Abstract
Background: The advent of the hepatitis B vaccine has achieved tremendous success in eradicating and reducing the burden of hepatitis B infection, which is the main culprit for hepatocellular carcinoma—one of the most fatal malignancies globally. Response to the vaccine is achieved in about 90–95% of healthy individuals and up to only 50% in immunocompromised patients. This review aimed to provide an overview of hepatitis B vaccine non-response, the mechanisms involved, B cell amnesia, and strategies to overcome it. Methods: Databases, including Google Scholar, PubMed, Scopus, Cochrane, and ClinicalTrials.org, were used to search and retrieve articles using keywords on hepatitis B vaccine non-response and B cell amnesia. The PRISMA guideline was followed in identifying studies, screening, selection, and reporting of findings. Results: A total of 133 studies on hepatitis B vaccine non-response, mechanisms, and prevention/management strategies were included in the review after screening and final selection. Factors responsible for hepatitis B vaccine non-response were found to include genetic, immunological factors, and B cell amnesia in healthy individuals. The genetic factors were sex, HLA haplotypes, and genetic polymorphisms in immune response markers (cytokines). Non-response was common in conditions of immunodeficiency, such as renal failure, haemodialysis, celiac disease, inflammatory bowel disease, hepatitis C co-infection, and latent hepatitis B infection. Others included diabetes mellitus and HIV infection. The mechanisms involved were impaired immune response by suppression of response (T helper cells) or induced suppression of response (through regulatory B and T cells). Discussion: A comprehensive and careful understanding of the patient factors and the nature of the vaccine contributes to developing effective preventive measures. These include revaccination or booster dose, vaccine administration through the intradermal route, and the use of adjuvants in the vaccine.
1. Introduction
Hepatitis B is one of the major infections contributing to the global burden of disease, with an estimated prevalence of over 350 million people with the chronic form of the disease, characterised by the presence of the surface antigen of hepatitis B (HBsAg) in the blood beyond six months, and resulting in approximately one million deaths annually [1]. It represents the main culprit in the development of liver cancer, also referred to as hepatocellular carcinoma and liver cirrhosis, accounting for about 50% and 33% of mortality, respectively, which is the third leading cause of mortality in cancer after lung and colorectal cancers [2]. In addition, it also leads to fulminant hepatitis on some rare occasions (approximately 1%) [3]. The development of a recombinant vaccine for hepatitis B has yielded significant success in reducing the burden of chronic infection with hepatitis B, especially in countries with wider coverage from childhood to adulthood [4]. The immune response to the hepatitis B vaccine is determined by assessing the concentration of neutralising antibody (anti-HBsAg), and the optimum level required is ≥10 IU/L, though only individuals that produce ≥100 IU/L are considered high responders in the UK, while individuals that produce the antibody within the range of ≥10 IU/L and ≥100 IU/L are regarded as low responders, meaning that their immunity period after vaccination will be less than that of high responders [5,6,7]. Despite the success achieved in the use of the vaccine, an adequate immune response is not achieved in about 5–10% of the population, especially healthcare personnel, and is higher in some vulnerable groups, such as immunocompromised individuals and chronic kidney disease patients on haemodialysis, where up to 50% non-response may occur. Non-response is also common in other conditions, such as celiac disease, diabetes mellitus, hepatitis C infection, use of immunomodulators, and in individuals with some genetic dispositions, etc. [8]. Again, evidence confirms that non-response to the hepatitis B vaccine is not only related to vaccinee-associated factors such as age, sex, obesity, and smoking, but may also depend on the nature of the vaccine, dose, and type of adjuvants used [9,10]. The non-response, if not curtailed, can lead to an enhanced occurrence of chronic hepatitis B infection, resulting in a higher risk of complications as well as transmission. Multiple research studies have been carried out to study the factors contributing to the non-response, ranging from epidemiological factors to defective immunological mechanisms and genetic polymorphisms involving antigen-presenting cells, cytokines, and cytokine receptors. This review is aimed at providing an overview of hepatitis B vaccine non-response: prevalence and occurrence in various disease conditions, mechanisms involved, relationship with B cell amnesia, and approaches to management in healthy and/or diseased population groups.
2. Methods
2.1. Search
The search for articles was carried out in the following databases: Google Scholar, PubMed, Web of Science, Cochrane, and clinicaltrials.org, with the keywords: “hepatitis B vaccine”, “non-response/hypo-response/non-responders”, “immunogenicity”, and “B cell amnesia”, using Boolean operators. The studies retrieved were published between 1989 and 2023. In this study, 133 were selected and included after screening and comparing against eligibility criteria. The lists of references of the articles selected were also explored for additional articles (Figure 1).
Figure 1.
Summary flowchart of identification, screening, and selection of studies for the review.
2.2. Eligibility Criteria
Criteria for inclusion included studies carried out in persons who have been vaccinated against the hepatitis B virus; studies that explore the possible causes of immune non-response to the vaccine against hepatitis B and the relationship between the immune non-response and B cell amnesia; studies that investigate potential mechanisms behind non-responsiveness, such as lymphocytes and cytokine expression levels; studies that explore the potential strategies for improving immune response to the vaccine, such as increased vaccination dose, intradermal administration, and the use of adjuvanted vaccines; studies that consider the possible pros and cons of these strategies, and any other factors that may influence the vaccine response.
The exclusion criteria include studies not published in English, studies that do not investigate the causes of immune non-response to the hepatitis B vaccine, studies that do not explore potential mechanisms or approaches to improve the response to the vaccine, or studies that do not provide complete data. Information was extracted from the studies that fulfilled the inclusion criteria using a data extraction form, and the data were synthesised using a narrative synthesis approach.
The protocol for the review was drafted and published in PROSPERO, an international database for prospective registration of systematic review protocols, with registration number: PROSPERO 2023 CRD42023457503, which is now available from: https://www.crd.york.ac.uk/PROSPERO/display_record.php?RecordID=457503 (accessed on 23 September 2023).
3. Results
3.1. Hepatitis B Vaccine Non-Response Prevalence and Risk Factors
Despite global efforts to reduce the impact of chronic hepatitis B infection through vaccination, which has been made part of the routine childhood immunisation in many countries, failure has been encountered in achieving absolute protection. This failure has been due to increasing reports of immune non-response to the vaccine. Non-response to the hepatitis B vaccine with the normal three-dose series has been reported in many studies within the range of 5–10% or 10–15% within healthy individuals [11,12]. Non-response is caused by a variety of factors, including advanced age, obesity, smoking, chronic illness, and hereditary factors [13]. The risk increases in individuals with other conditions, as explained below.
