
Advances in the Immunology of the Host–Parasite Interactions in African Trypanosomosis, including Single-Cell Transcriptomics
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Life Cycle of Trypanosomes
3. Features of Trypanosomes
4. Human African Trypanosomosis
5. Host Immune Responses to Trypanosomes
5.1. Macrophages and Their Cytokines
5.2. T Cells
5.3. B Cells
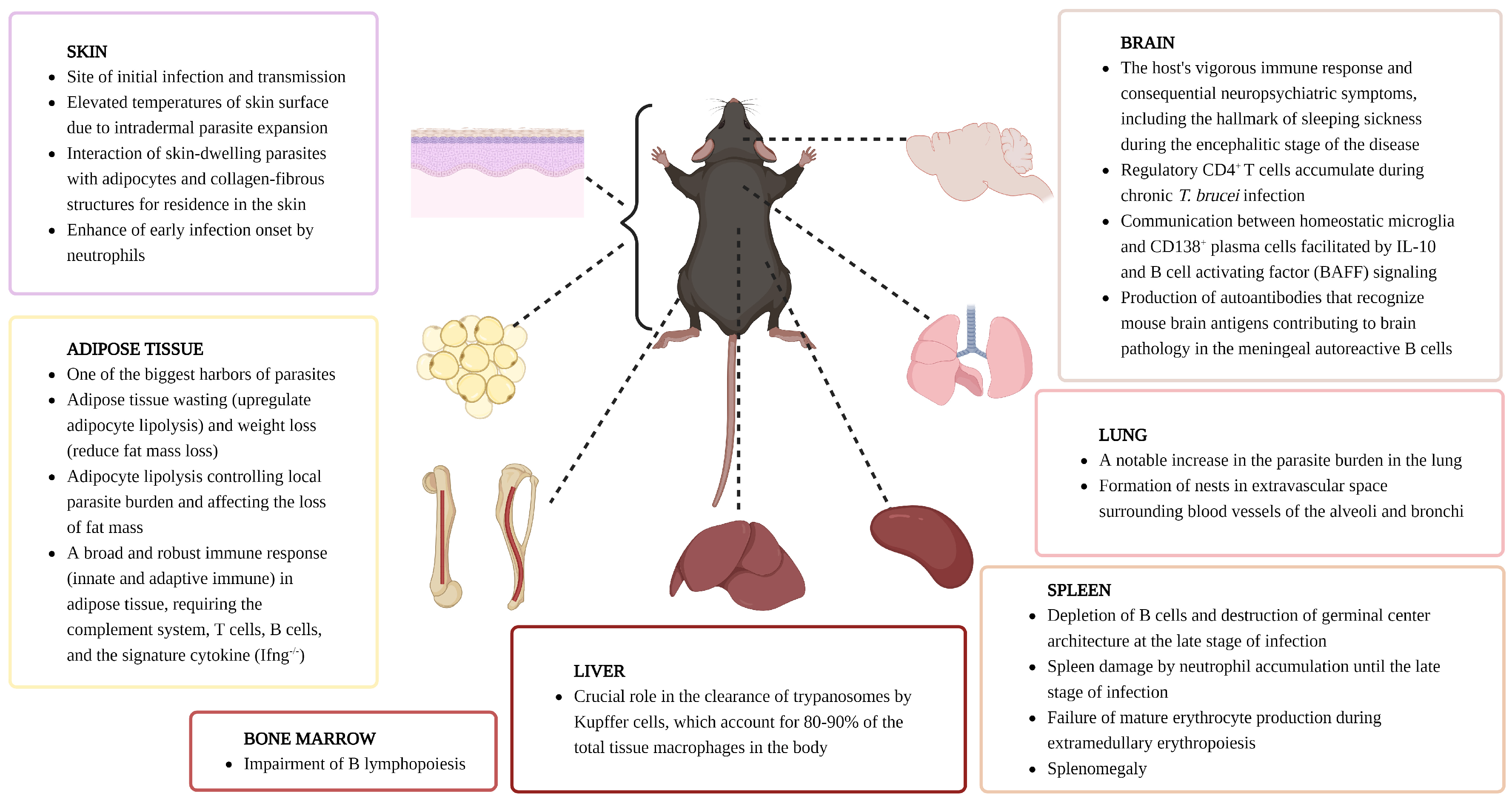
6. Host Immune Responses by Organ
6.1. Skin
6.2. Spleen
6.3. Bone Marrow
6.4. Adipose Tissue
6.5. Brain
6.6. Lung
6.7. Liver
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Human African trypanosomiasis (sleeping sickness): Epidemiological update. Wkly. Epidemiol. Rec. Relev. Épidémiol. Hebd. 2006, 81, 71–80. [Google Scholar]
- Gutierrez, C.; Desquesnes, M.; Touratier, L.; Büscher, P. Trypanosoma evansi: Recent outbreaks in Europe. Vet. Parasitol. 2010, 174, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Desquesnes, M.; Holzmuller, P.; Lai, D.-H.; Dargantes, A.; Lun, Z.-R.; Jittaplapong, S. Trypanosoma evansi and surra: A review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. BioMed Res. Int. 2013, 2013, 194176. [Google Scholar] [CrossRef] [PubMed]
- Robays, J.; Ebeja Kadima, A.; Lutumba, P.; Miaka mia Bilenge, C.; Kande Betu Ku Mesu, V.; De Deken, R.; Makabuza, J.; Deguerry, M.; Van Der Stuyft, P.; Boelaert, M. Human African trypanosomiasis amongst urban residents in Kinshasa: A case-control study. Trop. Med. Int. Health 2004, 9, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Vincendeau, P.; Bouteille, B. Immunology and immunopathology of African trypanosomiasis. An. Acad. Bras. Ciências 2006, 78, 645–665. [Google Scholar] [CrossRef]
- Kristjanson, P.M.; Swallow, B.M.; Rowlands, G.; Kruska, R.; De Leeuw, P. Measuring the costs of African animal trypanosomosis, the potential benefits of control and returns to research. Agric. Syst. 1999, 59, 79–98. [Google Scholar] [CrossRef]
- Coura, J.R.; Borges-Pereira, J. Chagas disease: 100 years after its discovery. A systemic review. Acta Trop. 2010, 115, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Horn, D. Antigenic variation in African trypanosomes. Mol. Biochem. Parasitol. 2014, 195, 123–129. [Google Scholar] [CrossRef]
- Matthews, K.R.; McCulloch, R.; Morrison, L.J. The within-host dynamics of African trypanosome infections. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140288. [Google Scholar] [CrossRef]
- Obishakin, E.; De Trez, C.; Magez, S. Chronic Trypanosoma congolense infections in mice cause a sustained disruption of the B-cell homeostasis in the bone marrow and spleen. Parasite Immunol. 2014, 36, 187–198. [Google Scholar] [CrossRef]
- Paterson, S.; Holmes, W.M.; Rodgers, J. Serial magnetic resonance imaging of splenomegaly in the Trypanosoma brucei infected mouse. PLoS Negl. Trop. Dis. 2022, 16, e0010962. [Google Scholar] [CrossRef] [PubMed]
- Radwanska, M.; Guirnalda, P.; De Trez, C.; Ryffel, B.; Black, S.; Magez, S. Trypanosomiasis-induced B cell apoptosis results in loss of protective anti-parasite antibody responses and abolishment of vaccine-induced memory responses. PLoS Pathog. 2008, 4, e1000078. [Google Scholar] [CrossRef] [PubMed]
- Jaitin, D.A.; Kenigsberg, E.; Keren-Shaul, H.; Elefant, N.; Paul, F.; Zaretsky, I.; Mildner, A.; Cohen, N.; Jung, S.; Tanay, A. Massively parallel single-cell RNA-seq for marker-free decomposition of tissues into cell types. Science 2014, 343, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Radwanska, M.; Vereecke, N.; Deleeuw, V.; Pinto, J.; Magez, S. Salivarian trypanosomosis: A review of parasites involved, their global distribution and their interaction with the innate and adaptive mammalian host immune system. Front. Immunol. 2018, 9, 2253. [Google Scholar] [CrossRef] [PubMed]
- Pays, E.; Vanhollebeke, B.; Vanhamme, L.; Paturiaux-Hanocq, F.; Nolan, D.P.; Pérez-Morga, D. The trypanolytic factor of human serum. Nat. Rev. Microbiol. 2006, 4, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Rojas, F.; Matthews, K.R. Quorum sensing in African trypanosomes. Curr. Opin. Microbiol. 2019, 52, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Briggs, E.M.; Marques, C.A.; Oldrieve, G.R.; Hu, J.; Otto, T.D.; Matthews, K.R. Profiling the bloodstream form and procyclic form Trypanosoma brucei cell cycle using single-cell transcriptomics. eLife 2023, 12, e86325. [Google Scholar] [CrossRef] [PubMed]
- Van Den Abbeele, J.; Claes, Y.; Van Bockstaele, D.; Le Ray, D.; Coosemans, M. Trypanosoma brucei spp. development in the tsetse fly: Characterization of the post-mesocyclic stages in the foregut and proboscis. Parasitology 1999, 118, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Vigneron, A.; O’Neill, M.B.; Weiss, B.L.; Savage, A.F.; Campbell, O.C.; Kamhawi, S.; Valenzuela, J.G.; Aksoy, S. Single-cell RNA sequencing of Trypanosoma brucei from tsetse salivary glands unveils metacyclogenesis and identifies potential transmission blocking antigens. Proc. Natl. Acad. Sci. USA 2020, 117, 2613–2621. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, S.; Foulon, S.; Crouzols, A.; Menafra, R.; Rotureau, B.; Griffiths, A.D.; Bastin, P. The establishment of variant surface glycoprotein monoallelic expression revealed by single-cell RNA-seq of Trypanosoma brucei in the tsetse fly salivary glands. PLoS Pathog. 2021, 17, e1009904. [Google Scholar] [CrossRef]
- Howick, V.M.; Peacock, L.; Kay, C.; Collett, C.; Gibson, W.; Lawniczak, M.K. Single-cell transcriptomics reveals expression profiles of Trypanosoma brucei sexual stages. PLoS Pathog. 2022, 18, e1010346. [Google Scholar] [CrossRef]
- Gibson, W. Liaisons dangereuses: Sexual recombination among pathogenic trypanosomes. Res. Microbiol. 2015, 166, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Gibson, W.; Peacock, L. Fluorescent proteins reveal what trypanosomes get up to inside the tsetse fly. Parasites Vectors 2019, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Magez, S.; Pinto Torres, J.E.; Oh, S.; Radwanska, M. Salivarian trypanosomes have adopted intricate host-pathogen interaction mechanisms that ensure survival in plain sight of the adaptive immune system. Pathogens 2021, 10, 679. [Google Scholar] [CrossRef] [PubMed]
- Pays, E.; Radwanska, M.; Magez, S. The pathogenesis of african trypanosomiasis. Annu. Rev. Pathol. Mech. Dis. 2023, 18, 19–45. [Google Scholar] [CrossRef] [PubMed]
- Trindade, S.; Rijo-Ferreira, F.; Carvalho, T.; Pinto-Neves, D.; Guegan, F.; Aresta-Branco, F.; Bento, F.; Young, S.A.; Pinto, A.; Van Den Abbeele, J.; et al. Trypanosoma brucei Parasites Occupy and Functionally Adapt to the Adipose Tissue in Mice. Cell Host Microbe 2016, 19, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Reuter, C.; Hauf, L.; Imdahl, F.; Sen, R.; Vafadarnejad, E.; Fey, P.; Finger, T.; Jones, N.G.; Walles, H.; Barquist, L. Vector-borne Trypanosoma brucei parasites develop in artificial human skin and persist as skin tissue forms. Nat. Commun. 2023, 14, 7660. [Google Scholar] [CrossRef] [PubMed]
