Author Contributions
Conceptualization, I.A., D.S.C.B. and C.S.; data curation, I.A., T.H.T., D.S.C.B., M.C., M.L.Y.W. and S.A.; formal analysis, I.A., D.S.C.B. and C.S., I.A., T.H.T., D.S.C.B., M.C. and S.A.; methodology, T.H.T., D.S.C.B. and C.S.; writing—original draft, D.S.C.B. and C.S.; writing—review and editing, I.A. and C.S. All authors have read and agreed to the published version of the manuscript.
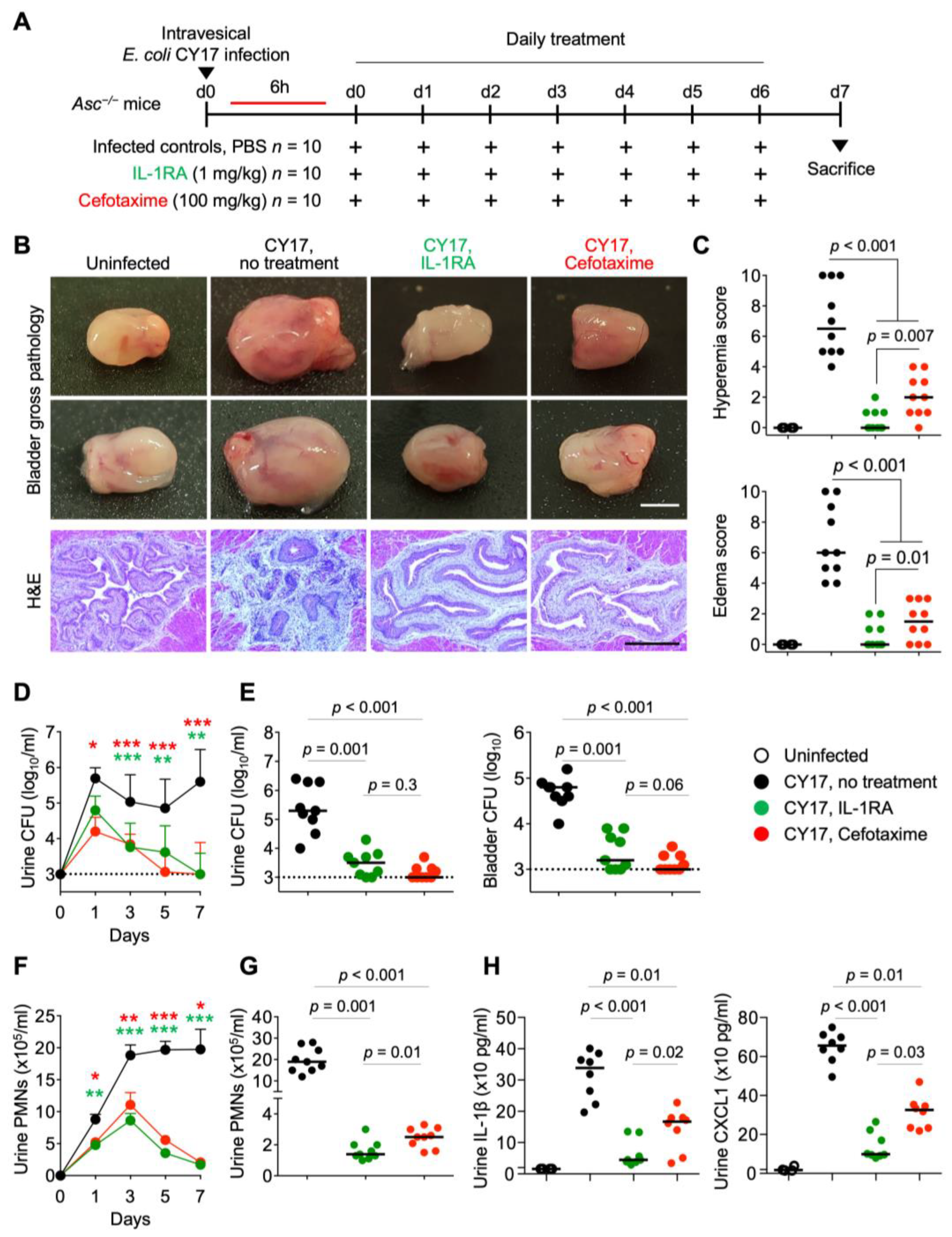
Figure 1.
Efficacy of IL-1RA treatment in acute cystitis compared to cefotaxime. (A) Schematic of the infection and treatment protocol. Genetically predisposed Asc−/− mice were infected with the acute cystitis isolate E. coli CY17 (antibiotic sensitive) at 108 CFU. Six hours post-infection, mice were treated i.p. with IL-1RA (anakinra, 1 mg/kg) or cefotaxime (100 mg/kg). Treatment was continued daily, until day 7, when mice were sacrificed (n = 9 mice per group from 2 experiments). (B) Treatment alleviated severe cystitis in CY17-infected mice. Differences in gross bladder pathology between treated (IL-1RA or cefotaxime) and untreated Asc−/− mice. (C) Gross pathology quantified by edema and hyperemia scores was reduced in treated mice compared to untreated mice. (D–G) Treatment accelerated bacterial clearance and reduced inflammation, quantified as neutrophil recruitment. (D) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, and 7, post-infection. (E) Bacterial counts in urine and bladder tissue (day 7). (F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, and 7, post-infection. (G) Quantification of neutrophils in urine (day 7). (H) Effects of treatment on inflammatory mediators IL-1β and CXCL1 in urine at day 7. Representative bladder images (B), scale bar = 2 mm. Data are presented as individual mice, and the horizontal lines represent the median (C,E,G,H) and mean ± SEM (D,F). Data were analyzed using the Kruskal–Wallis test, * = p < 0.05; ** = p < 0.01; *** = p < 0.001.
Figure 1.
Efficacy of IL-1RA treatment in acute cystitis compared to cefotaxime. (A) Schematic of the infection and treatment protocol. Genetically predisposed Asc−/− mice were infected with the acute cystitis isolate E. coli CY17 (antibiotic sensitive) at 108 CFU. Six hours post-infection, mice were treated i.p. with IL-1RA (anakinra, 1 mg/kg) or cefotaxime (100 mg/kg). Treatment was continued daily, until day 7, when mice were sacrificed (n = 9 mice per group from 2 experiments). (B) Treatment alleviated severe cystitis in CY17-infected mice. Differences in gross bladder pathology between treated (IL-1RA or cefotaxime) and untreated Asc−/− mice. (C) Gross pathology quantified by edema and hyperemia scores was reduced in treated mice compared to untreated mice. (D–G) Treatment accelerated bacterial clearance and reduced inflammation, quantified as neutrophil recruitment. (D) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, and 7, post-infection. (E) Bacterial counts in urine and bladder tissue (day 7). (F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, and 7, post-infection. (G) Quantification of neutrophils in urine (day 7). (H) Effects of treatment on inflammatory mediators IL-1β and CXCL1 in urine at day 7. Representative bladder images (B), scale bar = 2 mm. Data are presented as individual mice, and the horizontal lines represent the median (C,E,G,H) and mean ± SEM (D,F). Data were analyzed using the Kruskal–Wallis test, * = p < 0.05; ** = p < 0.01; *** = p < 0.001.