3.1.1. Chronic Hepatitis C
The presence of chronic hepatitis C infection significantly increases the severity of chronic hepatitis B, leading to a higher risk or actual occurrence of complications (cirrhosis of the liver and hepatocellular carcinoma) and a reduction in the immune response to the hepatitis B vaccine [14]. There was conflicting evidence on the immunogenicity of the hepatitis B vaccine in individuals infected with chronic hepatitis C [15]. Non-response in hepatitis C co-infection is generally higher (10–15%) than in the general population (5–10%) [16]. A recent finding observed that co-infection of hepatitis B with chronic hepatitis C in adults after treatment with direct-acting antiretroviral drugs had a significant non-response rate (43%), which was also associated with advanced age and the occurrence of isolated HbcAb or latent hepatitis B infection [17]. The mechanisms contributing to the immune non-response to the hepatitis B vaccine in hepatitis C include the involvement of Tim-3 (T cell immunoglobulin mucin domain-3) produced by monocytes, which is upregulated, resulting in an imbalance of IL-12/IL-23 (IL-12 inhibition and IL-23 enhancement) in the innate immune system and, consequently, accumulating Th17 in the adaptive immune system [18]. Another finding also confirmed the involvement of the programmed death receptor-1 (PD-1), usually expressed on the T helper cells, which is usually upregulated in chronic hepatitis C infection, thereby interfering with the function of lymphocytes in responding to the HBV vaccine due to dysregulation [19].
3.1.2. Chronic Kidney Disease (CKD) and Haemodialysis
CKD and haemodialysis patients are well known to be more vulnerable to complications of hepatitis B infection, such as cirrhosis and hepatocellular carcinoma, as they are highly susceptible to hepatitis B infection, likely due to increased administration of blood products in haemodialysis, contamination of equipment used for dialysis, and other physiological and environmental factors [20]. The risk is usually reduced with routine vaccination and effective haemodialysis [21,22]. Factors contributing to non-response, especially in haemodialysis, are multiple in number [23]. Some studies have observed a link between non-response in haemodialysis patients with conditions including hepatitis C, obesity, diabetes mellitus, nutrition, advanced age, and duration of haemodialysis [24]. IL-12, a potent regulatory cytokine that plays a significant role in hepatitis B virus clearance, was observed to be relatively lower in non-responders with chronic kidney disease undergoing dialysis [25]. Also, a significant interaction was observed using “chi-square automatic interaction detection” between gender, dose of vaccine, and dialysis frequency, with gender (male) being most important, especially when combined with a higher dialysis frequency [26]. Studies performed in patients with chronic kidney disease also identified a link between non-response to the hepatitis B vaccine and deficiency in vitamin D (both 25-hydroxycholciferol and 1,25-dihydroxycholecalciferol), as it plays a role in regulating the production of cytokines and enhancing the antimicrobial capacity of monocytes, neutrophils, and natural killer cells [27]. Also, a study on single-nucleotide polymorphisms of the vitamin D system, which include vitamin D receptor, vitamin D binding protein, and retinoic X receptor alpha, in chronic kidney disease patients associated only the rs1544410 polymorphism in the vitamin D receptor with non-response to the hepatitis B vaccine, which is more common with homozygotes “AA” and less common with homozygotes “GG” [28].
3.1.3. Celiac Disease
Hepatitis B vaccine non-response is significantly higher (50–70%) in celiac disease patients compared to normal individuals (5–10%) [29,30,31]. This makes them more susceptible to chronic hepatitis B infection depending on the patients’ age category, and level of diagnosis, and treatment [32]. The non-response has been associated with competition between HbsAg and glacial peptides in binding to HLA-DQ2, HLA-DQ8, HLA-DR3, and HLA-DR7 haplotypes, which leads to impairment of the response through antigen presentation, proliferation of T lymphocytes, and production of HbsAg-specific antibodies [33]. In these patients, the response may be improved by enhanced compliance with gluten-free diets and the administration of booster vaccinations [30,34,35].
3.1.4. Latent HBV Infection
Luo et al. [36] first performed a study in which non-responders to the hepatitis B vaccine were assessed and HBV DNA was detected in about 60–70% of them, indicating the presence of a latent HBV infection that may induce tolerance to the vaccine through immunosuppression, enhancement of the regulatory T cell activity, or inhibition of the response of B and T lymphocytes, as reported in other studies [37,38]. This is also referred to as “occult HBV infection”, which occurs after serorecovery from the infection with the maintenance of specific T cell activity that prevents reactivation [39]. Meanwhile, another finding indicated the reactivation of fulminant hepatitis B infection due to inflammatory bowel disease and treatment with infliximab [40,41].
3.1.5. Inflammatory Bowel Disease
Inflammatory bowel disease is a condition generally characterised by a state of immunosuppression as a result of using immunomodulators (corticosteroids, thiopurines, monoclonal antibodies, etc.) for treatment and the negative effect of the disease on the activity of the immune system. In addition, it involves many risk factors for transmission of hepatitis B virus, such as continuous blood transfusion, colonoscopy, surgery, and other parenteral medications [42]. As such, hepatitis B vaccination is included in the routine management of inflammatory bowel disease patients based on different standard guidelines [43]. Despite that, hepatitis B vaccine non-response has been observed to occur in about half of the vaccinated patients with IBD, likely due to the reasons listed above, which are more compounded in older patients [44,45,46]. Consequently, having considered non-response categories, new formulation types with special adjuvants, booster vaccination, and dose adjustment can be relatively effective in addressing the issue in this patient group [8,47]. Other conditions in which immunosuppressants are used, such as psoriasis (Th1 associated), atopic dermatitis (Th2 associated), and morphea, are highly susceptible to chronic infection, likely due to the nature of the disease condition and the drugs used [48]. Therefore, testing before the commencement of therapy is essential, as is the need for booster vaccination if needed.
3.1.6. Other Conditions
The immune response to the hepatitis B vaccine in diabetes, especially insulin-dependent diabetes mellitus (IDDM) in children, has been investigated and reported to be lower (average 58%) compared to their healthy counterparts (average 90%) [49,50]. This was in agreement with the report in a systematic review and meta-analysis confirming a generally weaker immune response to the hepatitis B vaccine in diabetics [51,52]. The non-response occurs as a result of the effect of diabetes on various components of the immune system, ranging from antigen processing and presentation to suppression of B cell responses. The presence of the DR3/DQ2 and DR4/DQ8 HLA haplotypes in about 90% of diabetics is also a strong marker of hepatitis B vaccine non-responsiveness [53]. HIV infection is also among the conditions in which HBV vaccine non-response is common. The prevalence among HIV patients has reached up to 44–76%, contrary to 5–10% in healthy people [54]. The major culprit for the non-response was a decreased level of CD4 T cells. Other cellular and molecular mechanisms include the upregulation of regulatory T cells (Tregs), leading to the release of IL-10, and subsequently deregulating the function of T and B cells through a complex mechanism [55,56].
