Biological Control of Escherichia coli O157:H7 in Dairy Manure-Based Compost Using Competitive Exclusion Microorganisms

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Competitive Exclusion Microorganism Isolation and Culture Conditions
2.3. Species Identification by Amplifying the 16S rRNA Gene
2.4. Compost Inoculation, Sampling and Bacterial Enumeration
2.4.1. E. coli O157:H7 Growth under Laboratory Conditions
2.4.2. E. coli O157:H7 Growth under Greenhouse Conditions
2.5. Statistical Analysis
3. Results
3.1. Isolation and Identification of CE Bacteria against E. coli O157:H7
3.2. Effectiveness of CE Treatment on the Growth Reduction in E. coli O157:H7 in Compost under Laboratory Conditions
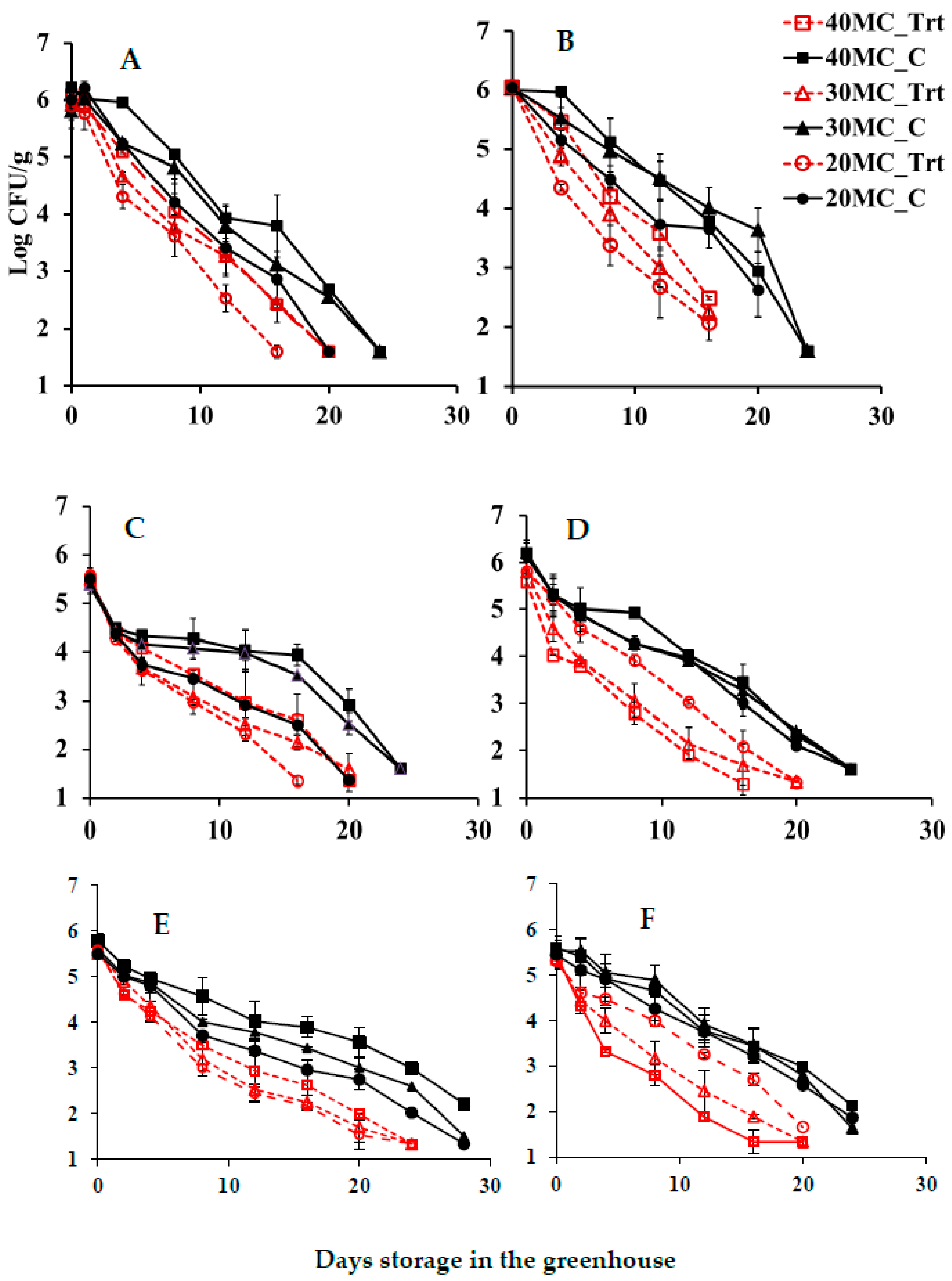
3.3. Effectiveness of CE Treatment on the Growth Reduction in E. coli O157:H7 in Compost under Greenhouse Conditions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyle, M.P.; Erickson, M.C. Summer meeting 2007—The problems with fresh produce: An overview. J. Appl. Microbiol. 2008, 105, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Pell, A.N. Manure and microbes: Public and animal health problem? J. Dairy. Sci. 1997, 80, 2673–2681. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Centers for Disease Control and Prevention (CDC). Plesiomonas shigelloides and Salmonella serotype Hartford infections associated with contaminated water supply-Livingston County, New York. Morb. Mortal. Wkly. Rep. 1996, 47, 394–396. [Google Scholar]
- Clements, D.; Acuña-Maldonado, L.; Fisk, C.; Stoeckel, D.; Wall, G.; Woods, K.; Bihn, E. FSMA Produce Safety Rule: Documentation Requirements for Commercial Soil Amendment Suppliers. 2019. Available online: https://ucanr.edu/sites/Small_Farms_/files/315707.pdf (accessed on 2 April 2024).
