
Inactivated Flagellin-Containing Vaccine Efficacy against Ovine Enzootic Abortion
 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
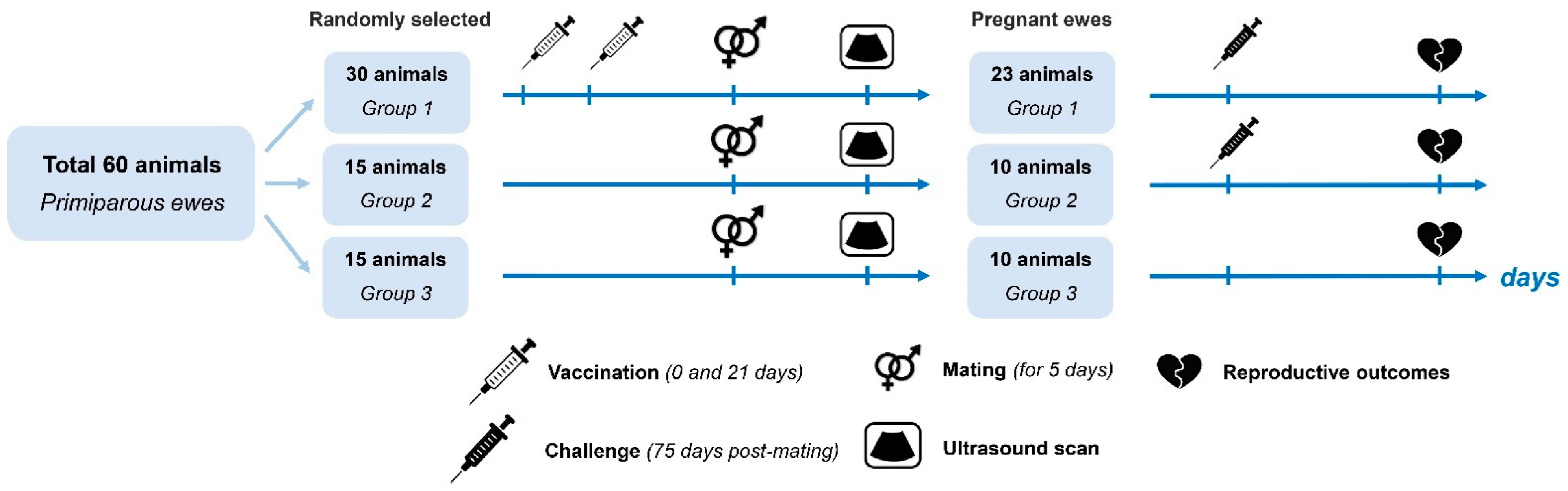
2. Materials and Methods
2.1. Animals
2.2. Statement of Ethics
2.3. C. abortus Strain
2.4. Obtaining the Vaccine Antigen
2.5. Recombinant Flagellin: Profile, Content in the Vaccine
2.6. Specific Antibody Detection
2.7. C. abortus DNA Detection
2.8. Ewe and Lamb Physiological Data Monitoring
2.9. Statistical Analysis
3. Results
3.1. Physiological Status Follow-Up on Ewes after Vaccination
3.2. Detection of Specific Antibodies after Vaccination and Infection with Homologous Strain
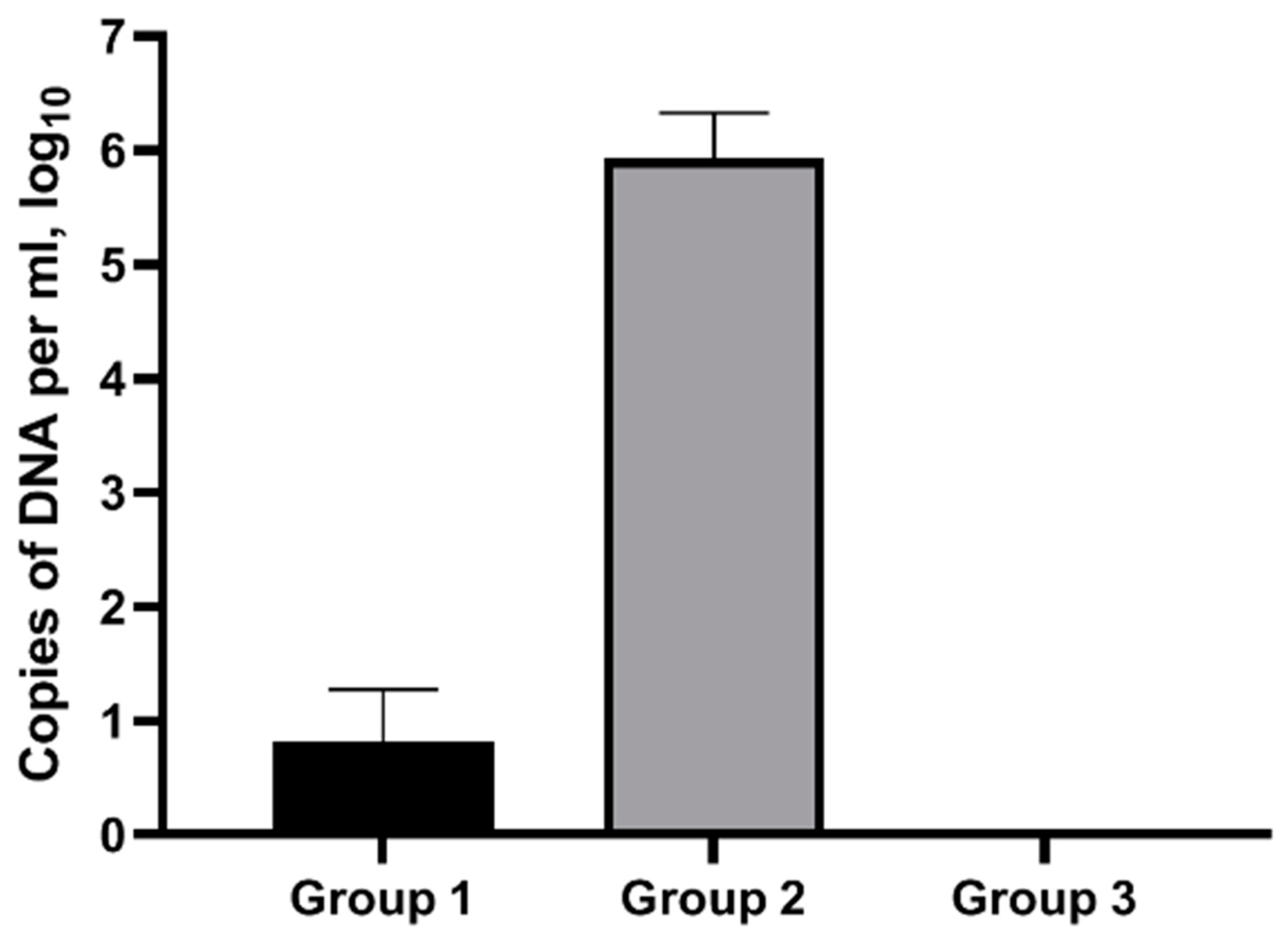
3.3. Impact of Infective Dose of C. abortus Virulent Srtain on Animals
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Halbert, G. Diseases of Sheep. Can. Vet. J. 2008, 49, 702. [Google Scholar]
- Longbottom, D.; Coulter, L.J. Animal Chlamydioses and Zoonotic Implications. J. Comp. Pathol. 2003, 128, 217–244. [Google Scholar] [CrossRef]
- Rodolakis, A.; Mohamad, K.Y. Zoonotic Potential of Chlamydophila. Vet. Microbiol. 2010, 140, 382–391. [Google Scholar] [CrossRef]
- Pospischil, A.; Thoma, R.; Hilbe, M.; Grest, P.; Gebbers, F.O. Abortion in Woman Caused by Caprine Chlamydophila abortus (Chlamydia psittaci serovar 1). Swiss Med. Wkly. 2002, 132, 64–66. [Google Scholar] [CrossRef]
