmunity and the benefit of targeting B cells in chronic graft-versus-host disease. Autoimmun. Rev. 2020, 19, 102493. [Google Scholar] [CrossRef]
- Xu, H.; Dong, P.; Ma, X.; Song, D.; Xue, D.; Xu, R.; Lu, H.; He, X. B cell-activating factor regulates the survival of B lymphocytes infected with human cytomegalovirus. Immunol. Lett. 2017, 187, 1–6. [Google Scholar] [CrossRef]
- Hu, S.; Wang, R.; Zhang, M.; Liu, K.; Tao, J.; Tai, Y.; Zhou, W.; Wang, Q.; Wei, W. BAFF promotes T cell activation through the BAFF-BAFF-R-PI3K-Akt signaling pathway. Biomed. Pharmacother. 2019, 114, 108796. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.-D.; Wang, Y.-H.; Fang, W.-F.; Hsiao, C.-J.; Chagnaadorj, A.; Lin, Y.-F.; Tang, K.-T.; Cheng, C.-W. Serum BAFF and thyroid autoantibodies in autoimmune thyroid disease. Clin. Chim. Acta 2016, 462, 96–102. [Google Scholar] [CrossRef]
- Yarchoan, M.; Ho, W.J.; Mohan, A.; Shah, Y.; Vithayathil, T.; Leatherman, J.; Dennison, L.; Zaidi, N.; Ganguly, S.; Woolman, S.; et al. Effects of B cell–activating factor on tumor immunity. JCI Insight 2020, 5, e136417. [Google Scholar] [CrossRef]
- Duan, J.H.; Jiang, Y.; Mu, H.; Tang, Z.Q. Expression of BAFF and BR3 in patients with systemic lupus erythematosus. Braz. J. Med. Biol. Res. 2016, 49. [Google Scholar] [CrossRef]
- Tanaka, Y. State-of-the-art treatment of systemic lupus erythematosus. Int. J. Rheum. Dis. 2020, 23, 465–471. [Google Scholar] [CrossRef]
- Wei, F.; Chang, Y.; Wei, W. The role of BAFF in the progression of rheumatoid arthritis. Cytokine 2015, 76, 537–544. [Google Scholar] [CrossRef]
- Kannel, K.; Alnek, K.; Vahter, L.; Gross-Paju, K.; Uibo, R.; Kisand, K.V. Changes in blood B cell-activating factor (BAFF) levels in multiple sclerosis: A sign of treatment outcome. PLoS ONE 2015, 10, e0143393. [Google Scholar] [CrossRef] [PubMed]
- Mo, F.; Luo, Y.; Yan, Y.; Li, J.; Lai, S.; Wu, W. Are activated B cells involved in the process of myocardial fibrosis after acute myocardial infarction? An in vivo experiment. BMC Cardiovasc. Disord. 2021, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Lian, H.J.; Li, S.; Fang, C.L.; Huang, H.; Xu, Z.J. The role of B cell activating factor in the differential diagnosis of usual interstitial pneumonia. Zhonghua Jiehe He Huxi Zazhi 2018, 41, 544–550. [Google Scholar] [PubMed]
- Xu, H.; Song, D.; Xu, R.; He, X. BAFF signaling drives interstitial transformation of mouse renal tubular epithelial cells in a Pin1-dependent manner. Cell Dev. Biol. Anim. 2021, 57, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Liossis, S.C.; Staveri, C. The Role of B Cells in Scleroderma Lung Disease Pathogenesis. Front. Med. 2022, 9, 936182. [Google Scholar] [CrossRef]
- Thapa, M.; Tedesco, D.; Gumber, S.; Elrod, E.J.; Han, J.H.; Kitchens, W.H.; Magliocca, J.F.; Adams, A.B.; Grakoui, A. Blockade of BAFF Reshapes the Hepatic B Cell Receptor Repertoire and Attenuates Autoantibody Production in Cholestatic Liver Disease. J. Immunol. 2020, 204, 3117–3128. [Google Scholar] [CrossRef]
- Nakamura, Y.; Abe, M.; Kawasaki, K.; Miyake, T.; Watanabe, T.; Yoshida, O.; Hirooka, M.; Matsuura, B.; Hiasa, Y. Depletion of B cell-activating factor attenuates hepatic fat accumulation in a murine model of nonalcoholic fatty liver disease. Sci. Rep. 2019, 9, 977. [Google Scholar] [CrossRef]
- Kim, N.D.; Luster, A.D. To B or not to B—That is the question for myocardial infarction. Nat. Med. 2013, 19, 1208–1210. [Google Scholar] [CrossRef]