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef] [PubMed]
- Blanc, M.; Marilley, L.; Beffa, T.; Aragno, M. Thermophilic bacterial communities in hot composts as revealed by most probable number counts and molecular (16S rDNA) methods. FEMS Microbiol. Ecol. 1999, 28, 141–149. [Google Scholar] [CrossRef]
- Brinton, W.F., Jr.; Evans, E. Standardized test for evaluation of compost self-heating. BioCycle 1995, 36, 64. [Google Scholar]
- Sobsey, M.D.; Khatib, L.A.; Hill, V.R.; Alocilja, E.; Pillai, S. Pathogens in animal wastes and the impacts of waste management practices on their survival, transport and fate. In White Papers on Animal Agriculture and the Environment; MidWest Plan Service (MWPS); Iowa State University: Ames, IA, USA, 2001; Chapter 17. [Google Scholar]
- Anderson, I.C.; Cairney, J.W.G. Diversity and ecology of soil fungal communities: Increased understanding through the application of molecular techniques. Environ. Microbiol. 2004, 6, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Freilich, S.; Zarecki, R.; Eilam, O.; Segal, E.S.; Henry, C.S.; Kupiec, M.; Gophna, U.; Sharan, R.; Ruppin, E. Competitive and cooperative metabolic interactions in bacterial communities. Nat. Commun. 2011, 2, 589. [Google Scholar] [CrossRef] [PubMed]
- Mead, G.C. Prospects for “competitive exclusion” treatment to control salmonellas and other foodborne pathogens in poultry. Vet. J. 2000, 158, 111–123. [Google Scholar] [CrossRef]
- Doyle, M.P.; Erickson, M.C. Opportunities for mitigating pathogen contamination during on-farm food production. Int. J. Food Microbiol. 2012, 152, 54–74. [Google Scholar] [CrossRef]
- Zhao, T.; Podtburg, T.C.; Zhao, P.; Schmidt, B.E.; Baker, D.A.; Cords, B.; Doyle, M.P. Control of Listeria spp. by Competitive-Exclusion Bacteria in Floor Drains of a Poultry Processing Plant. Appl. Environ. Microbiol. 2006, 72, 3314–3320. [Google Scholar] [CrossRef] [PubMed]
- Wray, C.; Davies, R.H. Competitive exclusion--an alternative to antibiotics. Vet. J. 2000, 159, 107–108. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Luo, F.; Jiang, X. Thermal Inactivation of Heat-Shocked Escherichia coli O157:H7, Salmonella, and Listeria monocytogenes in Dairy Compost. J. Food Prot. 2010, 73, 1633–1640. [Google Scholar] [CrossRef] [PubMed]
- Uhlich, G.A.; Reichenberger, E.R.; Cottrell, B.J.; Fratamico, P.; Andreozzi, E. Whole-Genome Sequence of Escherichia coli Serotype O157:H7 Strain B6914-ARS. Genome Announc. 2017, 5, e01191-17. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Harris, L.J.; Bender, J.; Bihn, E.A.; Blessington, T.; Danyluk, M.D.; Delaquis, P.; Goodridge, L.; Ibekwe, A.M.; Ilic, S.; Kniel, K.; et al. A framework for developing research protocols for evaluateon of microbial hazards and controls during production that pertain to the quality of agricultural water contacting fresh produce that may be consumed raw. J. Food Prot. 2012, 75, 2251–2273. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Benedict, R.C.; Palumbo, S.A. Protection against heat-injury in Staphylococcus aureus by solutes. J. Food Prot. 1982, 45, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Jiang, X. Isolation and Characterization of Competitive Exclusion Microorganisms from Animal Wastes-Based Composts against Listeria monocytogenes. J. Appl. Microbiol. 2022, 132, 4531–4543. [Google Scholar] [CrossRef]
- Olofsson, T.C.; Ahrne, S.; Molin, G. The bacterial flora of vacuum-packed coldsmoked salmon stored at 7 °C, identified by direct 16S rRNA gene analysis and pure culture technique. J. Appl. Microbiol. 2007, 103, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Wichuk, K.M.; McCartney, D. A review of the effectiveness of current time–temperature regulations on pathogen inactivation during composting. J. Environ. Eng. Sci. 2007, 6, 573–586. [Google Scholar] [CrossRef]
- Avcı, A.; Üzmez, S.; Alkan, F.B.; Bagana, İ.; Nurçeli, E.; Çiftçi, E. Antimicrobial activity spectrums of some Bacillus strains from various sources. GIDA 2016, 41, 523–528. [Google Scholar] [CrossRef]