3.2. Mechanisms Involved in Hepatitis B Vaccine Non-Response
3.2.1. Pattern Recognition Receptors
Pattern recognition receptors are an important player in the innate immune system. During the immune response to the hepatitis B virus, RIG-1 and TL3 play a significant role, and their functions might be impaired by the HBV polymerase [57]. Deficiency of TL3 in the placenta of HBsAg-positive mothers has been found to attenuate the vaccine response in infants, who are more vulnerable to chronic hepatitis infection, leading to deficiency in the cytokine levels in the innate immune system (IL-6, TNF-α, INF-α, and INF-γ, and Th1/Th2-specific (INF-γ, IL-4, and IL-10) cytokines), which are important in the immune response to the vaccine [58,59]. Other important pattern recognition receptors associated with the non-response are ficolin-2 and mannose-binding lectin, whose increases in concentration activate the lectin complement pathway [60]. This activation, conversely, results in bypassing the activity of the adaptive immune system and the development of immunological memory. Ficolin-2 and MBL also play a role in the pathogenesis of hepatitis B infection [61,62,63]. Mice with MBL deficiency produced a higher amount of hepatitis B antibodies than the wild-type mice, and when the MBL was reconstituted, the effect was abolished [64]. Also, a study from Indonesia revealed that a mutation of the MBL gene’s untranslated portion resulted in a non-response to the hepatitis vaccine [65]. Generally, within the innate immune system, elevated levels of proinflammatory and inflammatory genetic factors and cytokines at baseline predict a poor response to the hepatitis B vaccine, particularly in older individuals [66]. This was the first study to attempt to elucidate the mechanisms underlying non-response to the hepatitis B vaccine, especially at baseline.
3.2.2. Involvement of Helper T Cells (Th)
CD4+ T cells play a significant role in the production of antibodies during infection [67]. This process after hepatitis B vaccination is tightly regulated by both Th1 and Th2 [68]. Specifically, Th2 responses are essential for high anti-hepatitis B antibody titres by promoting B cell activation, differentiation, class switching, and the formation of memory B cells, all of which are critical for robust and sustained immunity [69]. The action of these helper cells (cell-mediated cytolytic control and humoral production of specific antibodies by Th1 and Th2, respectively) is mediated by specific cytokines, including IL-2, TNF-γ, and TGF produced by Th1, and IL-4, IL-5, IL-6, IL-10, and IL-13 produced by Th2 [70]. In agreement, Sabry et al. [71] discovered a significant inhibition in overall Th1-specific cytokines, and Honorati et al. [72] observed the same for TNF-γ (specific to Th1) and IL-3 (specific to Th2) in hepatitis B vaccine non-responders. However, other conflicting findings indicated involvement of only Th2-specific cytokines (IL-4 and IL-5) and Th0 (mixed Th1/Th2), with no TNF-γ and IL-2 [73], while Böcher et al. (1999), Doedée et al. (2016), and Vingerhoets et al. (1995) asserted the involvement of only IL-4, IL-5, and IL-10, which are Th1-specific, contrary to the predominant involvement of Th2 cytokines [67,69,74]. Effects of both defective Th1 and Th2 were observed due to lower levels of a wider range of Th1 and Th2-specific cytokines, viz., INF-γ, TNF-α, IL-2, IL-4, IL-10, and IL-12, indicating a generalised dysfunction of T-cell-mediated immune response [75]. Moreover, the differentiation of B cells into memory cells and plasma cells that produce high-affinity antibodies is also mediated by T follicular helper cells (Tfh) through a reaction at the germinal centre. The reaction also involved the action of B6 cells, a transcriptional repressor, and IL-12 [76,77]. The main characteristics of the Tfh cells at the germinal centre are the expression of co-stimulatory molecules (ICOS), programmed death 1 (PD-1), and chemokine receptor 5 (CXCR5), which is also expressed by the circulating Tfh (cTfh) available in and more easily extracted from peripheral blood [78,79]. For research purposes, the cTfh is generally utilised, as it is also capable of releasing IL-12, which is necessary for the maturation of B cells, differentiation to produce plasma cells, and the release of antibodies [80]. To explore the role of post-transcriptional regulation of Tfh differentiation mediated by miRNA, Xu et al. [81] investigated the role of miR17-92 involved in the differentiation of Tfh, induced during T cell activation and suppression after differentiation of the T cells. It was found that antibody production is promoted by the miR-17-92 cluster, while miR-18a and miR-17 play a role in the development of Tfh cells following hepatitis B immunisation. Th17, which is another subset of CD4+ T cells, also plays a regulatory function similar to IL-10 mediated by IL-17 and IL-22, thereby serving a complementary effect with Treg cells [82]. Activation of macrophages, recruitment of Th1, B cell maturation, and differentiation, are all linked to the level of Th17 cells in circulation [83]. IL-22 has been linked to the cellular immune response to the hepatitis B vaccine [84]. Thus, IL-17 and IL-22 have a significant protective role to play during hepatitis B infection and in the immune response to vaccines. CD150 is another signal regulatory molecule for lymphocyte activation that influences the function of T cells, especially CD4+ T cells. It was more expressed in a non-responders cohort compared to responders in an Indian [85] and Chinese [86] population. Investigation into genetic polymorphisms on IL-17 identified the role played by the lower frequency of the rs4711998 GG genotype in a non-response and non-significant relationship with the rs22275503, rs2227501, and rs1026786 genotypes of IL-22 [87].
Further investigation into the role of deltex1, a protein located at the transmembrane portion of T cells, and its role in T cell anergy has been conducted [88,89]. Ten SNPs were identified, and two of them (rs2384077 and rs10744794), which are situated at the first deltex1 intron, had a remarkable association with the degree of immunological response among infant and adult Han populations in China. Although the role of introns in transcription remains insignificant compared to the coding regions, SNPs in the first introns were found to be involved in transcriptional functions, as were the exons [90].