- Zeng, M.; Zhang, Y.; Zhang, X.; Zhang, W.; Yu, Q.; Zeng, W.; Ma, D.; Gan, J.; Yang, Z.; Jiang, X. Two birds with one stone: YQSSF regulates both proliferation and apoptosis of bone marrow cells to relieve chemotherapy-induced myelosuppression. J. Ethnopharmacol. 2022, 289, 115028. [Google Scholar]
- Zhang, Y.; Tao, M.; Chen, C.; Zhao, X.; Feng, Q.; Chen, G.; Fu, Y. BAFF Blockade Attenuates DSS-Induced Chronic Colitis via Inhibiting NLRP3 Inflammasome and NF-κB Activation. Front. Immunol. 2022, 13, 783254. [Google Scholar] [CrossRef]
- Zhou, W.; Yang, Y.; Mei, C.; Dong, P.; Mu, S.; Wu, H.; Zhou, Y.; Zheng, Y.; Guo, F.; Yang, J.Q. Inhibition of Rho-kinase downregulates Th17 cells and ameliorates hepatic fibrosis by Schistosoma japonicum infection. Cells 2019, 8, 1262. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Wang, Y.; Yi, X.; Wang, S. Identification of antigens in circulating immune complexes of Schistosomiasis japonica. Hunan Yi Ke Da Xue Xue Bao 1997, 22, 377–380. [Google Scholar] [PubMed]
- Yang, L.L.; Lv, Z.Y.; Hu, S.M.; He, S.J.; Li, Z.Y.; Zhang, S.M.; Zheng, H.Q.; Li, M.T.; Yu, X.B.; Fung, M.C.; et al. Schistosoma japonicum: Proteomics analysis of differentially expressed proteins from ultraviolet-attenuated cercariae compared to normal cercariae. Parasitol. Res. 2009, 105, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Brindley, P.J.; Bethony, J.M.; King, C.H.; Pearce, E.J.; Jacobson, J. Helminth infections: The great neglected tropical diseases. J. Clin. Investig. 2008, 118, 1311–1321. [Google Scholar] [CrossRef]
- Wilson, R.A. Schistosomiasis then and now: What has changed in the last 100 years? Parasitology 2020, 147, 507–515. [Google Scholar] [CrossRef]
- Matsushita, T.; Fujimoto, M.; Hasegawa, M.; Matsushita, Y.; Komura, K.; Ogawa, F.; Watanabe, R.; Takehara, K.; Sato, S. BAFF Antagonist Attenuates the Development of Skin Fibrosis in Tight-Skin Mice. J. Investig. Dermatol. 2007, 127, 2772–2780. [Google Scholar] [CrossRef]
- Mwinzi, P.N.; Ganley-Leal, L.; Black, C.L.; Evan Secor, W.; Karanja, D.M.; Colley, D.G. Circulating CD23+ B cell subset correlates with the development of resistance to Schistosoma mansoni reinfection in occupationally exposed adults who have undergone multiple treatments. J. Infect. Dis. 2009, 199, 272–279. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiong, D.-H.; Li, Y.; Xu, G.; Zhang, B.; Liu, Y.; Zhang, S.; Huang, Q.; Chen, S.; Zeng, F.; et al. Schistosoma japonicum Infection in Treg-Specific USP21 Knockout Mice. J. Immunol. Res. 2021, 2021, 6613162. [Google Scholar] [CrossRef]
- Shen, P.; Fillatreau, S. Antibody-independent functions of B cells: A focus on cytokines. Nat. Rev. Immunol. 2015, 15, 441–451. [Google Scholar] [CrossRef]
- Catalán, D.; Mansilla, M.A.; Ferrier, A.; Soto, L.; Oleinika, K.; Aguillón, J.C.; Aravena, O. Immunosuppressive Mechanisms of Regulatory B Cells. Front. Immunol. 2021, 12, 611795. [Google Scholar] [CrossRef]
- Barbosa, R.R.; Silva, S.P.; Silva, S.L.; Melo, A.C.; Pedro, E.; Barbosa, M.A.D.C.P.; Santos, M.C.; Victorino, R.; Sousa, A.E. Primary B-Cell Deficiencies Reveal a Link between Human IL-17-Producing CD4 T-Cell Homeostasis and B-Cell Differentiation. PLoS ONE 2011, 6, e22848. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Zhuo, Y.; He, F.; Lv, L.; Xing, M.; Guo, Y.; Zhang, Y.; Liu, J.; Li, Y.; Bai, T.; et al. Longitudinal Immune Profiling Highlights CD4+ T Cell Exhaustion Correlated with Liver Fibrosis in Schistosoma japonicum Infection. J. Immunol. 2023, 210, 82–95. [Google Scholar] [CrossRef] [PubMed]