- Watabe, M.; Rao, J.R.; Murphy, A.R.; Moore, J.E. Inhibition of Listeria ivanovii by Paenibacillus lentimorbus isolated from phase II mushroom compost. World J. Microbiol. Biotechnol. 2003, 19, 875–877. [Google Scholar] [CrossRef]
- Yin, H.-B.; Chen, C.-H.; Gu, G.; Nou, X.; Patel, J. Pre-Harvest Biocontrol of Listeria and Escherichia coli O157 on Lettuce and Spinach by Lactic Acid Bacteria. Int. J. Food Microbiol. 2023, 387, 110051. [Google Scholar] [CrossRef] [PubMed]
- Al-Ghafri, H.M.; Velazhahan, R.; Shahid, M.S.; Al-Sadi, A.M. Antagonistic Activity of Pseudomonas aeruginosa from Compost against Pythium aphanidermatum and Fusarium solani. Biocontrol Sci. Technol. 2020, 30, 642–658. [Google Scholar] [CrossRef]
- Burd, G.; Dixon, D.G.; Glick, B.R. Plant growth-promoting bacteria that decrease heavy metal toxicity in plants. Can. J. Microbiol. 2000, 46, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Gonciarz, R.L.; Renslo, A.R. Emerging role of ferrous iron in bacterial growth and host-pathogen interaction: New tools for chemical (micro)biology and antibacterial therapy. Curr. Opin. Chem. Biol. 2021, 61, 170–178. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- No. FDA-2011-N-0921; Standards for the Growing, Harvesting, Packing, and Holding of Produce for Human Consumption (Proposed Rule); FDA Food Safety Modernization Act (FSMA), 21 CFR Parts 16 and 112. US-Food and Drug Agenecy: Silverspring, MD, USA, 2013.
- Russ, C.F.; Yanko, W.A. Factors affecting salmonellae repopulation in composted sludges. Appl. Environ. Microbiol. 1981, 41, 597–602. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim, J.; Jiang, X. The growth potential of Escherichia coli O157:H7, Salmonella spp., and Listeria monocytogenes in dairy manure-based compost in a greenhouse setting under different seasons. J. Appl. Microbiol. 2010, 109, 2095–2104. [Google Scholar] [CrossRef]
- Aertsen, A.; Vanoirbeek, K.; De Spiegeleer, P.; Sermon, J.; Hauben, K.; Farewell, A.; Nyström, T.; Michiels, C.W. Heat shock protein-mediated resistance to high hydrostatic pressure in Escherichia coli. Appl. Environ. Microbiol. 2004, 70, 2660–2666. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tran, T.D.; Kwon, H.Y.; Kim, E.H.; Kim, K.W.; Briles, D.E.; Pyo, S.; Rhee, D.K. Decrease in penicillin susceptibility due to heat shock protein ClpL in Streptococcus pneumoniae. Antimicrob. Agents Chemother. 2011, 55, 2714–2728. [Google Scholar] [CrossRef]

{kind=link}
| E. coli O157:H7 Growth Reduction (log CFU/g) in Compost with Moisture Content (%) of | |||
|---|---|---|---|
| Temperature (°C) | 20 | 30 | 40 |
| 22 | 1.05 * | 2.59 | 3.36 |
| 30 | 2.12 | 2.25 | 2.57 |
| Season | Moisture (%) | Time (d) Required to Have Difference (p < 0.05) in Pathogen Reduction between Treatment and Control | |
|---|---|---|---|
| Approach #1 * | Approach #2 | ||
| Summer | 40 | 8 | 8 |
| 30 | 8 | 8 | |
| 20 | 8 | 12 | |
| Fall | 40 | 2 | 12 |
| 30 | 2 | 12 | |
| 20 | 16 | 16 | |
| Winter | 40 | 2 | 8 |
| 30 | 2 | 12 | |
| 20 | 16 | 16 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Wang, J. Biological Control of Escherichia coli O157:H7 in Dairy Manure-Based Compost Using Competitive Exclusion Microorganisms. Pathogens 2024, 13, 361. https://doi.org/10.3390/pathogens13050361
Jiang X, Wang J. Biological Control of Escherichia coli O157:H7 in Dairy Manure-Based Compost Using Competitive Exclusion Microorganisms. Pathogens. 2024; 13(5):361. https://doi.org/10.3390/pathogens13050361
Chicago/Turabian StyleJiang, Xiuping, and Jingxue Wang. 2024. "Biological Control of Escherichia coli O157:H7 in Dairy Manure-Based Compost Using Competitive Exclusion Microorganisms" Pathogens 13, no. 5: 361. https://doi.org/10.3390/pathogens13050361
APA StyleJiang, X., & Wang, J. (2024). Biological Control of Escherichia coli O157:H7 in Dairy Manure-Based Compost Using Competitive Exclusion Microorganisms. Pathogens, 13(5), 361. https://doi.org/10.3390/pathogens13050361