3.2.3. The Role of Regulatory B Cells
Regulatory B cells (Bregs) are a special category of cells in the immune system that inhibit the immune response through several mechanisms, which include activation of Treg cells, inhibition of CD+ Th cells, and inhibition of maturation and differentiation of plasma cells. This function is mediated through the regulation of the cytokine microenvironment, by the elevation of cytokines, such as IL-10, IL-35, and TGF-β, which have regulatory effects [91,92]. While the Bregs are known to be involved in several pathologies, such as cancer, multiple sclerosis, and systemic lupus erythromatosis [93], evidence shows variation in the elevated frequencies of two Bregs subpopulations, namely, CD24+/highCD27+ and CD24highCD38high, and enhanced IL-10 expression levels in hepatitis B non-responders [94,95]. The upregulated frequencies of CD24+/high CD27+ and CD24highCD38high were found to be reversed in non-responders, together with a lower expression level of IL-10 after receiving a booster dose with a recombinant third-generation vaccine for hepatitis B virus [96], which confirms the assertion by Bolther et al. (2018) [94] that the expression of IL-10-producing Bregs cells is not a good predictor of immunological response to the hepatitis B vaccination.
3.3. Genetic Variation in Hepatitis B Vaccine Non-Response
Variations in genetics have since been regarded as a major factor influencing the response to vaccination in more than 70% of non-responders [97], as vaccination of family relatives of known non-responders produces a higher non-response rate (58%) compared to the rate (5–10%) in the general population [98]. A study on gene expression identified about nine genes that were significantly upregulated in non-responders compared to responders, and these genes included nine coding RNAs, namely: transketolase-like 1 (TKTL1), carcino-embryonic antigen-related cell adhesion molecule 8 (CEACAM8), matrix metallopeptidase 8 (MMP8), bactericidal-permeability-increasing protein (BPI), folate receptor 3 (FOLR3), Defensin Alpha 1B (DEFA1B), Defensin Alpha 4 (DEFA4), Lactotransferrin (LTF), and transcobalamin 1 (TCN1), and the rest were non-coding genes involved in the regulation and expression process of CEACAM8. Three genes made of non-coding RNAs (a pseudogene and two long-non-coding RNAs) involved in the synthesis of proteins were downregulated [99]. These genes, which were present in the non-responding group, provided evidence of the presence of fixed genetic variations, which are independent of immunisation and could represent the transcriptome features of hepatitis B vaccine immune non-responders. miRNA-155, which is a small-non-coding RNA involved in several processes of immune response, was discovered at an increased amount in the serum of hepatitis B vaccine immune non-responders [100]. However, the non-response was more associated with SNPs in miR-26a-1 and miR-146a of the miRNA [100]. On another note, granulin gene (GRN) expression was significantly upregulated in non-responders, indicating a greater activation state of the pathway for neutrophil activation in innate immunity. This greater GRN expression in non-responders likely results in the production of a higher amount of smaller granulins that are known to enhance the expression of Il-8 and the recruitment of more neutrophils. The IFITM1 gene, essential for immune response signalling, was downregulated at baseline in responders, indicating a state of greater immune system activation in the non-responders. However, after vaccination, IFITM1 was upregulated in the responders. The mechanism behind this observation remains to be elucidated [101]. Generally, the gene expression profiles of responders reflected the expected B and T cell responses to the vaccine. After vaccination, no peak of differentially expressed genes (DEGs) was observed in non-responders. Ten DEGs were noticed, among which three (KCTD7, DCAF12, and MARCH 8) are related to immune response and are involved in what is referred to as the ubiquitination process. Marked downregulation of KCTD7 in non-responders is interestingly difficult to interpret, as no differences exist between responders and non-responders for the other corresponding components (DCAF12 and MARCH 8) of the ubiquitination process [102]. According to these data, it can be said that in non-responders, only a small number of immune pathways are stimulated by the hepatitis B vaccine, and this occurred at different stages and rates than in responders.
3.3.1. Antigen Processing and Presentation Genes
The immune response to vaccination involves the antigen processing and presentation to T cells, and antigen recognition by the T cells. The processing of the antigen and the presentation are mediated by the antigen-presenting cells (APC), containing a protein called the major histocompatibility complex (MHC). The communication that occurs between the APCs, B cells, and T cells is dependent on the function of surface molecules expressed on each of the cells, which include CD40, CD40L, CD25, and CD6 [103]. The genes on MHC DNA are important in the hepatitis B vaccine immune response, and it was found that the non-response gene is a recessive gene, which must exist in homozygous form to attenuate a response [104]. The first gene found to be responsible for the non-response was the [HLA-B8, SC01, DR3], especially in white people [105]. There is a significant heterogeneity in the expression levels of different HLA-I and HLA-II genes in antigen-presenting cells, particularly HLA-DRB5 and HLA-B, which are markedly downregulated [106]. This, in turn, affects antigen processing and presentation, as indicated in the “Kyoto Encyclopaedia of Genes and Genomes” (KEGG). Another role of the HLA was determined by assessing the influence of HLA-DPA1 and HLA-DPB1, where about nine alleles were found to be involved in keeping memory for the vaccine, which was determined after a booster vaccination [107]. Among the nine alleles, three were risk-associated while six were protective for the vaccine non-response [108]. The HLA-DPB1 alleles strongly related to non-response were determined to be at 05:01 and 09:01 after administering a booster vaccination in adolescent individuals with a history of active vaccination in the neonate period [107]. According to other findings aimed to provide more insight into the effect of genetic polymorphism in HLA-II alleles, DQB1-encoding β-portion on the antigen-binding groove of the DQ protein and DRB1 are also involved in the immune recognition and response. Ten different types of these alleles were found to be involved in the hepatitis B vaccine immune non-response [109]. DQB1*0401 was associated with non-response after vaccination with a three-dose schedule in a Japanese population [110]. Similarly, immune non-response was also associated with DRB1*07, according to studies from the USA [111] and Belgium [112], while a paradoxical finding was obtained in a study from Iran [113]. For DRB1*04, two studies from Taiwan [107] and Korea [114] provided evidence of its association with non-response. Conversely, a study in Germany involving pairs of twins associated DRB1*11 and DRB1*01 with a good immune response to the hepatitis B vaccination [115]. Another one from Taiwan also indicated that DRB1*08 favours a good response to booster vaccination. An investigation among the Chinese Han population identified a relationship between DRB1*14 and non- or hypo-response [116]. In summary, DRB1*01, DRB1*11, and DRB1*08, in contrast to DRB1*04 and DRB1*07, were in favour of non-response, as postulated in [117]. In a Japanese population, non- or hypo-response was significantly associated with HLA-DPB1*05:01, while a good response was associated with HLA-DPB1*04:02, HLA-DQB1*05:01, HLA-DRB1*01:01, and HLA-DRB1*08:03, as confirmed by the analysis of amino acid residues at different antigen-binding pockets of the HLA structural proteins [118]. The C4A locus of HLA class III was also found to be important, as it was downregulated in non-responders, especially HLA-C4AQ0, DRB1*0301, and DQB1*02 haplotypes, observed among non-responders and their close families [119,120]. These findings show that age, race, gender, and the nature of the virus may influence the functions of the genes involved. However, examining the influence of all these genes on the long-term immune response in those vaccinated early after birth revealed an insignificant relationship between these genes and the enduring immune response to the hepatitis B vaccination or booster vaccination [121]. Further genetic assessment of single-nucleotide polymorphisms (SNPs) by genome-wide association studies (GWAS) from different parts of the world, such as Japan, Thailand, China, Indonesia, and Korea, provided a holistic insight on the association of non-response with 10 different SNPs (rs2116260, rs3830066, rs5025825, rs35953215, rs7770370, rs6457709, rs35953215, rs3830066, rs7770370, rs3128961, rs9277535, rs9277542, and rs4282438) on loci HLA-DP. The strongest association was observed with rs7770370, which is also the major gene determining the response to booster dose, showing a significant effect on immunological memory in the long term [122]. Development of chronic hepatitis B infection and seroconversion was associated with rs9277535 and rs3077 on HLA-DP and rs2856718 and rs7453920 on HLA-DQ, in addition to their effect on non-response to hepatitis B vaccination [123,124,125,126].