![Pathogens 13 00042 g001]()
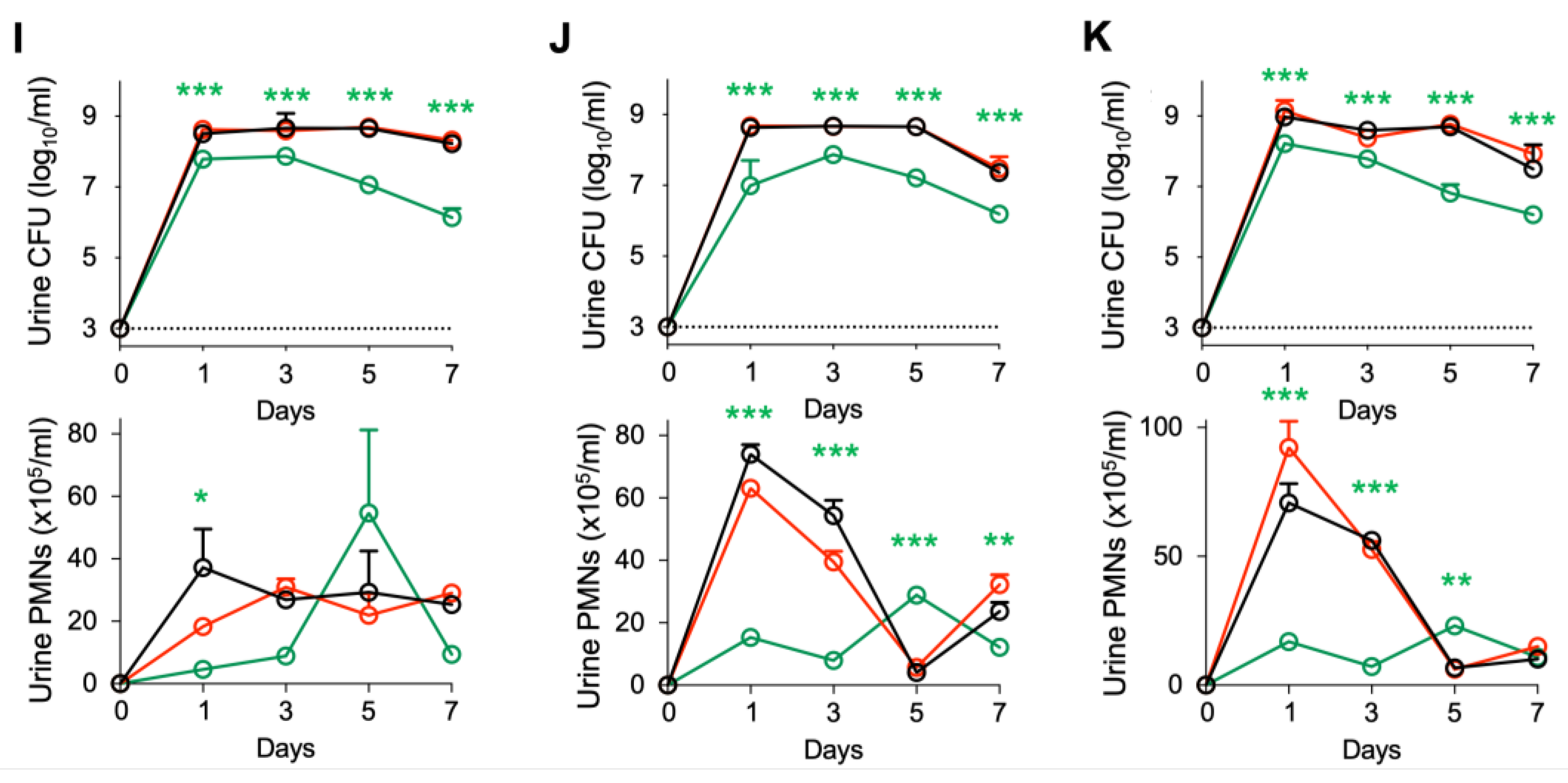
Figure 2.
Efficacy of IL-1RA treatment in
Asc−/− mice infected with ESBL strains.
Asc−/− mice were infected with each of four ESBL-producing
E. coli isolates (10
8 CFU,
n = 16 mice from 3 experiments) and treated according to the protocol shown in
Figure 1A with IL1-RA (anakinra, 1 mg/kg,
n = 17 mice) or cefotaxime (100 mg/kg
n = 11 mice) daily for seven days. Data in A-H represent ESBL strain 4, data in I-K represent ESBL strains 1–3. (
A,
B) Therapeutic efficacy of IL-1RA defined by a loss of gross pathology, hyperemia, and edema. No treatment effect was seen in cefotaxime-treated mice. (
C–
E) Accelerated bacterial clearance in IL-1RA- but not cefotaxime-treated mice. (
C) Kinetics of infection. Mean log
10 CFU/mL urine is shown for days 1, 3, 5, and 7, post-infection. (
D) Quantification of bacteria in urine (day 7). (
E) Quantification of bacteria in whole bladder tissues (day 7). (
F,
G) Neutrophil recruitment inhibited by IL-1RA but not cefotaxime treatment. (
F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, and 7, post-infection. (
G) Quantification of neutrophils in urine (day 7). (
H) Inhibition of urine IL-1β responses by IL-1RA treatment but not by cefotaxime. (
I–
K) Kinetics of infection and neutrophil recruitment. Bacterial counts and neutrophil numbers in the urine of mice infected with ESBL strains 1–3. Data are presented as individual mice, and the horizontal lines represent the median (
B,
D,
E,
G,
H) and mean ± SEM (
C,
F,
I–
K). Data were analyzed using the Kruskal–Wallis test, * =
p < 0.05; ** =
p < 0.01; *** =
p < 0.001.
Figure 2.
Efficacy of IL-1RA treatment in
Asc−/− mice infected with ESBL strains.