- Walder, G.; Hotzel, H.; Brezinka, C.; Gritsch, W.; Tauber, R.; Würzner, R.; Ploner, F. An unusual cause of sepsis during pregnancy: Recognizing infection with chlamydophila abortus. Obstet. Gynecol. 2005, 106, 1215–1217. [Google Scholar] [CrossRef] [PubMed]
- Montbrau, C.; Fontseca, M.; March, R.; Sitja, M.; Benavides, J.; Ortega, N.; Caro, M.R.; Salinas, J. Evaluation of the Efficacy of a New Commercially Available Inactivated Vaccine against Ovine Enzootic Abortion. Front. Vet. Sci. 2020, 7, 593. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.E.; Jones, K.A.; Machell, J.; Brebner, J.; Anderson, I.; How, S. Efficacy Trials with Tissue-Culture Grown, Inactivated Vaccines against Chlamydial Abortion in Sheep. Vaccine 1995, 13, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Caro, M.R.; Ortega, N.; Buendia, A.J.; Gallego, M.C.; Del Río, L.; Cuello, F.; Salinas, J. Protection Conferred by Commercially Available Vaccines against Chlamydophila abortus in a Mouse Model. Vet. Rec. 2001, 149, 492–493. [Google Scholar] [CrossRef] [PubMed]
- García de la Fuente, J.N.; Gutiérrez-Martín, C.B.; Ortega, N.; Rodríguez-Ferri, E.F.; Del Río, M.L.; González, O.R.; Salinas, J. Efficacy of Different Commercial and New Inactivated Vaccines against Ovine Enzootic Abortion. Vet. Microbiol. 2004, 100, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Rodolakis, A.; Souriau, A. Response of Goats to Vaccination with Temperature-Sensitive Mutants of Chlamydia psittaci Obtained by Nitrosoguanidine Mutagenesis. Am. J. Vet. Res. 1986, 47, 2627–2631. [Google Scholar] [PubMed]