3.3.2. T Cell Regulation Genes
In hepatitis B vaccine immune non-response, the genes involved in the activity of naive and memory T cells, such as the MAPK signalling pathway and Nf-kb, are downregulated, as are the genes involved in many activities, such as cell activation, oxidative phosphorylation, interferon-γ, calcium ion reactions, and the IL-4 production of the three T-cell subsets, which are the CD4+ effector cells, CD8+ effector T cells, and CD8+ effector memory T cells. Meanwhile, cytotoxicity-associated biomarkers in NK and NKT cells are upregulated [106]. Genetic polymorphism in genes related to the function of Tfh cells also contributes to hepatitis B vaccine non-response, especially SNPs rs3922, affecting the expression of CXCR5 and rs355687 in CXCL13. Moreover, the allele “A” of the rs3922 reduces the expression of the CXCR5 more than the corresponding allele “G” due to the higher non-response observed with the “GG” genotype, followed by the “AG”, than “AA” [127].
3.3.3. Genetic Polymorphism in Markers of the Immune System
Apart from the HLA genes, genetic polymorphism on the Th1 and Th2 cytokines and their receptors has largely been the focus of genomic research on hepatitis B vaccine non-response, owing to their essential contribution to the innate, cellular, and humoral immune systems. The effects of SNPs on various cytokines (IL-1β, IL4, IL4R, IL10, and IL12B) have been studied and reported in various populations from different parts of the globe [128,129,130]. For instance, the authors of [70] identified the role of IL12A and IL12B genes’ polymorphism in hepatitis B vaccine non-response. Also, a study on 53 SNPs on 21 different genes for toll-like receptors, cytokines, and their receptors, identified 4 SNPs (rs3804100 in TLR2, rs2243248 in the non-coding region of IL-4, rs1805015 in the coding region of IL-4RA, and rs1295686 in non-coding region of IL-13) strongly associated with non-response to the HBV vaccine [130]. Similarly, the INF-γ bioavailability and function are determined by the INFGR1 gene, whose genetic polymorphism, particularly the 170 and 182 allelic markers, is associated with hepatitis B vaccine immune non-response [131].
3.4. B Cell Amnesia-Associated Hepatitis B Vaccination Non-Response
The serum antibodies, despite being an essential measure for protection, are not the only indicators of successful immunisation. The generation of immunological memory is also an essential part of the immune response to vaccination [132]. The plasma cells generated during primary vaccination remain in the bone and produce antibodies constantly, while antibody detection in the serum long after vaccination is a marker for the activity of plasma cells that persist in circulation [133]. The plasma cells cannot detect antigens because they lack B cell receptors, which are present on the memory B cell surfaces. Thus, to induce the anamnestic response, B memory cells also generated from the germinal centre during the antigenic immune response are very necessary [134]. The memory B cells expressing a specific, high-affinity BCR upon encounter with their specific antigen, undergo differentiation into fresh plasma cells, and this accounts for the flare in high-affinity antibodies (IgG), typically described as an anamnestic response [135]. Some findings suggest independent regulation of plasma and memory B cells, hence the production of effective B cell memory in the absence of a defensive amount of anti-HbsAg antibodies [136,137]. Further proof of this is the non-progression to chronic infection by those in endemic areas without a sufficient level of antibodies after vaccination [138]. An examination of the distribution of specific lymphocytes identifies the same levels of memory B cells in both responders and non-responders, and the function of the memory B cells is maintained both in vivo and in vitro. Also, booster vaccination triggers no specific effect on the level of antibodies or memory B cell frequency [139]. However, in many cases, the memory B cell available in non-responders usually produces the same level of IgA and IgM, with a lower level of IgG, the detection of which depends on the type and sensitivity of the test method used [140]. This shows that even the non-responders, having supposed functional memory B cells, have a certain level of protection against chronic HBV infection [141]. The mechanism by which this happens remains an area for further research (Figure 2).
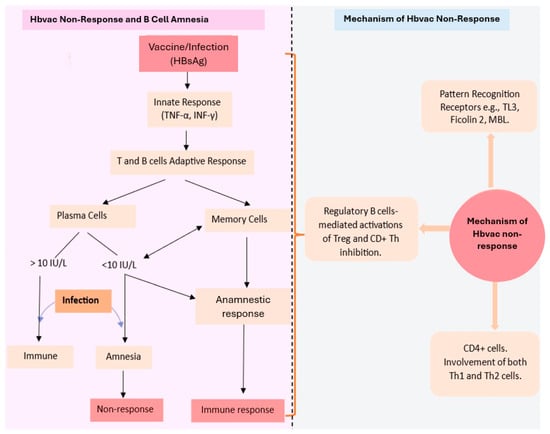
Figure 2.
Diagram illustrating the concept of hepatitis B vaccine non-response and B cell amnesia and the mechanisms involved in the hepatitis B vaccine non-response. Hbvac: hepatitis B vaccine; HBsAg: hepatitis B surface antigen; TNF-α: tumour necrotic factor; α INF-γ: interferon γ; IU/L: international unit per litre; TL3: toll-like receptor 3; MBL: mannose binding lectin; Th: T helper; Treg: regulatory T cells.