Asc−/− mice were infected with each of four ESBL-producing
E. coli isolates (10
8 CFU,
n = 16 mice from 3 experiments) and treated according to the protocol shown in
Figure 1A with IL1-RA (anakinra, 1 mg/kg,
n = 17 mice) or cefotaxime (100 mg/kg
n = 11 mice) daily for seven days. Data in A-H represent ESBL strain 4, data in I-K represent ESBL strains 1–3. (
A,
B) Therapeutic efficacy of IL-1RA defined by a loss of gross pathology, hyperemia, and edema. No treatment effect was seen in cefotaxime-treated mice. (
C–
E) Accelerated bacterial clearance in IL-1RA- but not cefotaxime-treated mice. (
C) Kinetics of infection. Mean log
10 CFU/mL urine is shown for days 1, 3, 5, and 7, post-infection. (
D) Quantification of bacteria in urine (day 7). (
E) Quantification of bacteria in whole bladder tissues (day 7). (
F,
G) Neutrophil recruitment inhibited by IL-1RA but not cefotaxime treatment. (
F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, and 7, post-infection. (
G) Quantification of neutrophils in urine (day 7). (
H) Inhibition of urine IL-1β responses by IL-1RA treatment but not by cefotaxime. (
I–
K) Kinetics of infection and neutrophil recruitment. Bacterial counts and neutrophil numbers in the urine of mice infected with ESBL strains 1–3. Data are presented as individual mice, and the horizontal lines represent the median (
B,
D,
E,
G,
H) and mean ± SEM (
C,
F,
I–
K). Data were analyzed using the Kruskal–Wallis test, * =
p < 0.05; ** =
p < 0.01; *** =
p < 0.001.
![Pathogens 13 00042 g002a]()
![Pathogens 13 00042 g002b]()
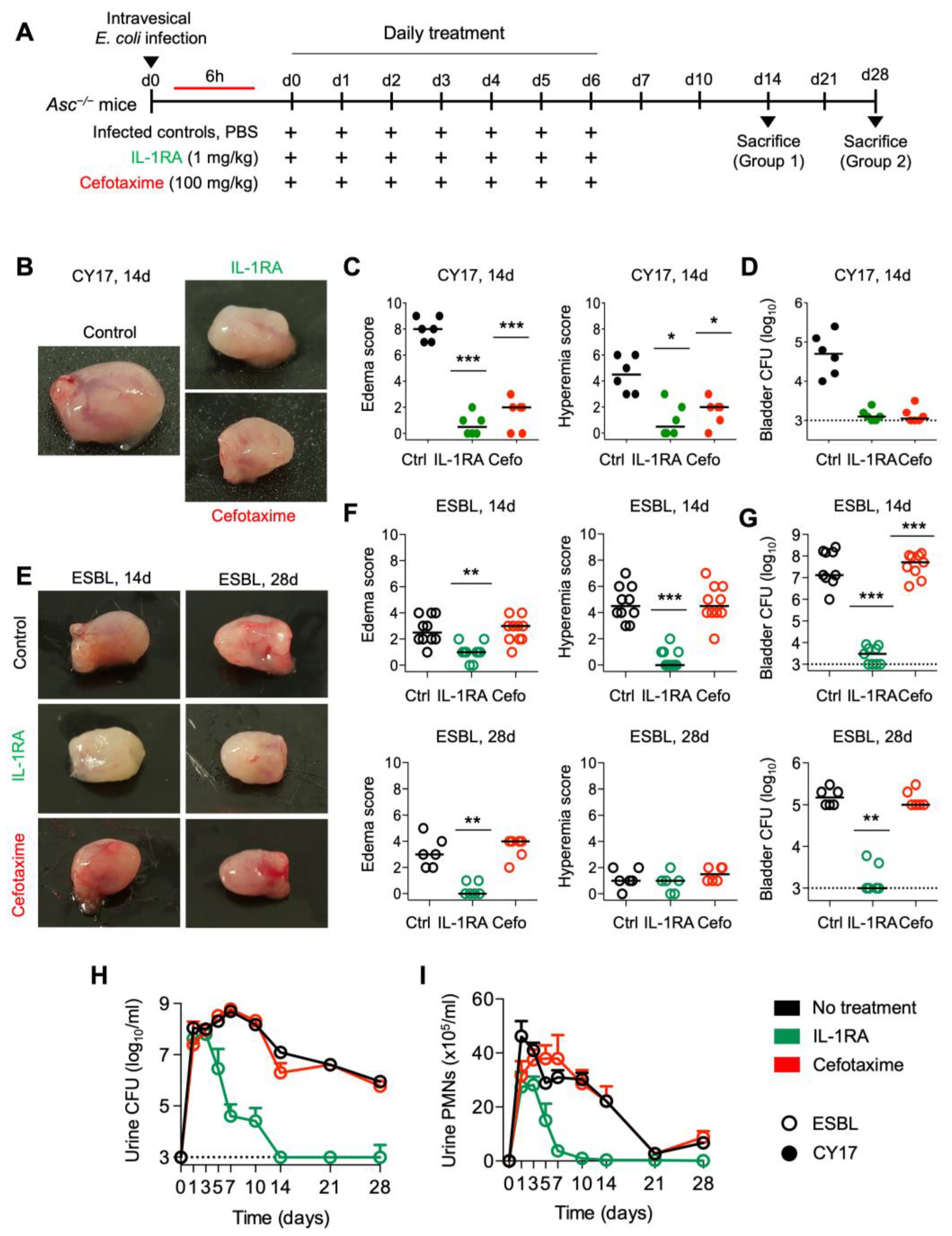
Figure 3.
Long-term protection by IL-1RA treatment in Asc−/− mice infected with CY17 and ESBL strains. (A) Schematic of the infection and treatment protocol. Asc−/− mice were infected with the CY17 or the ESBL4 strain at 108 CFU. Six hours post-infection, mice were treated i.p. with IL-1RA (anakinra, 1 mg/kg) or cefotaxime (100 mg/kg). Treatment was continued daily for 7 days, and the mice were followed without further intervention and sacrificed on day 14 or day 28 post-infection (n = 10 mice per group from 2 experiments). (B,C) IL-1RA treatment protected mice infected with CY17 from acute cystitis defined by low gross pathology scores and a lack of edema and hyperemia in treated compared to untreated mice. Cefotaxime treatment had similar treatment effect. (D) Bacterial counts for infection with the CY17 strain remained low in the treated groups. (E,F) IL-1RA treatment protected mice infected with the ESBL strain from acute cystitis, defined by low gross pathology scores and a lack of edema and hyperemia in treated compared to untreated mice. Cefotaxime treatment had no significant effect. (G) Log10 bacterial counts in bladder tissue 14- and 28-days post-infection. Bacterial counts for the ESBL strain remained low in the IL-1RA-treated groups. Cefotaxime treatment had no significant effect on the ESBL strain. (H) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, 7, 10, 14, 21, and 28 post-infection. IL-1RA treatment accelerated bacterial clearance. Cefotaxime had no significant effect. (I) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 10, 14, 21, and 28 post-infection. IL-1RA treatment reduced inflammation, quantified as neutrophil recruitment. Cefotaxime did not significantly reduce neutrophil numbers. Representative bladder images (B,E), scale bar = 2 mm. Data are presented as individual mice, and the horizontal line represents the median (C,D,F,G) and mean ± SEM (H,I). Data were analyzed using the Kruskal–Wallis test, * = p < 0.05; ** = p < 0.01; *** = p < 0.001.