- Chalmers, W.S.K.; Simpson, J.; Lee, S.J.; Baxendale, W. Use of a Live Chlamydial Vaccine to Prevent Ovine Enzootic Abortion. Vet. Rec. 1997, 141, 63–67. [Google Scholar] [CrossRef]
- Livingstone, M.; Wattegedera, S.R.; Palarea-Albaladejo, J.; Aitchison, K.; Corbett, C.; Sait, M.; Wilson, K.; Chianini, F.; Rocchi, M.S.; Wheelhouse, N.; et al. Efficacy of Two Chlamydia abortus Subcellular Vaccines in a Pregnant Ewe Challenge Model for Ovine Enzootic Abortion. Vaccines 2021, 9, 898. [Google Scholar] [CrossRef]
- Tan, T.W.; Herring, A.J.; Anderson, I.E.; Jones, G.E. Protection of Sheep against Chlamydia psittaci Infection with a Subcellular Vaccine Containing the Major Outer Membrane Protein. Infect. Immun. 1990, 58, 3101–3108. [Google Scholar] [CrossRef]
- Regulation 2019/1010/EU of the European Parliament and of the Council of 5 June 2019 on the Alignment of Reporting Obligations in the Field of Legislation Related to the Environment, and Amending Regulations (EC) No 166/2006 and (EU) No 995/2010 of the European Parliament and of the Council, Directives 2002/49/EC, 2004/35/EC, 2007/2/EC, 2009/147/EC and 2010/63/EU of the European Parliament and of the Council, Council Regulations (EC) No 338/97 and (EC) No 2173/2005, and Council Directive 86/278/EEC. Off. J. Eur. Union 2019, 170, 115–127. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32019R1010&qid=1685043653814 (accessed on 23 March 2024).