- Dwinger, R.; Rudin, W.; Moloo, S.; Murray, M. Development of Trypanosoma congolense, T vivax and T brucei in the skin reaction induced in goats by infected Glossina morsitans centralis: A light and electron microscopical study. Res. Vet. Sci. 1988, 44, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Magez, S.; Truyens, C.; Merimi, M.; Radwanska, M.; Stijlemans, B.; Brouckaert, P.; Brombacher, F.; Pays, E.; Baetselier, P.D. P75 Tumor Necrosis Factor–Receptor Shedding Occurs as a Protective Host Response during African Trypanosomiasis. J. Infect. Dis. 2004, 189, 527–539. [Google Scholar] [CrossRef]
- Machado, H.; Hofer, P.; Zechner, R.; Smith, T.K.; Figueiredo, L.M. Adipocyte lipolysis protects mice against Trypanosoma brucei infection. Nat. Microbiol. 2023, 8, 2020–2032. [Google Scholar] [CrossRef]
- Paris, Z.; Hashimi, H.; Lun, S.; Alfonzo, J.D.; Lukeš, J. Futile import of tRNAs and proteins into the mitochondrion of Trypanosoma brucei evansi. Mol. Biochem. Parasitol. 2011, 176, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Borst, P.; Fase-Fowler, F.; Gibson, W.C. Kinetoplast DNA of Trypanosoma evansi. Mol. Biochem. Parasitol. 1987, 23, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Moloo, S.; Sabwa, C.; Kabata, J. Vector competence of Glossina pallidipes and G. morsitans centralis for Trypanosoma vivax, T. congolense and T. b. brucei. Acta Trop. 1992, 51, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Blum, M.L.; Down, J.A.; Gurnett, A.M.; Carrington, M.; Turner, M.J.; Wiley, D.C. A structural motif in the variant surface glycoproteins of Trypanosoma brucei. Nature 1993, 362, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Frevert, U.; Reinwald, E. Trypanosoma congolense bloodstream forms evade complement lysis in vitro by shedding of immune complexes. Eur. J. Cell Biol. 1990, 52, 264–269. [Google Scholar] [PubMed]
- Engstler, M.; Pfohl, T.; Herminghaus, S.; Boshart, M.; Wiegertjes, G.; Heddergott, N.; Overath, P. Hydrodynamic flow-mediated protein sorting on the cell surface of trypanosomes. Cell 2007, 131, 505–515. [Google Scholar] [CrossRef] [PubMed]
- La Greca, F.; Haynes, C.; Stijlemans, B.; De Trez, C.; Magez, S. Antibody-mediated control of Trypanosoma vivax infection fails in the absence of tumour necrosis factor. Parasite Immunol. 2014, 36, 271–276. [Google Scholar] [CrossRef]
- Marcello, L.; Barry, J.D. Analysis of the VSG gene silent archive in Trypanosoma brucei reveals that mosaic gene expression is prominent in antigenic variation and is favored by archive substructure. Genome Res. 2007, 17, 1344–1352. [Google Scholar] [CrossRef]
- Hall, J.P.; Wang, H.; Barry, J.D. Mosaic VSGs and the scale of Trypanosoma brucei antigenic variation. PLoS Pathog. 2013, 9, e1003502. [Google Scholar] [CrossRef]
- Vassella, E.; Reuner, B.; Yutzy, B.; Boshart, M. Differentiation of African trypanosomes is controlled by a density sensing mechanism which signals cell cycle arrest via the cAMP pathway. J. Cell Sci. 1997, 110, 2661–2671. [Google Scholar] [CrossRef]
- Rojas, F.; Silvester, E.; Young, J.; Milne, R.; Tettey, M.; Houston, D.R.; Walkinshaw, M.D.; Perez-Pi, I.; Auer, M.; Denton, H. Oligopeptide signaling through TbGPR89 drives trypanosome quorum sensing. Cell 2019, 176, 306–317.e316. [Google Scholar] [CrossRef] [PubMed]
- MacGregor, P.; Szöőr, B.; Savill, N.J.; Matthews, K.R. Trypanosomal immune evasion, chronicity and transmission: An elegant balancing act. Nat. Rev. Microbiol. 2012, 10, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Briggs, E.M.; Rojas, F.; McCulloch, R.; Matthews, K.R.; Otto, T.D. Single-cell transcriptomic analysis of bloodstream Trypanosoma brucei reconstructs cell cycle progression and developmental quorum sensing. Nat. Commun. 2021, 12, 5268. [Google Scholar] [CrossRef]
- Franco, J.R.; Cecchi, G.; Priotto, G.; Paone, M.; Diarra, A.; Grout, L.; Simarro, P.P.; Zhao, W.; Argaw, D. Monitoring the elimination of human African trypanosomiasis at continental and country level: Update to 2018. PLoS Negl. Trop. Dis. 2020, 14, e0008261. [Google Scholar] [CrossRef] [PubMed]
- Masocha, W.; Kristensson, K. Human African trypanosomiasis: How do the parasites enter and cause dysfunctions of the nervous system in murine models? Brain Res. Bull. 2019, 145, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Poelvoorde, P.; Vanhamme, L.; Van Den Abbeele, J.; Switzer, W.M.; Pays, E. Distribution of apolipoprotein LI and trypanosome lytic activity among primate sera. Mol. Biochem. Parasitol. 2004, 134, 155–157. [Google Scholar] [CrossRef] [PubMed]
- Raper, J.; Friedman, D.J. Molecular one-upmanship. Nature 2013, 501, 322–323. [Google Scholar] [CrossRef]
- Bullard, W.; Kieft, R.; Capewell, P.; Veitch, N.J.; Macleod, A.; Hajduk, S. Haptoglobin-hemoglobin receptor independent killing of African trypanosomes by human serum and trypanosome lytic factors. Virulence 2012, 3, 72–76. [Google Scholar] [CrossRef]
- Lecordier, L.; Uzureau, P.; Tebabi, P.; Pérez-Morga, D.; Nolan, D.; Schumann Burkard, G.; Roditi, I.; Pays, E. Identification of Trypanosoma brucei components involved in trypanolysis by normal human serum. Mol. Microbiol. 2014, 94, 625–636. [Google Scholar] [CrossRef]
- Verdi, J.; Zipkin, R.; Hillman, E.; Gertsch, R.A.; Pangburn, S.J.; Thomson, R.; Papavasiliou, N.; Sternberg, J.; Raper, J. Inducible germline IgMs bridge trypanosome lytic factor assembly and parasite recognition. Cell Host Microbe 2020, 28, 79–88.e74. [Google Scholar] [CrossRef]
- Radwanska, M.; Claes, F.; Magez, S.; Magnus, E.; Perez-Morga, D.; Pays, E.; Büscher, P. Novel primer sequences for polymerase chain reaction-based detection of Trypanosoma brucei gambiense. Am. J. Trop. Med. Hyg. 2002, 67, 289–295. [Google Scholar] [CrossRef]
- Jamonneau, V.; Truc, P.; Grébaut, P.; Herder, S.; Ravel, S.; Solano, P.; De Meeus, T. Trypanosoma brucei gambiense Group 2: The unusual suspect. Trends Parasitol. 2019, 35, 983–995. [Google Scholar] [CrossRef] [PubMed]
- Capewell, P.; Cooper, A.; Clucas, C.; Weir, W.; Macleod, A. A co-evolutionary arms race: Trypanosomes shaping the human genome, humans shaping the trypanosome genome. Parasitology 2015, 142, S108–S119. [Google Scholar] [CrossRef] [PubMed]
- DeJesus, E.; Kieft, R.; Albright, B.; Stephens, N.; Hajduk, S. A single amino acid substitution in the group 1 Trypanosoma brucei gambiense haptoglobin-hemoglobin receptor abolishes TLF-1 binding. PLoS Pathog. 2013, 9, e1003317. [Google Scholar] [CrossRef] [PubMed]