Figure 3.
Long-term protection by IL-1RA treatment in Asc−/− mice infected with CY17 and ESBL strains. (A) Schematic of the infection and treatment protocol. Asc−/− mice were infected with the CY17 or the ESBL4 strain at 108 CFU. Six hours post-infection, mice were treated i.p. with IL-1RA (anakinra, 1 mg/kg) or cefotaxime (100 mg/kg). Treatment was continued daily for 7 days, and the mice were followed without further intervention and sacrificed on day 14 or day 28 post-infection (n = 10 mice per group from 2 experiments). (B,C) IL-1RA treatment protected mice infected with CY17 from acute cystitis defined by low gross pathology scores and a lack of edema and hyperemia in treated compared to untreated mice. Cefotaxime treatment had similar treatment effect. (D) Bacterial counts for infection with the CY17 strain remained low in the treated groups. (E,F) IL-1RA treatment protected mice infected with the ESBL strain from acute cystitis, defined by low gross pathology scores and a lack of edema and hyperemia in treated compared to untreated mice. Cefotaxime treatment had no significant effect. (G) Log10 bacterial counts in bladder tissue 14- and 28-days post-infection. Bacterial counts for the ESBL strain remained low in the IL-1RA-treated groups. Cefotaxime treatment had no significant effect on the ESBL strain. (H) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, 7, 10, 14, 21, and 28 post-infection. IL-1RA treatment accelerated bacterial clearance. Cefotaxime had no significant effect. (I) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 10, 14, 21, and 28 post-infection. IL-1RA treatment reduced inflammation, quantified as neutrophil recruitment. Cefotaxime did not significantly reduce neutrophil numbers. Representative bladder images (B,E), scale bar = 2 mm. Data are presented as individual mice, and the horizontal line represents the median (C,D,F,G) and mean ± SEM (H,I). Data were analyzed using the Kruskal–Wallis test, * = p < 0.05; ** = p < 0.01; *** = p < 0.001.
![Pathogens 13 00042 g003]()
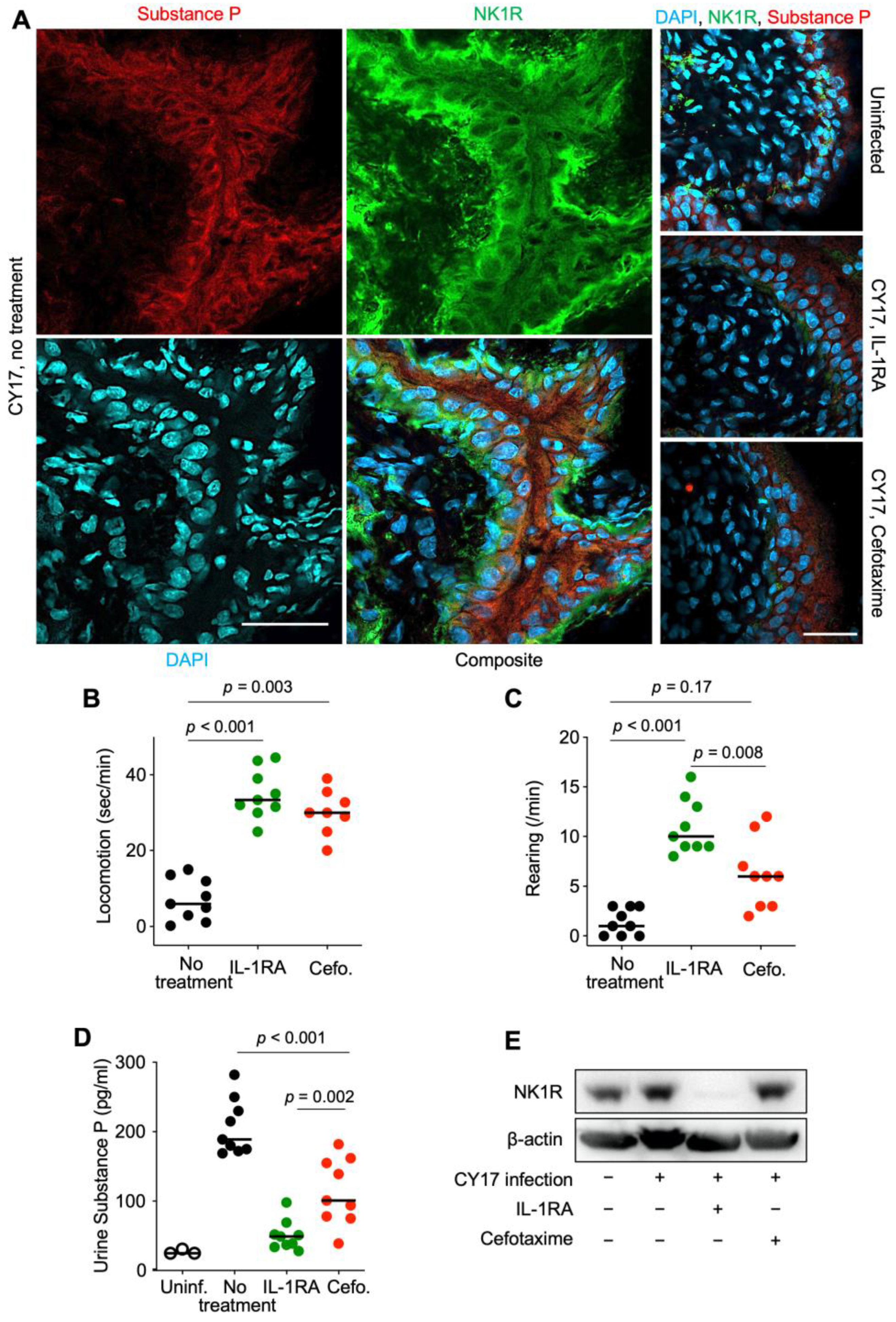
Figure 4.
Effects of IL-1RA treatment on pain sensing in nerve cells and pain behavior in infected mice. (A) Effects of treatment on the pain sensors substance P (red) and NK1R (green). Bladder tissue sections (day 7) were processed for immunohistochemistry and nuclei were counterstained using DAPI (blue). (B,C) Pain behavior was recorded on day 7 in CY17-infected mice and was quantified as (B) the time of movement (seconds/min) and (C) the amount of rearing behavior (events/min). (D) Urine substance P levels in uninfected mice, infected untreated Asc−/− mice, IL-1RA-treated, or cefotaxime-treated mice. (E) Western blot analysis of NK1R levels in whole bladder protein extracts (pooled samples from n = 4 mice per group). Representative images (A), n = 4 mice per group, scale bar = 2 mm. Data are presented as individual mice, and the horizontal line represents the median (B–D). Data were analyzed using the Kruskal–Wallis test.