- Council, N.R. Guide for the Care and Use of Laboratory Animals; The National Academies Press: Washington, DC, USA, 2010. [Google Scholar]
- Matveeva, I.; Nikitin, N.; Evtushenko, E.; Markova, E.; Yeremets, N. Chlamydia abortus Isolation and Identification in Aborted Ovine Fetus in Mari El Republic of Russia. Pathogens 2022, 11, 1408. [Google Scholar] [CrossRef]
- Livingstone, M.; Wheelhouse, N.; Maley, S.W.; Longbottom, D. Molecular Detection of Chlamydophila abortus in Post-Abortion Sheep at Oestrus and Subsequent Lambing. Vet. Microbiol. 2009, 135, 134–141. [Google Scholar] [CrossRef]
- McClenaghan, M.; Herring, A.J.; Aitken, I.D. Comparison of Chlamydia psittaci Isolates by DNA Restriction Endonuclease Analysis. Infect. Immun. 1984, 45, 384–389. [Google Scholar] [CrossRef]
- Hobson, D.; Johnson, F.W.A.; Byng, R.E. The Growth of the Ewe Abortion Chlamydial Agent in McCoy Cell Cultures. J. Comp. Pathol. 1977, 87, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Bavoil, P.; Ohlin, A.; Schachter, J. Role of Disulfide Bonding in Outer Membrane Structure and Permeability in Chlamydia trachomatis. Infect. Immun. 1984, 44, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, H.D.; Kromhout, J.; Schachter, J. Purification and Partial Characterization of the Major Outer Membrane Protein of Chlamydia trachomatis. Infect. Immun. 1981, 31, 1161–1176. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, H.D.; Schachter, J. Antigenic Analysis of the Major Outer Membrane Protein of Chlamydia spp. Infect. Immun. 1982, 35, 1024–1031. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, D.; Livingstone, M.; Aitchison, K.D.; Imrie, L.; Manson, E.; Wheelhouse, N.; Inglis, N.F. Proteomic Characterisation of the Chlamydia abortus Outer Membrane Complex (COMC) Using Combined Rapid Monolithic Column Liquid Chromatography and Fast MS/MS Scanning. PLoS ONE 2019, 14, e0224070. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ou, C.; Tian, D.; Ling, Y.; Pan, Q.; He, Q.; Eko, F.O.; He, C. Evaluation of an mpA-based Phage-Mediated DNA Vaccine against Chlamydia abortus in Piglets. Int. Immunopharmacol. 2013, 16, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ling, Y.; Liu, W.; Clark, J.R.; March, J.B.; Yang, J.; He, C. Protection of Mice against Chlamydophila abortus Infection with a Bacteriophage-mediated DNA Vaccine Expressing the Major Outer Membrane Protein. Vet. Immunol. Immonopathol. 2011, 144, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Héchard, C.; Grépinet, O.; Rodolakis, A. Protection Evaluation against Chlamydophila abortus Challenge by DNA Vaccination with a dnaK-encoding Plasmid in Pregnant and Non-pregnant Mice. Vet. Res. 2002, 33, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Héchard, C.; Grépinet, O.; Rodolakis, A. Evaluation of Protection against Chlamydophila abortus Challenge after DNA Immunization with the Major Outer-Membrane Protein-Encoding Gene in Pregnant and Non-pregnant Mice. J. Med. Microbiol. 2003, 52, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Longbottom, D.; Livingstone, M. Vaccination against Chlamydial Infections of Man and Animals. Vet. J. 2006, 171, 263–275. [Google Scholar] [CrossRef]
- Rodolakis, A.; Laroucau, K. Chlamydiaceae and Chlamydial Infections in Sheep or Goats. Vet. Microbiol. 2015, 181, 107–118. [Google Scholar] [CrossRef]