- Uzureau, P.; Uzureau, S.; Lecordier, L.; Fontaine, F.; Tebabi, P.; Homblé, F.; Grélard, A.; Zhendre, V.; Nolan, D.P.; Lins, L. Mechanism of Trypanosoma brucei gambiense resistance to human serum. Nature 2013, 501, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Mulindwa, J.; Matovu, E.; Enyaru, J.; Clayton, C. Blood signatures for second stage human African trypanosomiasis: A transcriptomic approach. BMC Med Genom. 2020, 13, 14. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Varin, A.; Chen, Y.; Liu, B.; Tryggvason, K.; Gordon, S. SR-A/MARCO–mediated ligand delivery enhances intracellular TLR and NLR function, but ligand scavenging from cell surface limits TLR4 response to pathogens. Blood J. Am. Soc. Hematol. 2011, 117, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Sellar, G.C.; Blake, D.J.; Reid, K. Characterization and organization of the genes encoding the A-, B-and C-chains of human complement subcomponent C1q. The complete derived amino acid sequence of human C1q. Biochem. J. 1991, 274, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.T.; Breden, F. The immunoglobulin heavy chain locus: Genetic variation, missing data, and implications for human disease. Genes Immun. 2012, 13, 363–373. [Google Scholar] [CrossRef]
- Mühleisen, T.W.; Leber, M.; Schulze, T.G.; Strohmaier, J.; Degenhardt, F.; Treutlein, J.; Mattheisen, M.; Forstner, A.J.; Schumacher, J.; Breuer, R. Genome-wide association study reveals two new risk loci for bipolar disorder. Nat. Commun. 2014, 5, 3339. [Google Scholar] [CrossRef]
- Suárez-Rama, J.J.; Arrojo, M.; Sobrino, B.; Amigo, J.; Brenlla, J.; Agra, S.; Paz, E.; Brión, M.; Carracedo, Á.; Páramo, M. Resequencing and association analysis of coding regions at twenty candidate genes suggest a role for rare risk variation at AKAP9 and protective variation at NRXN1 in schizophrenia susceptibility. J. Psychiatr. Res. 2015, 66, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Gottesmann, C. GABA mechanisms and sleep. Neuroscience 2002, 111, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Liu, Z.; Zhou, X.; Yuan, G. Association of sleep duration with apolipoproteins and the apolipoprotein B/A1 ratio: The China health and nutrition survey. Nutr. Metab. 2018, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Caljon, G.; Van Reet, N.; De Trez, C.; Vermeersch, M.; Perez-Morga, D.; Van Den Abbeele, J. The dermis as a delivery site of Trypanosoma brucei for tsetse flies. PLoS Pathog. 2016, 12, e1005744. [Google Scholar] [CrossRef] [PubMed]
- Capewell, P.; Cren-Travaillé, C.; Marchesi, F.; Johnston, P.; Clucas, C.; Benson, R.A.; Gorman, T.-A.; Calvo-Alvarez, E.; Crouzols, A.; Jouvion, G. The skin is a significant but overlooked anatomical reservoir for vector-borne African trypanosomes. Elife 2016, 5, e17716. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.G. Clinical features, diagnosis, and treatment of human African trypanosomiasis (sleeping sickness). Lancet Neurol. 2013, 12, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.F.; Sinton, M.C.; Chandrasegaran, P.; Lestari, A.N.; Heslop, R.; Cheaib, B.; Ogunsola, J.; Ngoyi, D.M.; Kuispond Swar, N.-R.; Cooper, A. γδ T cells control murine skin inflammation and subcutaneous adipose wasting during chronic Trypanosoma brucei infection. Nat. Commun. 2023, 14, 5279. [Google Scholar] [CrossRef]
- Caljon, G.; Mabille, D.; Stijlemans, B.; De Trez, C.; Mazzone, M.; Tacchini-Cottier, F.; Malissen, M.; Van Ginderachter, J.A.; Magez, S.; De Baetselier, P. Neutrophils enhance early Trypanosoma brucei infection onset. Sci. Rep. 2018, 8, 11203. [Google Scholar] [CrossRef]
- Deleeuw, V.; Phạm, H.T.T.; De Poorter, I.; Janssens, I.; De Trez, C.; Radwanska, M.; Magez, S. Trypanosoma brucei brucei causes a rapid and persistent influx of neutrophils in the spleen of infected mice. Parasite Immunol. 2019, 41, e12664. [Google Scholar] [CrossRef]
- Anosa, V.O.; Kaneko, J.J. Pathogenesis of Trypanosoma brucei infection in deer mice (Peromyscus maniculatus). Ultrastructural pathology of the spleen, liver, heart, and kidney. Vet. Pathol. 1984, 21, 229–237. [Google Scholar] [CrossRef]
- Crilly, N.P.; Mugnier, M.R. Thinking outside the blood: Perspectives on tissue-resident Trypanosoma brucei. PLoS Pathog. 2021, 17, e1009866. [Google Scholar] [CrossRef]
- Kaushik, R.S.; Uzonna, J.E.; Zhang, Y.; Gordon, J.R.; Tabel, H. Innate resistance to experimental African trypanosomiasis: Differences in cytokine (TNF-α, IL-6, IL-10 and IL-12) production by bone marrow-derived macrophages from resistant and susceptible mice. Cytokine 2000, 12, 1024–1034. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M.; Ericsson, A.C. Function of macrophages in disease: Current understanding on molecular mechanisms. Front. Immunol. 2021, 12, 620510. [Google Scholar] [CrossRef]
- Guerriero, J.L. Macrophages: Their untold story in T cell activation and function. Int. Rev. Cell Mol. Biol. 2019, 342, 73–93. [Google Scholar] [PubMed]
- Arango Duque, G.; Descoteaux, A. Macrophage cytokines: Involvement in immunity and infectious diseases. Front. Immunol. 2014, 5, 491. [Google Scholar] [CrossRef] [PubMed]
- Shoda, L.K.; Kegerreis, K.A.; Suarez, C.E.; Roditi, I.; Corral, R.S.; Bertot, G.M.; Norimine, J.; Brown, W.C. DNA from protozoan parasites Babesia bovis, Trypanosoma cruzi, and T. brucei is mitogenic for B lymphocytes and stimulates macrophage expression of interleukin-12, tumor necrosis factor alpha, and nitric oxide. Infect. Immun. 2001, 69, 2162–2171. [Google Scholar] [CrossRef] [PubMed]
- Magez, S.; Stijlemans, B.; Baral, T.; De Baetselier, P. VSG-GPI anchors of African trypanosomes: Their role in macrophage activation and induction of infection-associated immunopathology. Microbes Infect. 2002, 4, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Wei, G.; Pan, W.; Tabel, H. Trypanosoma congolense infections: Antibody-mediated phagocytosis by Kupffer cells. J. Leukoc. Biol. 2004, 76, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, R.S.; Uzonna, J.E.; Gordon, J.R.; Tabel, H. Innate resistance to Trypanosoma congolense infections: Differential production of nitric oxide by macrophages from susceptible BALB/c and resistant C57Bl/6 mice. Exp. Parasitol. 1999, 92, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Okomo-Assoumou, M.C.; Daulouede, S.; Lemesre, J.-L.; N’Zila-Mouanda, A.; Vincendeau, P. Correlation of high serum levels of tumor necrosis factor-α with disease severity in human African trypanosomiasis. Am. J. Trop. Med. Hyg. 1995, 53, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.; Gow, J.; Kennedy, P.; Jennings, F.; Murray, M. Immunopathology of experimental African sleeping sickness: Detection of cytokine mRNA in the brains of Trypanosoma brucei brucei-infected mice. Infect. Immun. 1991, 59, 4636–4640. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.F.; Chandrasegaran, P.; Sinton, M.C.; Briggs, E.M.; Otto, T.D.; Heslop, R.; Bentley-Abbot, C.; Loney, C.; de Lecea, L.; Mabbott, N.A.; et al. Single cell and spatial transcriptomic analyses reveal microglia-plasma cell crosstalk in the brain during Trypanosoma brucei infection. Nat. Commun. 2022, 13, 5752. [Google Scholar] [CrossRef] [PubMed]