- Lund, F.E.; Garvy, B.A.; Randall, T.D.; Harris, D.P. Regulatory roles for cytokine-producing B cells in infection and autoimmune disease. Curr. Dir. Autoimmun. 2005, 8, 25–54. [Google Scholar] [PubMed]
- Mizoguchi, A.; Bhan, A.K. A case for regulatory B cells. J. Immunol. 2006, 176, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Karnowski, A.; Chevrier, S.; Belz, G.T.; Mount, A.; Emslie, D.; D’Costa, K.; Tarlinton, D.M.; Kallies, A.; Corcoran, L.M. B and T cells collaborate in antiviral responses via IL-6, IL-21, and transcriptional activator and coactivator, Oct2 and OBF-1. J. Exp. Med. 2012, 209, 2049–2064. [Google Scholar] [CrossRef]
- Li, M.; Wang, H.; Ni, Y.; Li, C.; Xu, X.; Chang, H.; Xu, Z.; Hou, M.; Ji, M. Helminth-induced CD9+ B-cell subset alleviates obesity-associated inflammation via IL-10 production. Int. J. Parasitol. 2021, 52, 111–123. [Google Scholar] [CrossRef]
- Haeberlein, S.; Obieglo, K.; Ozir-Fazalalikhan, A.; Chayé, M.A.M.; Veninga, H.; Van Der Vlugt, L.E.P.M.; Voskamp, A.; Boon, L.; Haan, J.M.M.D.; Westerhof, L.B.; et al. Schistosome egg antigens, including the glycoprotein IPSE/alpha-1, trigger the development of regulatory B cells. PLoS Pathog. 2017, 13, e1006539. [Google Scholar] [CrossRef]
- Wahl, S.M.; Frazier-Jessen, M.; Jin, W.W.; Kopp, J.B.; Sher, A.; Cheever, A.W. Cytokine regulation of schistosome-induced granuloma and fibrosis. Kidney Int. 1997, 51, 1370–1375. [Google Scholar] [CrossRef]
- Phanish, M.K.; Wahab, N.A.; Colville-Nash, P.; Hendry, B.M.; Dockrell, M.E. The differential role of Smad2 and Smad3 in the regulation of pro-fibrotic TGFbeta1 responses in human proximal-tubule epithelial cells. Biochem. J. 2016, 393, 601–607. [Google Scholar] [CrossRef]
- Wang, S.; Wilkes, M.C.; Leof, E.B.; Hirschberg, R. Noncanonical TGF-beta pathways, mTORC1 and Abl, in renal interstitial fibrogenesis. Am. J. Physiol. Ren. Physiol. 2010, 298, F142–F149. [Google Scholar]
- Doenhoff, M.; Musallam, R.; Bain, J.; McGregor, A. Studies on the host-parasite relationship in Schistosoma mansoni-infected mice: The immunological dependence of parasite egg excretion. Immunology 1978, 35, 771–778. [Google Scholar] [PubMed]
- Lamb, E.W.; Crow, E.T.; Lim, K.; Liang, Y.-S.; Lewis, F.A.; Davies, S.J. Conservation of CD4+ T cell-dependent developmental mechanisms in the blood fluke pathogens of humans. Int. J. Parasitol. 2007, 37, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Blank, R.B.; Lamb, E.W.; Tocheva, A.S.; Crow, E.T.; Lim, K.C.; McKerrow, J.H.; Davies, S.J. The common γ chain cytokines interleukin (IL)–2 and IL-7 indirectly modulate blood fluke development via effects on CD4+ T cells. J. Infect. Dis. 2006, 194, 1609–1616. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, P.; Mei, C.; Yang, Y.; Zhou, Y.; Xu, Y.; Song, L.; Yu, C. Blocking BAFF Alleviates Hepatic Fibrosis in Schistosoma japonicum-Infected Mice. Pathogens 2023, 12, 793. https://doi.org/10.3390/pathogens12060793
Dong P, Mei C, Yang Y, Zhou Y, Xu Y, Song L, Yu C. Blocking BAFF Alleviates Hepatic Fibrosis in Schistosoma japonicum-Infected Mice. Pathogens. 2023; 12(6):793. https://doi.org/10.3390/pathogens12060793
Chicago/Turabian StyleDong, Panpan, Congjin Mei, Yingying Yang, Yonghua Zhou, Yongliang Xu, Lijun Song, and Chuanxin Yu. 2023. "Blocking BAFF Alleviates Hepatic Fibrosis in Schistosoma japonicum-Infected Mice" Pathogens 12, no. 6: 793. https://doi.org/10.3390/pathogens12060793