- Caspe, S.G.; Livingstone, M.; Frew, D.; Aitchison, K.; Wattegedera, S.R.; Entrican, G.; Palarea-Albaladejo, J.; McNeilly, T.N.; Milne, E.; Sargison, N.D.; et al. The 1B Vaccine Strain of Chlamydia abortus Produces Placental Pathology Indistinguishable from a Wild Type Infection. PLoS ONE 2020, 15, e0242526. [Google Scholar] [CrossRef]
- Entrican, G.; Wheelhouse, N.; Wattegedera, S.R.; Longbottom, D. New Challenges for Vaccination to Prevent Chlamydial Abortion in Sheep. Com. Immunol. Microbiol. 2012, 35, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Igietseme, J.U.; Black, C.M.; Caldwell, H.D. Chlamydia Vaccines: Strategies and Status. BioDrugs 2002, 16, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Batteiger, B.E.; Rank, R.G.; Bavoil, P.M.; Soderberg, L.S.F. Partial Protection against Genital Reinfection by Immunization of Guinea-Pigs with Isolated Outer-Membrane Proteins of the Chlamydial Agent of Guinea-Pig Inclusion Conjunctivitis. J. Gen. Microbiol. 1993, 139, 2965–2972. [Google Scholar] [CrossRef]
- De Sa, C.; Souriau, A.; Bernard, F.; Salinas, J.; Rodolakis, A. An Oligomer of the Major Outer Membrane Protein of Chlamydia psittaci is Recognized by Monoclonal Antibodies which Protect Mice from Abortion. Infect. Immun. 1995, 63, 4912–4916. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Theodor, I.; Peterson, E.M.; de la Maza, L.M. Immunization with an Acellular Vaccine Consisting of the Outer Membrane Complex of Chlamydia trachomatis Induces Protection against a Genital Challenge. Infect. Immun. 1997, 65, 3361–3369. [Google Scholar] [CrossRef] [PubMed]
- Herring, A.J. The function of the major outer membrane protein (MOMP) in the biology and pathogenicity of Chlamydia. Alpe Adria Microbiol. J. 1996, 5, 223–232. [Google Scholar]
- Herring, A.J.; Jones, G.E.; Dunbar, S.M.; Nettleton, P.F.; Fitzgerald, T.A.; Anderson, I.E.; Chapman, S.N.; Wilson, T.M.A. Recombinant vaccines against Chlamydia psittaci—An overview of results using bacterial expression and a new approach using a plant virus ‘overcoat’ system. In Proceedings of the Ninth International Symposium on Human Chlamydial Infection, San Francisco, CA, USA, 21–26 June 1998; Stephens, R.S., Byrne, G.I., Christiansen, G., Clarke, I.N., Grayston, J.T., Rank, R.G., Ridgway, G.L., Saikku, P., Schachter, J., Stamm, W.E., Eds.; International Chlamydia Symposium: San Francisco, CA, USA, 1998; pp. 434–437. [Google Scholar]
- Huleatt, J.W.; Jacobs, A.R.; Tang, J.; Desai, P.; Kopp, E.B.; Huang, Y.; Song, L.; Nakaar, V.; Powell, T.J. Vaccination with recombinant fusion proteins incorporating Toll-like receptor ligands induces rapid cellular and humoral immunity. Vaccine 2007, 25, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a Vaccine Adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef]
- Gupta, S.K.; Bajwa, P.; Deb, R.; Chellappa, M.M.; Dey, S. Flagellin a Toll-like Receptor 5 Agonist as an Adjuvant in Chicken Vaccines. Clin. Vaccine Immunol. 2014, 21, 261–270. [Google Scholar] [CrossRef]
- Cheng, C.; Jain, P.; Bettahi, I.; Palm, S.; Tifrea, D.; de la Mazam, L.M. A TLR2 agonist is a more effective adjuvant for a Chlamydia major outer membrane protein vaccine than ligands to other TLR and NOD receptors. Vaccine 2011, 29, 6641–6649. [Google Scholar] [CrossRef]