Figure 4.
Effects of IL-1RA treatment on pain sensing in nerve cells and pain behavior in infected mice. (A) Effects of treatment on the pain sensors substance P (red) and NK1R (green). Bladder tissue sections (day 7) were processed for immunohistochemistry and nuclei were counterstained using DAPI (blue). (B,C) Pain behavior was recorded on day 7 in CY17-infected mice and was quantified as (B) the time of movement (seconds/min) and (C) the amount of rearing behavior (events/min). (D) Urine substance P levels in uninfected mice, infected untreated Asc−/− mice, IL-1RA-treated, or cefotaxime-treated mice. (E) Western blot analysis of NK1R levels in whole bladder protein extracts (pooled samples from n = 4 mice per group). Representative images (A), n = 4 mice per group, scale bar = 2 mm. Data are presented as individual mice, and the horizontal line represents the median (B–D). Data were analyzed using the Kruskal–Wallis test.
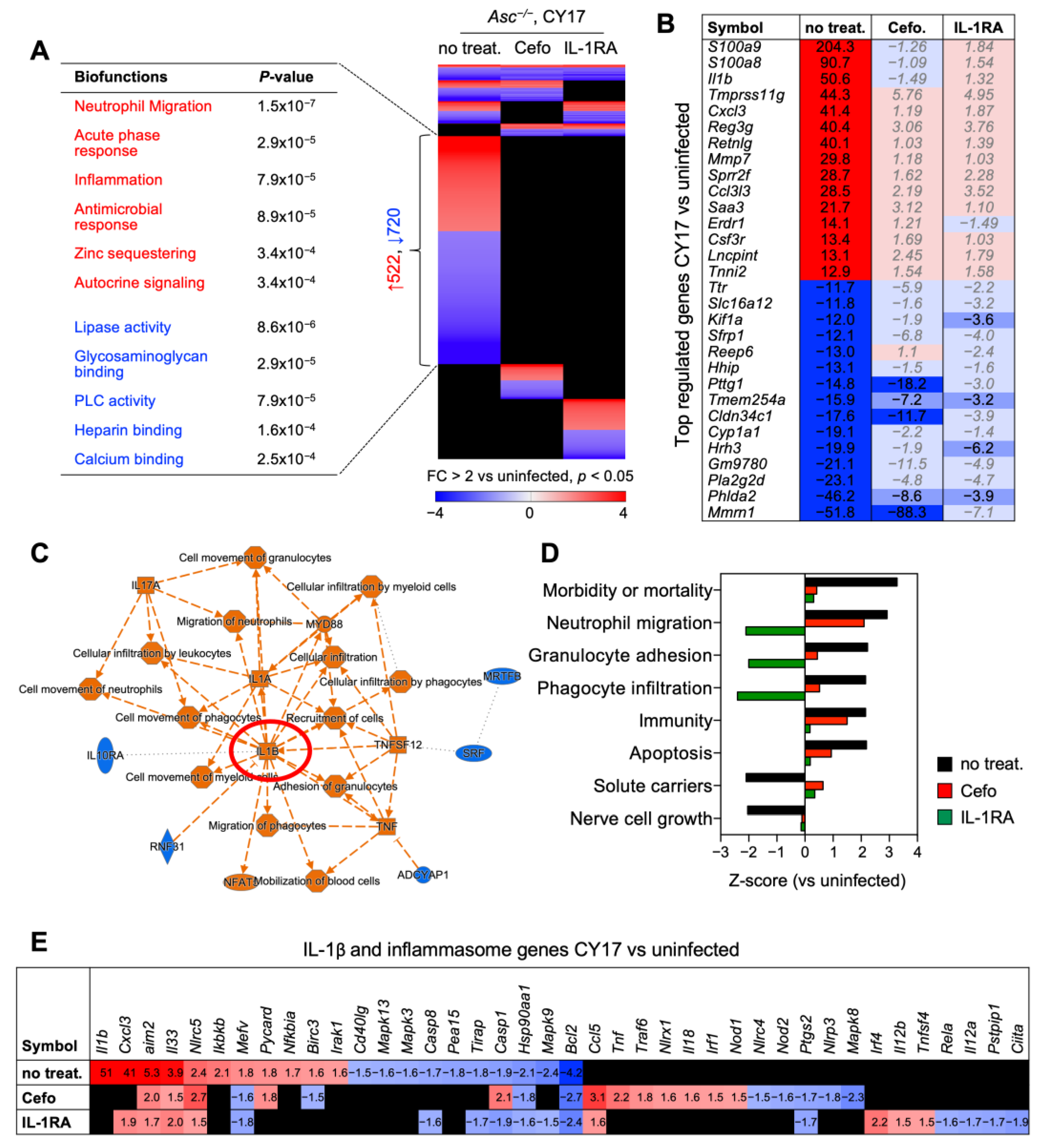
Figure 5.
IL-1RA treatment inhibits the expression of pro-inflammatory genes in infected Asc−/− mice. Whole bladder mRNA was subjected to transcriptomic analysis. (A) Gene expression profiles were compared between CY17-infected, untreated controls, and IL-1RA- or cefotaxime-treated mice (n = 2 mice per treatment group, n = 1 for uninfected control). Heat map of regulated genes in the treatment groups and CY17-infected controls compared to uninfected controls (cut-off FC ≥ 2, p < 0.05). Biofunctions of genes specific to CY17-infected controls included neutrophil migration, acute phase responses, inflammation, and antimicrobial responses. These responses were not activated in cefotaxime- or IL-1RA-treated mice (red = activated, blue = inhibited, black = not significantly regulated, compared to uninfected controls). (B) Top-regulated genes in CY17-infected mice treated with cefotaxime or IL-1RA compared to untreated controls. (C) IL-1 and MyD88-centric gene network in CY17-infected mice compared to uninfected controls (orange = activated function, blue = inhibited function). (D) Top-regulated biofunctions defined by Ingenuity Pathway Analysis (IPA) in CY17-infected untreated mice, compared to cefotaxime- or IL-1RA-treated mice (cut-off Z-score > ±1.5, p-value < 0.05). (E) Top-regulated IL-1β and inflammasome-regulated genes in CY17-infected mice. Activation of IL-1β and inflammasome-related genes in CY17-infected untreated mice and inhibition in IL-1RA- and cefotaxime-treated mice.
Figure 5.