3.5. Strategies in Addressing HBV Non-Response
3.5.1. Revaccination/Booster Dose
Revaccination in normal non-responders to vaccination requires careful consideration in selecting the type of vaccine to be used, as the nature of the vaccine, dose, and frequency of vaccination affect seroconversion in previous non-responders [142]. For example, in randomised controlled trials comparing the effectiveness of various brands of vaccine in previous non-responders, HBVaxPRO-40 and Fendrix produced a more significant response than Engerix, Twinrix, and HBVaxPRO-10, which necessitates expanding the indication for these vaccines or careful choice of a vaccine to enhance immunisation of real or suspected non-responders [143,144]. The revaccination of non-responders is also referred to as booster vaccination, in which a single higher dose (up to 60 µg) or an entirely new vaccination course of three doses is administered again [145,146]. Interestingly, the single higher dose has been found to have high immunogenicity and safety in several clinical trials [147]. After the booster dose, a long-lasting immunity is achieved when the anti-HbsAg level is greater than 100 IU/L [148]. A higher vaccine dose (or double dose) during the first immunisation may be used to achieve a better response in patients at risk of non-response conditions, such as chronic hepatitis C infection [149]. In non-responders with chronic kidney disease undergoing haemodialysis, revaccination is a common practice, in addition to using a higher (40 µg) vaccine dose at increased frequency due to the significant rate of non-response among them [150]. Meanwhile, a revaccination may be unnecessary in some perceived non-responders, as seroprotection may be delayed, as reported in a case report of a patient with haematological malignancy (a smouldering multiple myeloma) [151]. However, an experimental error in the laboratory, an HBV infection, or booster vaccination might be a risk factor. Demographic indices, such as age, gender, BMI, alcohol, and smoking, might also affect the response to revaccination, as reported in many studies [152].
3.5.2. Use of Intradermal Route for Vaccine Administration
A change in the route of vaccine administration from the usual intramuscular route to the intradermal route has achieved considerable and comparable success in both the general population and special categories, such as people with chronic kidney disease, haemodialysis, celiac disease, and liver disease, though hypertension and advanced age limit the response in some of the categories [153]. As in celiac disease patients, a 40% higher response (anti-HbsAg ≥ 100 IU/L) was reported in booster vaccination through ID (Engerix B 2 μg), in contrast to IM (Engerix B 10 μg), in which only a 7.1% higher response was achieved [154]. In chronic liver disease patients, a significant response was achieved (≥10 IU/L in 70% of non-responders) with a high-dose intradermal vaccine (40 µg), without any increase in adverse events or dermatological reaction risk [155]. The reason for the enhanced response through the intradermal route compared to the intramuscular route could be the presence of dendritic and Langerhans cells, rich in MHC II molecules in human skin, warranting direct T cell activation, the release of inflammatory cytokines (especially IL-1) by keratinocytes that further enhance MHC-II expression on Langerhans cells, and the quick transfer of the antigen to the lymph node, where they persist for longer compared to IM, which is mostly localised [156,157]. This was also observed with vaccines for other infections, including influenza, smallpox, rabies, and polio [158]. Besides the enhanced immunogenicity, ID administration requires a lower vaccine dose and allows easy detection of immune responses, implying cost reduction, and recently, an enhanced delivery system using micro-needles [159]. However, most of the studies on the effectiveness of the ID route are not comprehensive, hence the need for studies that will focus on investigating the optimum dose, schedule of administration, and overall efficacy of the ID administration devices [160]. Meanwhile, a difference in response was also observed between administration through the gluteal muscle (yielding a lower response rate) and the deltoid muscle (yielding a higher rate of response) [161].
3.5.3. Use of Adjuvants
Aluminium salts were the first adjuvants used to enhance the immune response of the body to the HBV vaccination. They act by eliciting the Th2-mediated response, leading to an increased rate of antibody production, and poorly inducing the Th1-cell-mediated response to enhance viral elimination [162]. The aluminium salts have undergone several modifications to improve their adjuvant action, including a strong capacity for adsorption, stability of suspension, and improved cell-mediated immune response [163]. Th1- (INF-γ) and Th2 (IL-13)-specific cytokines may be utilised as vaccine adjuvants to boost antibody responses [72]. Levamisole, IL-2, and granulocyte-macrophage colony-stimulating factor (GM-SF) are commonly employed as vaccine adjuvants in recombinant hepatitis B vaccines [164]. However, granulocyte colony-stimulating factor (G-CSF) was found to be more safe and effective than the GM-CSF, likely as a result of its restricted effect on only neutrophils, in addition to stimulation of antigen-presenting cells and dendritic cells capable of inducing Th2 lymphocytes [165]. Meanwhile, the efficacy of the GM-SF was better in chronic kidney disease [166] and HIV disease patients [167] than in healthy non-responders, in whom booster vaccination with a double dose of the recombinant vaccine produced a better response [168]. A hydrogel delivery system produced with a co-polymer (monomethoxy poly(ethylene glycol)-co-poly(lactic-co-glycolic acid (mPEG-PLGA) hydrogel) that is biodegradable and thermosensitive has been used to enhance the delivery of the GM-CSF together with the HbsAg at a local site, where activation of the immune system occurs through the activation of cytokines and dendritic cells in non-response mice strains [169]. Liposomes of cationic DC-Chol lipid have also been found to be more effective than aluminium hydroxide as an adjuvant in non-response mice, by enhancing cell-mediated immune responses [170]. Various other vaccine adjuvants have been formulated for specific categories of individuals with additional epitopes of viral proteins to enhance the immune response. PreS1/S2 envelope protein antigens are added to recombinant vaccines from the Chinese hamster ovary, referred to as third-generation hepatitis B vaccines, used for non-responders to conventional yeast-derived recombinant vaccines, immunosuppressed patients, overweight individuals, and patients with renal failure [171]. The third-generation vaccine has been proven to be more immunogenic by inducing more robust T and B cell immune responses, with an acceptable safety profile despite a few more side effects of injection site reaction, which was considered mild [172,173]. The immunomodulatory action of β-glycosphingolipid mediated through natural killer T cells (NKT cells) has been tested, and a positive effect was obtained, which further buttressed the role of the NKT cells in the HBV immune response [174]. For kidney transplant recipients, a single Fendrix® dose, which is a third-generation vaccine containing the S and preS1/S2 antigens formulated with the adjuvant 3-O-desacyl-40-monophosphoryl lipid A (MPL), elicited a 60% response in previous non-responders after transplant, in addition to an enhanced hepatitis virus-specific cellular and humoral immune response [175]. This is comparable to the effect of conventional vaccines on healthy individuals. MPL adjuvant was also effective in patients with renal insufficiency who were previous non-responders, according to research carried out in Germany [176]. It was also formulated with Saponaria Molina to enhance the response after liver transplant [177]. These adjuvant vaccines are not usually used in healthy individuals due to concerns about side effects, difficulty in licensing highly immunogenic substances, and competing commercial interests among producers of conventional vaccines [178].