- Montague, P.; Bradley, B.; Rodgers, J.; Kennedy, P.G.E. Microarray profiling predicts early neurological and immune phenotypic traits in advance of CNS disease during disease progression in Trypanosoma. b. brucei infected CD1 mouse brains. PLoS Negl. Trop. Dis. 2021, 15, e0009892. [Google Scholar] [CrossRef] [PubMed]
- Baral, T.N. Immunobiology of African trypanosomes: Need of alternative interventions. BioMed Res. Int. 2010, 2010, 389153. [Google Scholar] [CrossRef] [PubMed]
- Onyilagha, C.; Singh, R.; Gounni, A.S.; Uzonna, J.E. Thymic stromal lymphopoietin is critical for regulation of proinflammatory cytokine response and resistance to experimental Trypanosoma congolense infection. Front. Immunol. 2017, 8, 803. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Inoue, M.; Kuriki, K.; Yamaguchi, H.; Nagasawa, H.; Mikami, T.; Fujisaki, K.; Suzuki, N.; Hirumi, H. Interleukin 4 is a crucial cytokine in controlling Trypanosoma brucei gambiense infection in mice. Vet. Parasitol. 1999, 86, 173–184. [Google Scholar] [CrossRef]
- Shi, M.; Wei, G.; Tabel, H. Trypanosoma congolense infections: MHC class II-restricted immune responses mediate either protection or disease, depending on IL-10 function. Parasite Immunol. 2007, 29, 107–111. [Google Scholar] [CrossRef]
- Namangala, B.; Noël, W.; De Baetselier, P.; Brys, L.; Beschin, A. Relative contribution of interferon-γ and interleukin-10 to resistance to murine African trypanosomosis. J. Infect. Dis. 2001, 183, 1794–1800. [Google Scholar] [CrossRef]
- Tellier, J.; Nutt, S.L. The unique features of follicular T cell subsets. Cell. Mol. Life Sci. 2013, 70, 4771–4784. [Google Scholar] [CrossRef]
- Shi, M.; Wei, G.; Pan, W.; Tabel, H. Experimental African trypanosomiasis: A subset of pathogenic, IFN-γ-producing, MHC class II-restricted CD4+ T cells mediates early mortality in highly susceptible mice. J. Immunol. 2006, 176, 1724–1732. [Google Scholar] [CrossRef]
- Vaidya, T.; Bakhiet, M.; Hill, K.L.; Olsson, T.; Kristensson, K.; Donelson, J.E. The gene for a T lymphocyte triggering factor from African trypanosomes. J. Exp. Med. 1997, 186, 433–438. [Google Scholar] [CrossRef]
- Hamadien; Lycke; Bakhiet. Induction of the trypanosome lymphocyte-triggering factor (TLTF) and neutralizing antibodies to the TLTF in experimental African trypanosomiasis. Immunology 1999, 96, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Cnops, J.; De Trez, C.; Stijlemans, B.; Keirsse, J.; Kauffmann, F.; Barkhuizen, M.; Keeton, R.; Boon, L.; Brombacher, F.; Magez, S. NK-, NKT-and CD8-derived IFNγ drives myeloid cell activation and erythrophagocytosis, resulting in trypanosomosis-associated acute anemia. PLoS Pathog. 2015, 11, e1004964. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, G.; Shi, M. Interferon gamma in African trypanosome infections: Friends or foes? Front. Immunol. 2017, 8, 1105. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Sun, D.; Wu, H.; Zhang, M.; Huan, H.; Xu, J.; Zhang, X.; Zhou, H.; Shi, M. Distinct contributions of CD4+ and CD8+ T cells to pathogenesis of Trypanosoma brucei infection in the context of gamma interferon and interleukin-10. Infect. Immun. 2015, 83, 2785–2795. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Magez, S.; Radwanska, M. From helping to regulating–A transcriptomic profile of Ifng+ Il10+ Il21+ Cd4+ Th1 cells indicates their role in regulating inflammation during experimental trypanosomosis. Front. Trop. Dis. 2023, 4, 1127022. [Google Scholar] [CrossRef]
- Onyilagha, C.; Jia, P.; Jayachandran, N.; Hou, S.; Okwor, I.; Kuriakose, S.; Marshall, A.; Uzonna, J.E. The B cell adaptor molecule Bam32 is critically important for optimal antibody response and resistance to Trypanosoma congolense infection in mice. PLoS Negl. Trop. Dis. 2015, 9, e0003716. [Google Scholar] [CrossRef]
- Borst, P. Antigenic variation and allelic exclusion. Cell 2002, 109, 5–8. [Google Scholar] [CrossRef]
- Morrison, W.I.; Murray, M. The role of humoral immune responses in determining susceptibility of A/J and C57BL/6 mice to infection with Trypanosoma congolense. Parasite Immunol. 1985, 7, 63–79. [Google Scholar] [CrossRef]
- Williams, D.; Taylor, K.; Newson, J.; Gichuki, B.; Naessens, J. The role of anti-variable surface glycoprotein antibody responses in bovine trypanotolerance. Parasite Immunol. 1996, 18, 209–218. [Google Scholar] [CrossRef]
- Pan, W.; Ogunremi, O.; Wei, G.; Shi, M.; Tabel, H. CR3 (CD11b/CD18) is the major macrophage receptor for IgM antibody-mediated phagocytosis of African trypanosomes: Diverse effect on subsequent synthesis of tumor necrosis factor α and nitric oxide. Microbes Infect. 2006, 8, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Magez, S.; Pinto Torres, J.E.; Obishakin, E.; Radwanska, M. Infections with extracellular trypanosomes require control by efficient innate immune mechanisms and can result in the destruction of the mammalian humoral immune system. Front. Immunol. 2020, 11, 382. [Google Scholar] [CrossRef] [PubMed]
- Gatto, D.; Brink, R. The germinal center reaction. J. Allergy Clin. Immunol. 2010, 126, 898–907. [Google Scholar] [CrossRef]
- Takahashi, Y.; Miyamoto, H.; Fukuma, T.; Nishiyama, T.; Araki, T.; Shinka, S. In vivo interaction between Trypanosoma gambiense and leucocytes in mice. Zentralblatt Bakteriol. Mikrobiol. Hyg. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1987, 264, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Magez, S.; Schwegmann, A.; Atkinson, R.; Claes, F.; Drennan, M.; De Baetselier, P.; Brombacher, F. The role of B-cells and IgM antibodies in parasitemia, anemia, and VSG switching in trypanosoma brucei–infected mice. PLoS Pathog. 2008, 4, e1000122. [Google Scholar] [CrossRef] [PubMed]
- Baral, T.N.; De Baetselier, P.; Brombacher, F.; Magez, S. Control of Trypanosoma evansi infection is IgM mediated and does not require a type I inflammatory response. J. Infect. Dis. 2007, 195, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Guevarra, R.B.; Magez, S.; Radwanska, M. Single-cell transcriptome profiling and the use of AID deficient mice reveal that B cell activation combined with antibody class switch recombination and somatic hypermutation do not benefit the control of experimental trypanosomosis. PLoS Pathog. 2021, 17, e1010026. [Google Scholar] [CrossRef]