IL-1RA treatment inhibits the expression of pro-inflammatory genes in infected Asc−/− mice. Whole bladder mRNA was subjected to transcriptomic analysis. (A) Gene expression profiles were compared between CY17-infected, untreated controls, and IL-1RA- or cefotaxime-treated mice (n = 2 mice per treatment group, n = 1 for uninfected control). Heat map of regulated genes in the treatment groups and CY17-infected controls compared to uninfected controls (cut-off FC ≥ 2, p < 0.05). Biofunctions of genes specific to CY17-infected controls included neutrophil migration, acute phase responses, inflammation, and antimicrobial responses. These responses were not activated in cefotaxime- or IL-1RA-treated mice (red = activated, blue = inhibited, black = not significantly regulated, compared to uninfected controls). (B) Top-regulated genes in CY17-infected mice treated with cefotaxime or IL-1RA compared to untreated controls. (C) IL-1 and MyD88-centric gene network in CY17-infected mice compared to uninfected controls (orange = activated function, blue = inhibited function). (D) Top-regulated biofunctions defined by Ingenuity Pathway Analysis (IPA) in CY17-infected untreated mice, compared to cefotaxime- or IL-1RA-treated mice (cut-off Z-score > ±1.5, p-value < 0.05). (E) Top-regulated IL-1β and inflammasome-regulated genes in CY17-infected mice. Activation of IL-1β and inflammasome-related genes in CY17-infected untreated mice and inhibition in IL-1RA- and cefotaxime-treated mice.
![Pathogens 13 00042 g005]()
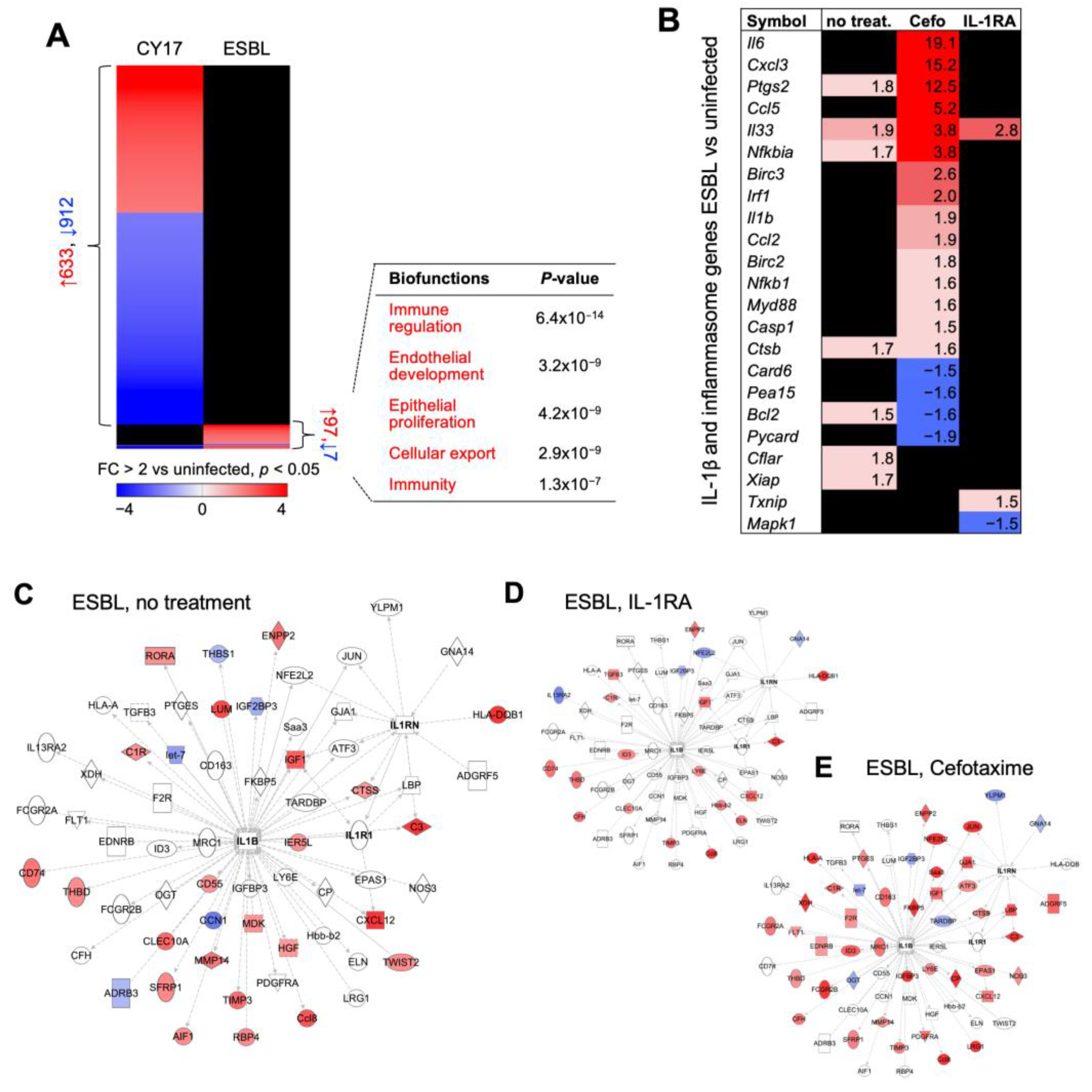
Figure 6.
IL-1RA treatment inhibits and cefotaxime treatment activates the expression of pro-inflammatory genes in infected Asc−/− mice. (A) Gene expression profiles were compared between CY17-infected and ESBL4-infected mice and uninfected controls (n = 2 mice per treatment group, n = 1 for uninfected control, cut-off FC ≥ 2, p < 0.05). A subset of ESBL-specific genes was identified and biofunctions were related to immune regulation, endothelial development, epithelial proliferation, and cellular export. (B) Heat map of IL-1β and inflammasome-regulated genes in the ESBL-infected treatment groups compared to uninfected controls (n = 2 mice per treatment group, n = 1 for uninfected control). (C–E) Limited effects of ESBL on gene expression, shown by an IL-1β and IL-RA-specific gene network (cut-off FC ≥ 2, p < 0.05). Gene networks from IL-RA or cefotaxime-treated mice are shown.
Figure 6.
IL-1RA treatment inhibits and cefotaxime treatment activates the expression of pro-inflammatory genes in infected Asc−/− mice. (A) Gene expression profiles were compared between CY17-infected and ESBL4-infected mice and uninfected controls (n = 2 mice per treatment group, n = 1 for uninfected control, cut-off FC ≥ 2, p < 0.05). A subset of ESBL-specific genes was identified and biofunctions were related to immune regulation, endothelial development, epithelial proliferation, and cellular export. (B) Heat map of IL-1β and inflammasome-regulated genes in the ESBL-infected treatment groups compared to uninfected controls (n = 2 mice per treatment group, n = 1 for uninfected control). (C–E) Limited effects of ESBL on gene expression, shown by an IL-1β and IL-RA-specific gene network (cut-off FC ≥ 2, p < 0.05). Gene networks from IL-RA or cefotaxime-treated mice are shown.