Another form of adjuvant is co-administration with other vaccines. The response to hepatitis B vaccines has been enhanced in non-responders and hypo-responders when administered together (in the same formulation) as a double dose with hepatitis A vaccine, in which a 90–95% response was achieved in previous non-responders [179]. The combined hepatitis B vaccine with the tetanus–diphtheria vaccine in dialysis patients who were non-responders also produced an enhanced but short-lived response [180], while the combination in healthy non-responders produced an enhanced but non-significant response compared to a single recombinant hepatitis B vaccine [181]. Another study compared the response of the combination in non-responders among pregnant women, haemodialysis patients, and healthy individuals that are non-responders and found that the response was enhanced more significantly in healthy non-responders compared to the other groups [182].
3.5.4. Strategies in Chronic Kidney Disease (CKD) Patients
In CKD patients, especially those under haemodialysis, strategies employed include an increase in vaccine dose and frequency, addition of special adjuvants, revaccination or addition of a booster dose, and changes in the route of administering the vaccine [183,184,185,186]. Supplementation with vitamin D is also an effective enhancer of the immune response, in addition to the protection it affords to chronic kidney patients against hyperthyroidism and other conditions [187] (Table 1).

Table 1.
Some common hepatitis B vaccines and their key features.
4. Discussion
Numerous factors, including the host immune status, defective immune cells, genetic variation, type, and nature of the vaccine, have been associated with non-response to the hepatitis B vaccine. The production and kinetics of cytokines from the peripheral blood mononuclear cells (PBMC) and their proliferation levels play a significant role in categorising individuals in relation to HBV vaccine response into high responders, low responders, non-responders, and naïve categories. The results of the present review indicate that non-response related to cell-mediated immunity is more CD4+-involved than CD8+, as hypothesised in an early report [188]. Meanwhile, the genetic mechanisms involved show that the non-response to the HBV vaccine involves a specific mechanism and does not indicate a general malfunction of the immune system, especially among healthy non-responders, where an adequate immunological response may be observed in the immune cells by stimulation with different antigens [189]. Also, in non-responders, despite the inadequate humoral response, there could be enough cellular response to protect them against the infection, as evidenced by the effect of the booster dose, revealing an anamnestic response. For genetic polymorphism, the majority of studies characterised non-responders as having specific alleles that are not present in responders, showing that the non-response alleles are markers for the genetic risk of immune pathologies, such as celiac disease, diabetes, and inflammatory bowel disease, and vice versa.
Strategies to curtail non-response include revaccination or booster vaccination, the use of adjuvants, and changing the route of administration. These strategies are sometimes influenced and determined by the nature of HLA haplotypes in the recipients. A proper understanding of the mechanisms and factors involved in the non-response significantly influenced the development of means and strategies to prevent the non-response, thereby reducing the burden of chronic hepatitis B infection. The mechanisms of non-response largely influenced by genetic polymorphism vary substantially among different populations around the globe, which necessitates a comprehensive understanding to allow proper implementation of the various prevention strategies. There seems to be a great paucity of information about the mechanisms of non-response from the West African populations, who are among the high pandemic areas with high vulnerability to chronic HBV infection and a poor healthcare system for effective management of the disease [190]. From the foregoing, B cell amnesia remains an area in need of further research to determine its cause and effect as it relates to hepatitis B vaccine non-response.
In conclusion, the non-response to the hepatitis B vaccine can be said to be multi-specific and polyclonal, therefore requiring a comprehensive understanding and systematic approach to mitigate it. This will facilitate the achievement of the WHO 2030 goal of achieving 90% global immunisation, a prevalence reduction to 0.1%, and an 80% improvement in treatment. Although the vaccine proves effective significantly, its role in the program needs to be reevaluated considering the various limitations discussed in this review.
Author Contributions
Conceptualisation, A.O.J. and S.A.H.; methodology, A.O.J. and S.A.H.; resources, M.U.I. and A.S.A.; data curation, N.B.; writing—original draft preparation, N.B.; writing—review and editing, S.A.H., A.S.A. and N.B.; visualisation, A.O.J. and M.U.I.; supervision, A.O.J. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- WHO. Global Progress Report on HIV, Viral Hepatitis and Sexually Transmitted Infections, 2021: Accountability for the Global Health Sector Strategies 2016–2021: Actions for Impact; World Health Organization: Geneva, Switzerland, 2021.
- Li, Q.; Cao, M.; Lei, L.; Yang, F.; Li, H.; Yan, X.; He, S.; Zhang, S.; Teng, Y.; Xia, C. Burden of liver cancer: From epidemiology to prevention. Chin. J. Cancer Res. 2022, 34, 554. [Google Scholar] [CrossRef]
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.; Bray, F. Observatory:“Cancer Today”; International Agency for Research on Cancer: Lyon, France, 2020. [Google Scholar]
- Yardeni, D.; Chang, K.-M.; Ghany, M.G. Current best practice in hepatitis B management and understanding long-term prospects for cure. Gastroenterology 2023, 164, 42–60.e46. [Google Scholar] [CrossRef]