- Murray, M.; Murray, P.; Jennings, F.; Fisher, E.; Urquhart, G. The pathology of Trypanosoma brucei infection in the rat. Res. Vet. Sci. 1974, 16, 77–84. [Google Scholar] [CrossRef]
- Hudson, K.; Byner, C.; Freeman, J.; Terry, R. Immunodepression, high IgM levels and evasion of the immune response in murine trypanosomiasis. Nature 1976, 264, 256–258. [Google Scholar] [CrossRef]
- Luckins, A.; Mehlitz, D. Observations on serum immunoglobulin levels in cattle infected with Trypanosoma brucei, T. vivax and T. congolense. Ann. Trop. Med. Parasitol. 1976, 70, 479–480. [Google Scholar] [CrossRef]
- Drennan, M.B.; Stijlemans, B.t.; Van Den Abbeele, J.; Quesniaux, V.J.; Barkhuizen, M.; Brombacher, F.; De Baetselier, P.; Ryffel, B.; Magez, S. The induction of a type 1 immune response following a Trypanosoma brucei infection is MyD88 dependent. J. Immunol. 2005, 175, 2501–2509. [Google Scholar] [CrossRef] [PubMed]
- Kobayakawa, T.; Louis, J.; Izui, S.; Lambert, P.-H. Autoimmune response to DNA, red blood cells, and thymocyte antigens in association with polyclonal antibody synthesis during experimental African trypanosomiasis. J. Immunol. 1979, 122, 296–301. [Google Scholar] [CrossRef]
- Onyilagha, C.; Uzonna, J.E. Host immune responses and immune evasion strategies in African trypanosomiasis. Front. Immunol. 2019, 10, 2738. [Google Scholar] [CrossRef] [PubMed]
- Frid, K.; Einstein, O.; Friedman-Levi, Y.; Binyamin, O.; Ben-Hur, T.; Gabizon, R. Aggregation of MBP in chronic demyelination. Ann. Clin. Transl. Neurol. 2015, 2, 711–721. [Google Scholar] [CrossRef] [PubMed]
- D’Aversa, T.G.; Eugenin, E.A.; Lopez, L.; Berman, J.W. Myelin basic protein induces inflammatory mediators from primary human endothelial cells and blood–brain barrier disruption: Implications for the pathogenesis of multiple sclerosis. Neuropathol. Appl. Neurobiol. 2013, 39, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Quintana, J.F.; Sinton, M.C.; Chandrasegaran, P.; Kumar Dubey, L.; Ogunsola, J.; Al Samman, M.; Haley, M.; McConnell, G.; Kuispond Swar, N.-R.; Ngoyi, D.M. The murine meninges acquire lymphoid tissue properties and harbour autoreactive B cells during chronic Trypanosoma brucei infection. PLoS Biol. 2023, 21, e3002389. [Google Scholar] [CrossRef] [PubMed]
- Bockstal, V.; Guirnalda, P.; Caljon, G.; Goenka, R.; Telfer, J.C.; Frenkel, D.; Radwanska, M.; Magez, S.; Black, S.J. T. brucei infection reduces B lymphopoiesis in bone marrow and truncates compensatory splenic lymphopoiesis through transitional B-cell apoptosis. PLoS Pathog. 2011, 7, e1002089. [Google Scholar] [CrossRef]
- Moon, S.; Janssens, I.; Kim, K.H.; Stijlemans, B.; Magez, S.; Radwanska, M. Detrimental effect of Trypanosoma brucei brucei infection on memory B cells and host ability to recall protective B-cell responses. J. Infect. Dis. 2022, 226, 528–540. [Google Scholar] [CrossRef]
- Pham, H.T.T.; Magez, S.; Choi, B.; Baatar, B.; Jung, J.; Radwanska, M. Neutrophil metalloproteinase driven spleen damage hampers infection control of trypanosomiasis. Nat. Commun. 2023, 14, 5418. [Google Scholar] [CrossRef]
- Alfituri, O.A.; Quintana, J.F.; MacLeod, A.; Garside, P.; Benson, R.A.; Brewer, J.M.; Mabbott, N.A.; Morrison, L.J.; Capewell, P. To the skin and beyond: The immune response to African trypanosomes as they enter and exit the vertebrate host. Front. Immunol. 2020, 11, 1250. [Google Scholar] [CrossRef]
- Theis, J.; Bolton, V. Trypanosoma equiperdum: Movement from the dermis. Exp. Parasitol. 1980, 50, 317–330. [Google Scholar] [CrossRef]
- Sudarshi, D.; Lawrence, S.; Pickrell, W.O.; Eligar, V.; Walters, R.; Quaderi, S.; Walker, A.; Capewell, P.; Clucas, C.; Vincent, A. Human African trypanosomiasis presenting at least 29 years after infection—What can this teach us about the pathogenesis and control of this neglected tropical disease? PLoS Negl. Trop. Dis. 2014, 8, e3349. [Google Scholar] [CrossRef]
- Jamonneau, V.; Ilboudo, H.; Kaboré, J.; Kaba, D.; Koffi, M.; Solano, P.; Garcia, A.; Courtin, D.; Laveissière, C.; Lingue, K. Untreated human infections by Trypanosoma brucei gambiense are not 100% fatal. PLoS Negl. Trop. Dis. 2012, 6, e1691. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Jameson, J.; Ugarte, K.; Chen, N.; Yachi, P.; Fuchs, E.; Boismenu, R.; Havran, W.L. A role for skin γδ T cells in wound repair. Science 2002, 296, 747–749. [Google Scholar] [CrossRef]
- Konieczny, P.; Xing, Y.; Sidhu, I.; Subudhi, I.; Mansfield, K.P.; Hsieh, B.; Biancur, D.E.; Larsen, S.B.; Cammer, M.; Li, D. Interleukin-17 governs hypoxic adaptation of injured epithelium. Science 2022, 377, eabg9302. [Google Scholar] [CrossRef] [PubMed]
- Roark, C.L.; Simonian, P.L.; Fontenot, A.P.; Born, W.K.; O’Brien, R.L. γδ T cells: An important source of IL-17. Curr. Opin. Immunol. 2008, 20, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.R.; Costanzo, A.E.; Jameson, J.M. Dysfunctional γδ T cells contribute to impaired keratinocyte homeostasis in mouse models of obesity. J. Investig. Dermatol. 2011, 131, 2409–2418. [Google Scholar] [CrossRef] [PubMed]
- Fielding, J.W.; Hallissey, M.T.; Tilanus, H.W. The Spleen. In Upper Gastrointestinal Surger; Springer: London, UK, 2005; pp. 59–67. [Google Scholar]
- MacLean, L.M.; Odiit, M.; Chisi, J.E.; Kennedy, P.G.; Sternberg, J.M. Focus–specific clinical profiles in human african trypanosomiasis caused by Trypanosoma brucei rhodesiense. PLoS Negl. Trop. Dis. 2010, 4, e906. [Google Scholar] [CrossRef]
- Bisoffi, Z.; Beltrame, A.; Monteiro, G.; Arzese, A.; Marocco, S.; Rorato, G.; Anselmi, M.; Viale, P. African trypanosomiasis gambiense, Italy. Emerg. Infect. Dis. 2005, 11, 1745. [Google Scholar] [CrossRef]
- Nguyen, H.T.T.; Radwanska, M.; Magez, S. Tipping the balance between erythroid cell differentiation and induction of anemia in response to the inflammatory pathology associated with chronic trypanosome infections. Front. Immunol. 2022, 13, 1051647. [Google Scholar] [CrossRef] [PubMed]
- Blom-Potar, M.C.; Chamond, N.; Cosson, A.; Jouvion, G.; Droin-Bergère, S.; Huerre, M.; Minoprio, P. Trypanosoma vivax infections: Pushing ahead with mouse models for the study of Nagana. II. Immunobiological dysfunctions. PLoS Negl. Trop. Dis. 2010, 4, e793. [Google Scholar] [CrossRef] [PubMed]
- Onyilagha, C.; Okwor, I.; Kuriakose, S.; Singh, R.; Uzonna, J. Low-dose intradermal infection with trypanosoma congolense leads to expansion of regulatory T cells and enhanced susceptibility to reinfection. Infect. Immun. 2014, 82, 1074–1083. [Google Scholar] [CrossRef] [PubMed]
- Guilliams, M.; Oldenhove, G.; Noel, W.; Hérin, M.; Brys, L.; Loi, P.; Flamand, V.; Moser, M.; De Baetselier, P.; Beschin, A. African trypanosomiasis: Naturally occurring regulatory T cells favor trypanotolerance by limiting pathology associated with sustained type 1 inflammation. J. Immunol. 2007, 179, 2748–2757. [Google Scholar] [CrossRef] [PubMed]