- Honko, A.N.; Sriranganathan, N.; Lees, C.J.; Mizel, S.B. Flagellin is an Effective Adjuvant for Immunization against Lethal Respiratory Challenge with Yersinia pestis. Infect. Immun. 2006, 74, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Mizel, S.B.; Graff, A.H.; Sriranganathanm, N.; Ervin, S.; Lees, C.J.; Lively, M.O.; Hantgan, R.R.; Thomas, M.J.; Wood, J.; Bell, B. Flagellin-F1-V Fusion Protein is an Effective Plague Vaccine in Mice and Two Species of Nonhuman Primates. Clin. Vaccine Immunol. 2009, 16, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Bargieri, D.Y.; Rosa, D.S.; Braga, C.J.M.; Carvalho, B.O.; Costa, F.T.; Espindola, N.M.; Vaz, A.J.; Soares, I.S.; Ferreira, L.C.; Rodrigues, M.M. New Malaria Vaccine Candidates Based on the Plasmodium vivax Merozoite Surface Protein-1 and the TLR-5 Agonist Salmonella Typhimurium FliC Flagellin. Vaccine 2008, 26, 6132–6142. [Google Scholar] [CrossRef]
- Lee, S.E.; Kim, S.Y.; Jeong, B.C.; Kim, Y.R.; Bae, S.J.; Ahn, O.S.; Lee, J.J.; Song, H.C.; Kim, J.M.; Choy, H.E.; et al. A Bacterial Flagellin, Vibrio vulnificus FlaB, Has a Strong Mucosal Adjuvant Activity To Induce Protective Immunity. Infect. Immun. 2006, 74, 694–702. [Google Scholar] [CrossRef]
- Skountzou, I.; Martin, M.; Wang, B.; Ye, L.; Kutsonanos, D.; Weldon, W.; Jacob, J.; Compans, R. Salmonella Flagellins are Potent Adjuvants for Intranasally Administered Whole Inactivated Influenza Vaccine. Vaccine 2010, 28, 4103–4112. [Google Scholar] [CrossRef]
- McDonald, W.F.; Huleatt, J.W.; Foellmer, H.G.; Hewitt, D.; Tang, J.; Desai, P.; Price, A.; Jacobs, A.; Takahashi, V.N.; Huang, Y.; et al. A West Nile Virus Recombinant Protein Vaccine that Coactivates Innate and Adaptive Immunity. J. Infect. Dis. 2007, 195, 1607–1617. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Group 1 | Group 2 | Group 3 | |
|---|---|---|---|
| Number of ewes | 23 | 10 | 10 |
| Percentage of live lambs, % | 93.1 | 15.2 | 94.6 |
| Total lambs (viable lambs) | 73 (68) | 33 (5) | 37 (35) |
| Abortions | 0 | 6 | 0 |
| Number of deliveries/deliveries with all live lambs | 23/21 | 4/3 | 10/9 |
| Early deliveries | 2 | 3 | 0 |
| Number of non-viable or stillborn lambs | 5 | 28 | 2 |
| Lamb’s weight at birth | 4.1 ± 0.54 | 3.45 ± 0.34 | 4.4 ± 0.46 |
| Pregnancy duration | 140.5 ± 5.7 | 131.1 ± 9.8 | 145.2 ± 5.2 |
| Group of Ewes | Group 1 | Group 3 |
|---|---|---|
| Group 2 | 0.0006 | 0.022 |
| Group 1 | 1 |
| Group of Ewes | Group 1 | Group 3 |
|---|---|---|
| Group 2 | 0.0003 × 10−11 | 0.0006 × 10−8 |
| Group 1 | 1 |
| Number of Fetuses | Number of Animals | ||
|---|---|---|---|
| Group 1 | Group 2 | Group 3 | |
| 1 | 5 | 3 | 1 |
| 2 | 6 | 1 | 3 |
| 3 | 4 | 2 | 1 |
| 4 | 0 | 0 | 0 |
| 5 | 4 | 2 | 3 |
| 6 | 4 | 2 | 2 |
| Total | 73 | 33 | 37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruglova, M.; Nikitin, N.; Evtushenko, E.; Matveeva, I.; Mazurov, A.; Pavlenko, I.; Popova, V.; Bogomolova, O.; Vasilyev, S.; Markova, E.; et al. Inactivated Flagellin-Containing Vaccine Efficacy against Ovine Enzootic Abortion. Pathogens 2024, 13, 277. https://doi.org/10.3390/pathogens13040277
Kruglova M, Nikitin N, Evtushenko E, Matveeva I, Mazurov A, Pavlenko I, Popova V, Bogomolova O, Vasilyev S, Markova E, et al. Inactivated Flagellin-Containing Vaccine Efficacy against Ovine Enzootic Abortion. Pathogens. 2024; 13(4):277. https://doi.org/10.3390/pathogens13040277
Chicago/Turabian StyleKruglova, Maria, Nikolai Nikitin, Ekaterina Evtushenko, Irina Matveeva, Aleksandr Mazurov, Igor Pavlenko, Vera Popova, Olesya Bogomolova, Stepan Vasilyev, Evgeniya Markova, and et al. 2024. "Inactivated Flagellin-Containing Vaccine Efficacy against Ovine Enzootic Abortion" Pathogens 13, no. 4: 277. https://doi.org/10.3390/pathogens13040277