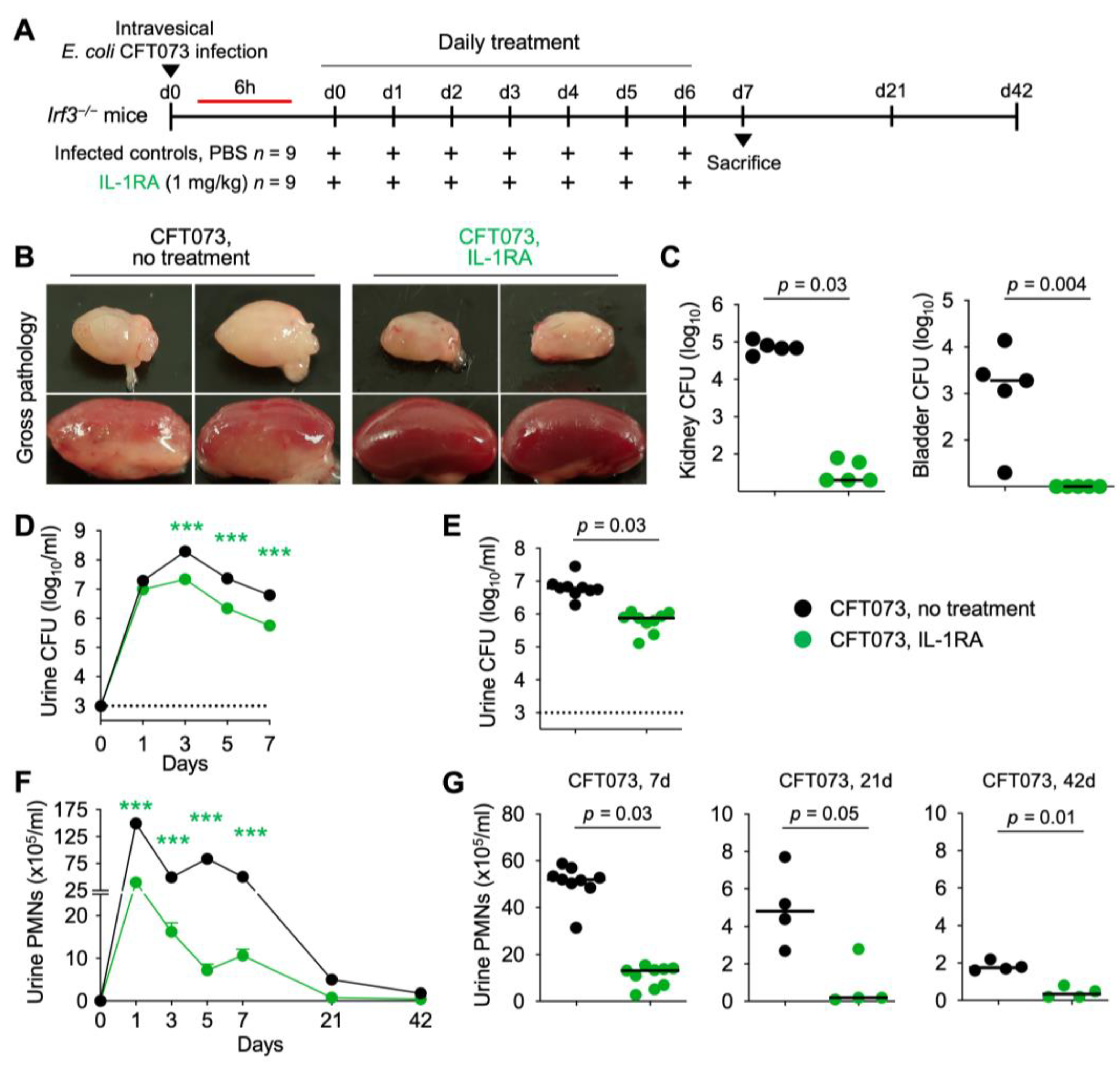
Figure 7.
Efficacy of IL-1RA treatment in Irf3−/− mice with acute pyelonephritis. (A) Schematic of the infection and treatment protocol. Genetically predisposed Irf3−/− mice were infected with the uropathogenic strain E. coli CFT073 (antibiotic sensitive) at 108 CFU. Six hours post-infection, mice were treated I.P. with IL-1RA (anakinra, 1 mg/kg). Treatment was continued daily, until day 7, when mice were sacrificed. Another group was followed for 21- and 42-days post-infection (n = 9 mice per group). (B) Therapeutic efficacy of IL-1RA defined by a loss of gross pathology, hyperemia, and edema. Treatment alleviated acute pyelonephritis in infected mouse kidneys. (C–G) Treatment accelerated bacterial clearance and reduced inflammation, quantified as neutrophil recruitment. (C) Bacterial counts in kidney and bladder tissue (day 7). (D) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, and 7 post-infection. (E) Bacterial counts in urine (day 7). (F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 21, and 42 post-infection. (G) Quantification of neutrophils in urine (days 7, 21, and 42). Representative bladder images (B). Data are presented as individual mice and the horizontal lines represent the median (C,E,G) and mean ± SEM (D,F). Data were analyzed using the Kruskal–Wallis test, *** = p < 0.001.
Figure 7.
Efficacy of IL-1RA treatment in Irf3−/− mice with acute pyelonephritis. (A) Schematic of the infection and treatment protocol. Genetically predisposed Irf3−/− mice were infected with the uropathogenic strain E. coli CFT073 (antibiotic sensitive) at 108 CFU. Six hours post-infection, mice were treated I.P. with IL-1RA (anakinra, 1 mg/kg). Treatment was continued daily, until day 7, when mice were sacrificed. Another group was followed for 21- and 42-days post-infection (n = 9 mice per group). (B) Therapeutic efficacy of IL-1RA defined by a loss of gross pathology, hyperemia, and edema. Treatment alleviated acute pyelonephritis in infected mouse kidneys. (C–G) Treatment accelerated bacterial clearance and reduced inflammation, quantified as neutrophil recruitment. (C) Bacterial counts in kidney and bladder tissue (day 7). (D) Kinetics of infection. Mean log10 CFU/mL urine is shown for days 1, 3, 5, and 7 post-infection. (E) Bacterial counts in urine (day 7). (F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 21, and 42 post-infection. (G) Quantification of neutrophils in urine (days 7, 21, and 42). Representative bladder images (B). Data are presented as individual mice and the horizontal lines represent the median (C,E,G) and mean ± SEM (D,F). Data were analyzed using the Kruskal–Wallis test, *** = p < 0.001.