- Evrard, M.; Kwok, I.W.; Chong, S.Z.; Teng, K.W.; Becht, E.; Chen, J.; Sieow, J.L.; Penny, H.L.; Ching, G.C.; Devi, S. Developmental analysis of bone marrow neutrophils reveals populations specialized in expansion, trafficking, and effector functions. Immunity 2018, 48, 364–379.e368. [Google Scholar] [CrossRef] [PubMed]
- Grieshaber-Bouyer, R.; Radtke, F.A.; Cunin, P.; Stifano, G.; Levescot, A.; Vijaykumar, B.; Nelson-Maney, N.; Blaustein, R.B.; Monach, P.A.; Nigrovic, P.A. The neutrotime transcriptional signature defines a single continuum of neutrophils across biological compartments. Nat. Commun. 2021, 12, 2856. [Google Scholar] [CrossRef]
- Xie, X.; Shi, Q.; Wu, P.; Zhang, X.; Kambara, H.; Su, J.; Yu, H.; Park, S.-Y.; Guo, R.; Ren, Q. Single-cell transcriptome profiling reveals neutrophil heterogeneity in homeostasis and infection. Nat. Immunol. 2020, 21, 1119–1133. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C.; Ramirez-Acuna, J.M.; Perez-Romero, B.A.; Guerrero-Rodriguez, J.F.; Martinez-Avila, N.; Martinez-Fierro, M.L. The roles of matrix metalloproteinases and their inhibitors in human diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Lin, M.; Jackson, P.; Tester, A.M.; Diaconu, E.; Overall, C.M.; Blalock, J.E.; Pearlman, E. Matrix metalloproteinase-8 facilitates neutrophil migration through the corneal stromal matrix by collagen degradation and production of the chemotactic peptide Pro-Gly-Pro. Am. J. Pathol. 2008, 173, 144–153. [Google Scholar] [CrossRef]
- Charzewski, Ł.; Krzyśko, K.A.; Lesyng, B. Structural characterisation of inhibitory and non-inhibitory MMP-9–TIMP-1 complexes and implications for regulatory mechanisms of MMP-9. Sci. Rep. 2021, 11, 13376. [Google Scholar] [CrossRef]
- Magez, S.; Radwanska, M.; Drennan, M.; Fick, L.; Baral, T.N.; Brombacher, F.; Baetselier, P.D. Interferon-γ and nitric oxide in combination with antibodies are key protective host immune factors during Trypanosoma congolense Tc13 infections. J. Infect. Dis. 2006, 193, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Cox, F. Pathogenesis of animal trypanosomiasis. Nature 1979, 277, 603–604. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; Wagers, A.J.; Manz, M.G.; Prohaska, S.S.; Scherer, D.C.; Beilhack, G.F.; Shizuru, J.A.; Weissman, I.L. Biology of hematopoietic stem cells and progenitors: Implications for clinical application. Annu. Rev. Immunol. 2003, 21, 759–806. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W.; Tedder, T.F. B lymphocytes: How they develop and function. Blood J. Am. Soc. Hematol. 2008, 112, 1570–1580. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Yang, K.; Foster, S.J.; Kondo, M.; Kelsoe, G. Inflammation controls B lymphopoiesis by regulating chemokine CXCL12 expression. J. Exp. Med. 2004, 199, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Camara, O.; Camara, M.; Falzon, L.C.; Ilboudo, H.; Kaboré, J.; Compaoré, C.F.A.; Fèvre, E.M.; Büscher, P.; Bucheton, B.; Lejon, V. Performance of clinical signs and symptoms, rapid and reference laboratory diagnostic tests for diagnosis of human African trypanosomiasis by passive screening in Guinea: A prospective diagnostic accuracy study. Infect. Dis. Poverty 2023, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Sinton, M.C.; Girard, A.; Ogunsola, J.; Chandrasegaran, P.; Capewell, P.; Perona-Wright, G.; Ngoyi, D.M.; Kuispond, N.; Bucheton, B.; Camara, M.; et al. Interleukin-17 drives sex-dependent weight loss and changes in feeding behaviour during Trypanosoma brucei infection. bioRxiv 2022. [Google Scholar] [CrossRef]
- Machado, H.; Bizarra-Rebelo, T.; Costa-Sequeira, M.; Trindade, S.; Carvalho, T.; Rijo-Ferreira, F.; Rentroia-Pacheco, B.; Serre, K.; Figueiredo, L.M. Trypanosoma brucei triggers a broad immune response in the adipose tissue. PLoS Pathog. 2021, 17, e1009933. [Google Scholar] [CrossRef]
- Norreen-Thorsen, M.; Struck, E.C.; Öling, S.; Zwahlen, M.; Von Feilitzen, K.; Odeberg, J.; Lindskog, C.; Pontén, F.; Uhlen, M.; Dusart, P.J. A human adipose tissue cell-type transcriptome atlas. Cell Rep. 2022, 40, 111046. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose tissue in control of metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Lafontan, M. Advances in adipose tissue metabolism. Int. J. Obes. 2008, 32, S39–S51. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.W.; Dixit, V.D. Adipose tissue as an immunological organ. Obesity 2015, 23, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. What we talk about when we talk about fat. Cell 2014, 156, 20–44. [Google Scholar] [CrossRef] [PubMed]
- Frontini, A.; Cinti, S. Distribution and development of brown adipocytes in the murine and human adipose organ. Cell Metab. 2010, 11, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Maretich, P.; Kajimura, S. The common and distinct features of brown and beige adipocytes. Trends Endocrinol. Metab. 2018, 29, 191–200. [Google Scholar] [CrossRef]
- Trindade, S.; De Niz, M.; Costa-Sequeira, M.; Bizarra-Rebelo, T.; Bento, F.; Dejung, M.; Narciso, M.V.; López-Escobar, L.; Ferreira, J.; Butter, F. Slow growing behavior in African trypanosomes during adipose tissue colonization. Nat. Commun. 2022, 13, 7548. [Google Scholar] [CrossRef] [PubMed]
- Sinton, M.C.; Chandrasegaran, P.R.; Capewell, P.; Cooper, A.; Girard, A.; Ogunsola, J.; Perona-Wright, G.; M Ngoyi, D.; Kuispond, N.; Bucheton, B. IL-17 signalling is critical for controlling subcutaneous adipose tissue dynamics and parasite burden during chronic murine Trypanosoma brucei infection. Nat. Commun. 2023, 14, 7070. [Google Scholar] [CrossRef]
- Mogk, S.; Meiwes, A.; Shtopel, S.; Schraermeyer, U.; Lazarus, M.; Kubata, B.; Wolburg, H.; Duszenko, M. Cyclical appearance of African trypanosomes in the cerebrospinal fluid: New insights in how trypanosomes enter the CNS. PLoS ONE 2014, 9, e91372. [Google Scholar] [CrossRef]
- Monroy, F.; Dusanic, D. The kidney form of Trypanosoma musculi: A distinct stage in the life cycle? Parasitol. Today 2000, 16, 107–110. [Google Scholar] [CrossRef]
- Masocha, W.; Rottenberg, M.E.; Kristensson, K. Migration of African trypanosomes across the blood–brain barrier. Physiol. Behav. 2007, 92, 110–114. [Google Scholar] [CrossRef]
- Kennedy, P.G. Human African trypanosomiasis–neurological aspects. J. Neurol. 2006, 253, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Buguet, A.; Mpanzou, G.; Bentivoglio, M. Human African trypanosomiasis: A highly neglected neurological disease. In Neglected Tropical Diseases and Conditions of the Nervous System; Springer: New York, NY, USA, 2014; pp. 165–181. [Google Scholar]
- Siletti, K.; Hodge, R.; Mossi Albiach, A.; Lee, K.W.; Ding, S.-L.; Hu, L.; Lönnerberg, P.; Bakken, T.; Casper, T.; Clark, M. Transcriptomic diversity of cell types across the adult human brain. Science 2023, 382, eadd7046. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Wu, X.; Jiang, L.; Zhang, Y. Single-cell RNA-seq reveals hypothalamic cell diversity. Cell Rep. 2017, 18, 3227–3241. [Google Scholar] [CrossRef] [PubMed]
- Mhlanga, J.D.; Bentivoglio, M.; Kristensson, K. Neurobiology of cerebral malaria and African sleeping sickness. Brain Res. Bull. 1997, 44, 579–589. [Google Scholar] [CrossRef]
- Coles, J.A.; Myburgh, E.; Ritchie, R.; Hamilton, A.; Rodgers, J.; Mottram, J.C.; Barrett, M.P.; Brewer, J.M. Intravital imaging of a massive lymphocyte response in the cortical dura of mice after peripheral infection by trypanosomes. PLoS Negl. Trop. Dis. 2015, 9, e0003714. [Google Scholar] [CrossRef] [PubMed]
- De Vlaminck, K.; Van Hove, H.; Kancheva, D.; Scheyltjens, I.; Antunes, A.R.P.; Bastos, J.; Vara-Perez, M.; Ali, L.; Mampay, M.; Deneyer, L. Differential plasticity and fate of brain-resident and recruited macrophages during the onset and resolution of neuroinflammation. Immunity 2022, 55, 2085–2102.e2089. [Google Scholar] [CrossRef]
- Stock, A.D.; Der, E.; Gelb, S.; Huang, M.; Weidenheim, K.; Ben-Zvi, A.; Putterman, C. Tertiary lymphoid structures in the choroid plexus in neuropsychiatric lupus. JCI Insight 2019, 4, e124203. [Google Scholar] [CrossRef]
- Pikor, N.B.; Prat, A.; Bar-Or, A.; Gommerman, J.L. Meningeal tertiary lymphoid tissues and multiple sclerosis: A gathering place for diverse types of immune cells during CNS autoimmunity. Front. Immunol. 2016, 6, 657. [Google Scholar] [CrossRef]
- Curtis, J.L. Cell-mediated adaptive immune defense of the lungs. Proc. Am. Thorac. Soc. 2005, 2, 412–416. [Google Scholar] [CrossRef]
- Gopallawa, I.; Dehinwal, R.; Bhatia, V.; Gujar, V.; Chirmule, N. A four-part guide to lung immunology: Invasion, inflammation, immunity, and intervention. Front. Immunol. 2023, 14, 1119564. [Google Scholar] [CrossRef]
- Borger, J.G. Spatiotemporal Cellular Networks Maintain Immune Homeostasis in the Lung. EMJ Respir. 2020, 8, 108–119. [Google Scholar] [CrossRef]
- Mabille, D.; Dirkx, L.; Thys, S.; Vermeersch, M.; Montenye, D.; Govaerts, M.; Hendrickx, S.; Takac, P.; Van Weyenbergh, J.; Pintelon, I. Impact of pulmonary African trypanosomes on the immunology and function of the lung. Nat. Commun. 2022, 13, 7083. [Google Scholar] [CrossRef] [PubMed]
- Grant, D.M. Detoxification pathways in the liver. J. Inherit. Metab. Dis. 1991, 14, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Langhans, W. Role of the liver in the control of glucose-lipid utilization and body weight. Curr. Opin. Clin. Nutr. Metab. Care 2003, 6, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar] [CrossRef] [PubMed]
- Jenne, C.N.; Kubes, P. Immune surveillance by the liver. Nat. Immunol. 2013, 14, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Knolle, P.A.; Gerken, G. Local control of the immune response in the liver. Immunol. Rev. 2000, 174, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Q.; Reid, S.A.; Fung, M.-C.; Inoue, N.; Lun, Z.-R. Analysis of gene expression profiles in the liver and spleen of mice infected with Trypanosoma evansi by using a cDNA microarray. Parasitol. Res. 2009, 104, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-Q.; Luckins, A.; Lun, Z.-R. Trypanosoma brucei brucei: A comparison of gene expression in the liver and spleen of infected mice utilizing cDNA microarray technology. Exp. Parasitol. 2011, 128, 256–264. [Google Scholar] [CrossRef]
- Liu, G.; Fu, Y.; Yosri, M.; Chen, Y.; Sun, P.; Xu, J.; Zhang, M.; Sun, D.; Strickland, A.B.; Mackey, Z.B. CRIg plays an essential role in intravascular clearance of bloodborne parasites by interacting with complement. Proc. Natl. Acad. Sci. 2019, 116, 24214–24220. [Google Scholar] [CrossRef]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Authié, E. Trypanosomiasis and trypanotolerance in cattle: A role for congopain? Parasitol. Today 1994, 10, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Bull, H.; Zhou, X.; Tabel, H. Intradermal infections of mice by low numbers of african trypanosomes are controlled by innate resistance but enhance susceptibility to reinfection. J. Infect. Dis. 2011, 203, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Ndungu, K.; Thungu, D.; Wamwiri, F.; Mireji, P.; Ngae, G.; Gitonga, P.; Mulinge, J.; Auma, J.; Thuita, J. Route of inoculation influences Trypanosoma congolense and Trypanosoma brucei brucei virulence in Swiss white mice. PLoS ONE 2019, 14, e0218441. [Google Scholar] [CrossRef] [PubMed]
- Caljon, G.; Van Den Abbeele, J.; Stijlemans, B.t.; Coosemans, M.; De Baetselier, P.; Magez, S. Tsetse fly saliva accelerates the onset of Trypanosoma brucei infection in a mouse model associated with a reduced host inflammatory response. Infect. Immun. 2006, 74, 6324–6330. [Google Scholar] [CrossRef] [PubMed]
- Caljon, G.; De Ridder, K.; De Baetselier, P.; Coosemans, M.; Van Den Abbeele, J. Identification of a tsetse fly salivary protein with dual inhibitory action on human platelet aggregation. PLoS ONE 2010, 5, e9671. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Getahun, M.N.; Ngiela, J.; Makwatta, J.O.; Ahuya, P.; Simon, T.K.; Kamau, S.K.; Torto, B.; Masiga, D. Metabolites from trypanosome-infected cattle as sensitive biomarkers for animal trypanosomosis. Front. Microbiol. 2022, 13, 2517. [Google Scholar] [CrossRef]
- Vincent, I.M.; Daly, R.; Courtioux, B.; Cattanach, A.M.; Bieler, S.; Ndung’u, J.M.; Bisser, S.; Barrett, M.P. Metabolomics identifies multiple candidate biomarkers to diagnose and stage human African trypanosomiasis. PLoS Negl. Trop. Dis. 2016, 10, e0005140. [Google Scholar] [CrossRef]
- Anderson, L.; Seilhamer, J. A comparison of selected mRNA and protein abundances in human liver. Electrophoresis 1997, 18, 533–537. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef]
- Pratt, J.M.; Petty, J.; Riba-Garcia, I.; Robertson, D.H.; Gaskell, S.J.; Oliver, S.G.; Beynon, R.J. Dynamics of protein turnover, a missing dimension in proteomics. Mol. Cell. Proteom. 2002, 1, 579–591. [Google Scholar] [CrossRef]
- International Human Genome Sequencing Consortium. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Hegde, P.S.; White, I.R.; Debouck, C. Interplay of transcriptomics and proteomics. Curr. Opin. Biotechnol. 2003, 14, 647–651. [Google Scholar] [CrossRef]
- Nie, L.; Wu, G.; Culley, D.E.; Scholten, J.C.; Zhang, W. Integrative analysis of transcriptomic and proteomic data: Challenges, solutions and applications. Crit. Rev. Biotechnol. 2007, 27, 63–75. [Google Scholar] [CrossRef]