![Pathogens 13 00042 g007]()
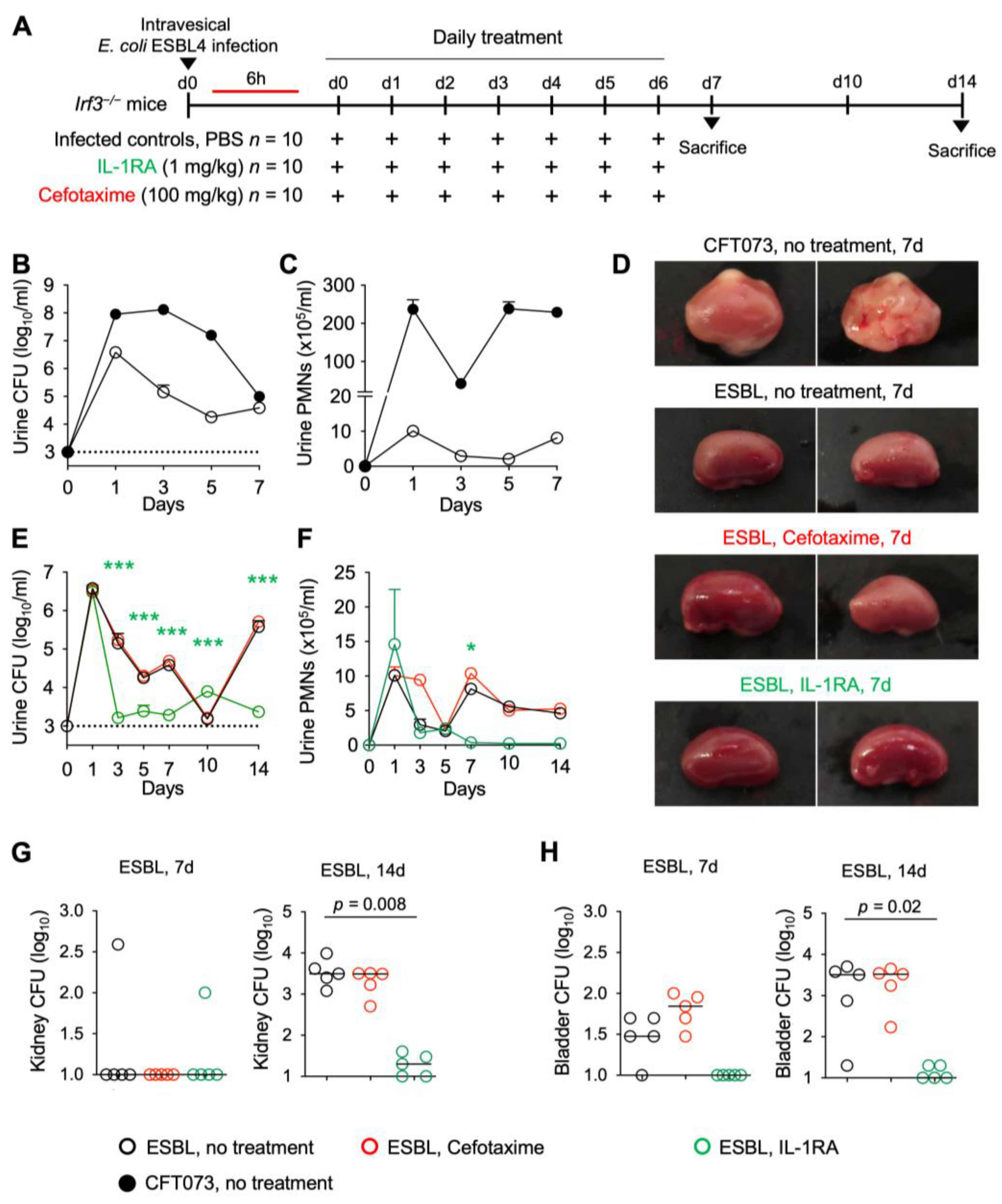
Figure 8.
Efficacy of IL-1RA treatment on kidney infection in
Irf3−/− mice infected with an ESBL isolate.
Irf3−/− mice were infected with the ESBL4
E. coli isolate (10
8 CFU,
n = 10 mice from 1 experiment, 5 mice per time point) and treated according to the protocol shown in
Figure 1A with IL-1RA (anakinra, 1 mg/kg,
n = 10 mice) or cefotaxime (100 mg/kg
n = 10 mice) daily for seven days. PBS was used in the control mice. (
A) Schematic of the treatment protocol. (
B,
C) Comparison of mice infected with the uropathogenic strains CFT073 and ESBL4. (
D) Therapeutic efficacy of IL-1RA defined by a loss of gross kidney pathology, hyperemia, and edema. No treatment effect was seen in cefotaxime-treated mice. (
E) Kinetics of infection. Accelerated bacterial clearance in IL-1RA- but not cefotaxime-treated mice. Mean log
10 CFU/mL urine is shown for days 1, 3, 5, 7, 10, and 14 post-infection. (
F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 10, and 14 post-infection. Neutrophil recruitment was inhibited by IL-1RA but not cefotaxime treatment. (
G) Quantification of bacteria in whole kidney tissue (days 7 and 14). (
H) Quantification of bacteria in whole bladder tissue (days 7 and 14). Data are presented as individual mice, and the horizontal lines represent the median (
G,
H) and mean ± SEM (
B,
C,
E,
F). Data were analyzed using the Kruskal–Wallis test, * =
p < 0.05, *** =
p < 0.001.
Figure 8.
Efficacy of IL-1RA treatment on kidney infection in
Irf3−/− mice infected with an ESBL isolate.
Irf3−/− mice were infected with the ESBL4
E. coli isolate (10
8 CFU,
n = 10 mice from 1 experiment, 5 mice per time point) and treated according to the protocol shown in
Figure 1A with IL-1RA (anakinra, 1 mg/kg,
n = 10 mice) or cefotaxime (100 mg/kg
n = 10 mice) daily for seven days. PBS was used in the control mice. (
A) Schematic of the treatment protocol. (
B,
C) Comparison of mice infected with the uropathogenic strains CFT073 and ESBL4. (
D) Therapeutic efficacy of IL-1RA defined by a loss of gross kidney pathology, hyperemia, and edema. No treatment effect was seen in cefotaxime-treated mice. (
E) Kinetics of infection. Accelerated bacterial clearance in IL-1RA- but not cefotaxime-treated mice. Mean log
10 CFU/mL urine is shown for days 1, 3, 5, 7, 10, and 14 post-infection. (
F) Kinetics of neutrophil recruitment. Mean neutrophil numbers in urine are shown for days 1, 3, 5, 7, 10, and 14 post-infection. Neutrophil recruitment was inhibited by IL-1RA but not cefotaxime treatment. (
G) Quantification of bacteria in whole kidney tissue (days 7 and 14). (
H) Quantification of bacteria in whole bladder tissue (days 7 and 14). Data are presented as individual mice, and the horizontal lines represent the median (
G,
H) and mean ± SEM (
B,
C,
E,
F). Data were analyzed using the Kruskal–Wallis test, * =
p < 0.05, *** =
p < 0.001.
![Pathogens 13 00042 g008]()


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