- Haider, S.; Pal, R. Integrated analysis of transcriptomic and proteomic data. Curr. Genom. 2013, 14, 91–110. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Trypanosome Species | Host | Organ | Targeted Cells | Outcome of Infection | Technology | References |
|---|---|---|---|---|---|---|
| Trypanosoma b. rhodesiense | Human | Blood/Cerebral spinal fluid | Not available | Upregulation of gene expression related to the innate immune response in the early stage of infection. Upregulation of gene expression related to the anti-inflammation and neuro-degeneration in the late stage of infection. | RNA-seq (Illumina NextSeq500 System) | [56] |
| Trypanosoma (b.) evansi | Mouse | Spleen | B cells (transitional B cells, B1 cells, atypical memory B cells, marginal zone B cells, follicular B cells, germinal center-like B cells, plasma cells) | Rapid activation of mature B cells (GC-like B cells, atypical memory B cells, and plasma B cells). Depletion of the entire B cell compartment. Improved parasitemia control and maintenance of the B cell compartment in the infected AID−/− mice. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq6000 System) | [107] |
| Trypanosoma (b.) evansi | Mouse | Spleen | CD4+ T cells (naïve CD4+ T cells, Th1 cells, regulatory T cells, T follicular helper cells, cycling T cells, myeloid cell phenotype expressing T cells) | Th1 cells simultaneously produce IFN-γ and IL-10. Prediction of the role of T follicular helper cells in abrogation of germinal center formation. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq6000 System) | [96] |
| Trypanosoma (b.) evansi | Mouse | Spleen | Macrophages (monocytes- derived macrophages, erythroblast island macrophages, tissue resident macrophages), Erythroid cells (early and late erythroid precursor cells, pro-erythroblast, basophilic erythroblast, polychromatophilic erythroblast, orthochromatic erythroblast) | Tissue-resident macrophages simultaneously express markers of both classic and alternatively activated cells. Inhibition of the production of fully mature red blood cells during extramedullary erythropoiesis. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq6000 System) | [132] |
| Trypanosoma (b.) evansi | Mouse | Bone marrow | B cells (early B lineage cells) | Loss of B cell replenishment. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq 6000 System) | [107] |
| Trypanosome b. brucei | Mouse | Bone marrow | Neutrophils (pre-neutrophils, immature neutrophils, mature neutrophils) | No significant effect on the overall pattern of neutrophil differentiation. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq 6000 System) | [119] |
| Trypanosoma b. brucei | Mouse | Spleen | Neutrophils (pre-neutrophils, immature neutrophils, mature neutrophils), Plasma cells | Expansion of splenic neutrophil subpopulations and classification in four previously defined subpopulations. Prevention of organ damage, increase in plasma cells and prolonged host survival after neutrophil depletion. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq 6000 System) | [119] |
| Trypanosome b. brucei | Mouse | Adipose Tissue | Neutrophils, Macrophages, Monocytes, CD4+ T cells, CD8+ T cells, B cells | Lymphocytes produce type 1 effector cytokines, namely IFN-γ and TNF-α. The accumulation of antigen-specific IgM and IgG antibodies in adipose tissue. Inducing a broad and robust immune response in adipose tissue, requiring the complement system to reduce parasite burden locally, and emphasizing the critical role of T and/or B cells and the signature cytokine (Ifng−/−) in the adaptive immune system’s response. | RNA-Seq (Illumina HiSeq 2000 System) | [149] |
| Trypanosome b. brucei | Mouse | Adipose Tissue | Adipocyte, T cells (Th17 cells, CD27− Vɣ6+ cells) | IL-17, derived from Th17 and Vγ6+ cells, orchestrates the immune response, culminating in adipose tissue wasting and weight loss. | RNA-Seq (Illumina NovaSeq 6000 System), scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq 6000 System) | [158] |
| Trypanosome b. brucei | Mouse | Brain | Myeloid (homeostatic microglia, monocytes, border-associated macrophages, infection-associated mononuclear phagocytes), T cells (follicular-like regulatory CD4+ T cells, cytotoxic CD8+ T cells), B cells (CD138+ plasma cells) | Glial responses to infection occur near circumventricular organs, including the lateral and third ventricles. Crosstalk between homeostatic microglia and CD138+ plasma cells, facilitated by IL-10 and B cell activating factor (BAFF) signaling. | scRAN-seq with spatial transcriptomics (10× Genomic Chromium Controller/ Illumina NextSeq 550 System) | [82] |
| Trypanosome b. brucei | Mouse | Brain | Meningeal fibroblasts, CD21/35+ follicular dendritic cells, CXCR5+ PD1+ T follicular helper-like phenotype, GL7+ CD95+ GC-like B cells, plasmablasts/plasma cells | Immune landscape remodeling in the meninges containing T follicular helper cells-like T cells, GL7+ CD95+ GC-like B cells, and plasmablasts/plasma cells. Accumulation of meningeal autoreactive B cells that generate autoantibodies recognizing a broad range of host antigens, including myelin basic protein. | scRNA-seq (10× Genomic Chromium Controller/ Illumina NovaSeq 6000 System) | [116] |
| Trypanosome b. brucei | Mouse | Lung | T cells (γδ T cells), NK cells, Macrophages, B cells | The presence of trypanosomes is non-detrimental to lung function. Co-infections could affect susceptibility for opportunistic infections. Negative regulation of T cell activation, strong induction of γδ T cell. B cell memory depletion. Strong macrophage polarization towards classic activation. | NanoString nCounter (nCounter MAX Analysis System) | [174] |
| Trypanosoma b. brucei | Mouse | Skin | IL-17A-producing Vγ6+ cells | Expansion of dermal IL-17A-producing Vγ6+ cells (γδ T cell subsets) in subcutaneous adipose tissue. Potential interactions between subcutaneous adipocytes and Vγ6+ cells. | scRAN-seq with spatial transcriptomics (10× Genomic Chromium Controller/ Illumina NovaSeq6000 System) | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, B.; Vu, H.T.; Vu, H.T.; Radwanska, M.; Magez, S. Advances in the Immunology of the Host–Parasite Interactions in African Trypanosomosis, including Single-Cell Transcriptomics. Pathogens 2024, 13, 188. https://doi.org/10.3390/pathogens13030188
Choi B, Vu HT, Vu HT, Radwanska M, Magez S. Advances in the Immunology of the Host–Parasite Interactions in African Trypanosomosis, including Single-Cell Transcriptomics. Pathogens. 2024; 13(3):188. https://doi.org/10.3390/pathogens13030188
Chicago/Turabian StyleChoi, Boyoon, Hien Thi Vu, Hai Thi Vu, Magdalena Radwanska, and Stefan Magez. 2024. "Advances in the Immunology of the Host–Parasite Interactions in African Trypanosomosis, including Single-Cell Transcriptomics" Pathogens 13, no. 3: 188. https://doi.org/10.3390/pathogens13030188