- Gelinas, L.; Abu-Raya, B.; Ruck, C.; Cai, B.; Kollmann, T.R. Hepatitis B virus vaccine–induced cell-mediated immunity correlates with humoral immune response following primary vaccination during infancy. ImmunoHorizons 2017, 1, 42–52. [Google Scholar] [CrossRef]
- Rahmani, A.; Montecucco, A.; Kusznir Vitturi, B.; Debarbieri, N.; Dini, G.; Durando, P. Long-Term Effectiveness of Hepatitis B Vaccination in the Protection of Healthcare Students in Highly Developed Countries: A Systematic Review and Meta-Analysis. Vaccines 2022, 10, 1841. [Google Scholar] [CrossRef]
- Hudu, S.A.; Malik, Y.A.; Niazlin, M.T.; Harmal, N.S.; Adnan, A.; Alshrari, A.S.; Sekawi, Z. Antibody and immune memory persistence post infant hepatitis B vaccination. Patient Prefer. Adherence 2013, 7, 981. [Google Scholar] [CrossRef]
- Yanny, B.; Konyn, P.; Najarian, L.M.; Mitry, A.; Saab, S. Management approaches to hepatitis B virus vaccination nonresponse. Gastroenterol. Hepatol. 2019, 15, 93–99. [Google Scholar]
- Medeiros, R.P.; Terrault, N.A.; Mazo, D.F.; Oliveira, C.P.; Dodge, J.; Zitelli, P.M.; Lopes, M.H.; Carrilho, F.J.; Pessoa, M.G. Impaired anti-HBV vaccine response in non-cirrhotic chronic HCV is not overcome by double dose regimen: Randomized control trial. Ann. Hepatol. 2023, 28, 100891. [Google Scholar] [CrossRef]
- Hudu, S.A.; Shinkafi, S.H.; Umar, S. An overview of recombinant vaccine technology, adjuvants and vaccine delivery methods. Int. J. Pharm. Pharm. Sci. 2016, 8, 19–24. [Google Scholar] [CrossRef]
- Singh, R.; Kaul, R.; Kaul, A.; Khan, K. A comparative review of HLA associations with hepatitis B and C viral infections across global populations. World J. Gastroenterol. 2007, 13, 1770. [Google Scholar] [CrossRef]
- Gunasekaran, J.; Sree, P.N. Prevalence of Non-Responsiveness to an Indigenous Recombinant Hepatitis B Vaccine: A Study among Health Care Workers in A Tertiary Hospital. Int. J. Mod. Trends Sci. Technol. 2020, 6, 1–5. [Google Scholar] [CrossRef]
- Khafagy, A.; AlJahdaly, I.; Goweda, R. Hepatitis B Vaccine: Assessment of Immunologic Response, Coverage Rate, and Factors Influencing Seroreactivity. Clin. Lab. 2020, 66, 1351. [Google Scholar] [CrossRef]
- Sun, Y.-T.; Zhang, Y.-X.; Tang, H.; Mao, Q.; Wang, X.-Z.; Zhang, L.-Y.; Chen, H.; Zhong, Y.-N.; Lin, S.-M.; Zhang, D.-Z. Clinical characteristics and current management of hepatitis B and C in China. World J. Gastroenterol. 2014, 20, 13582. [Google Scholar] [CrossRef]
- Wiedmann, M.; Liebert, U.G.; Oesen, U.; Porst, H.; Wiese, M.; Schroeder, S.; Halm, U.; Mössner, J.; Berr, F. Decreased immunogenicity of recombinant hepatitis B vaccine in chronic hepatitis C. Hepatology 2000, 31, 230–234. [Google Scholar] [CrossRef]
- Minakari, M.; Tahmasebi, A.; Motlagh, M.H.; Ataei, B.; Yaran, M.; Kalantari, H.; Tavakkoli, H. Efficacy of double dose recombinant hepatitis B vaccination in chronic hepatitis C patients, compared to standard dose vaccination. Int. J. Prev. Med. 2014, 5, 145. [Google Scholar]
- Abd El-Wahab, E.W.; Metwally, M.; Lotfy, N. Effectiveness of hepatitis B vaccination in chronic HCV patients after successful generic direct acting antiviral therapy: Significance of isolated hepatitis B core antibodies. Trop. Med. Int. Health 2021, 26, 882–894. [Google Scholar] [CrossRef]
- Wang, J.M.; Ma, C.J.; Li, G.Y.; Wu, X.Y.; Thayer, P.; Greer, P.; Smith, A.M.; High, K.P.; Moorman, J.P.; Yao, Z.Q. Tim-3 alters the balance of IL-12/IL-23 and drives TH17 cells: Role in hepatitis B vaccine failure during hepatitis C infection. Vaccine 2013, 31, 2238–2245. [Google Scholar] [CrossRef]
- Moorman, J.P.; Zhang, C.L.; Ni, L.; Ma, C.J.; Zhang, Y.; Wu, X.Y.; Thayer, P.; Islam, T.M.; Borthwick, T.; Yao, Z.Q. Impaired hepatitis B vaccine responses during chronic hepatitis C infection: Involvement of the PD-1 pathway in regulating CD4+ T cell responses. Vaccine 2011, 29, 3169–3176. [Google Scholar] [CrossRef]
- Udomkarnjananun, S.; Takkavatakarn, K.; Praditpornsilpa, K.; Nader, C.; Eiam-Ong, S.; Jaber, B.L.; Susantitaphong, P. Hepatitis B virus vaccine immune response and mortality in dialysis patients: A meta-analysis. J. Nephrol. 2020, 33, 343–354. [Google Scholar] [CrossRef]
- Somi, M.H.; Hajipour, B. Improving hepatitis B vaccine efficacy in end-stage renal diseases patients and role of adjuvants. Int. Sch. Res. Not. 2012, 2012, 960413. [Google Scholar] [CrossRef]
- Grzegorzewska, A.E.; Pajzderski, D.; Sowińska, A.; Mostowska, A.; Jagodziński, P.P. IL4R and IL13 polymorphic variants and development of antibodies to surface antigen of hepatitis B virus in hemodialysis patients in response to HBV vaccination or infection. Vaccine 2013, 31, 1766–1770. [Google Scholar] [CrossRef]
- Shabeeb, S.B.; Lilo, K.M.; Ibraheem, S.R. Immune Response Evaluation after Hepatitis B Vaccination among Patients Undergoes Hemodialysis. Biochem. Cell. Arch. 2022, 22, 1913. [Google Scholar]
- Asan, A.; Demirhan, H.; Sorkun, H.Ç.; Özkan, S.; Aydın, M.; Akın, D.; Tatar, B.; Çatak, B.; Şener, A.; Köse, Ş. Factors affecting responsiveness to hepatitis B immunization in dialysis patients. Int. Urol. Nephrol. 2017, 49, 1845–1850. [Google Scholar] [CrossRef]
- Allam Mohamed, E.; Abdel Fattah El-Ballat, M.; Mahmoud El-Sherif, A.; Saeed Al-Shurbagy, M.; Mohammed Abdel Kayioum, M. Interleukin-12 level and its relation to hepatitis b virus vaccine response in hemodialysis patients. Al-Azhar Med. J. 2017, 46, 113–120. [Google Scholar] [CrossRef]
- Feng, Y.; Wang, J.; Shao, Z.; Chen, Z.; Yao, T.; Dong, S.; Wu, Y.; Shi, X.; Shi, J.; Liu, G. Predicting related factors of immunological response to hepatitis B vaccine in hemodialysis patients based on integration of decision tree classification and logistic regression. Hum. Vaccines Immunother. 2021, 17, 3214–3220. [Google Scholar] [CrossRef]
- Asghari, A.; Jafari, F.; Jameshorani, M.; Chiti, H.; Naseri, M.; Ghafourirankouhi, A.; Kooshkaki, O.; Abdshah, A.; Parsamanesh, N. Vitamin D role in hepatitis B: Focus on immune system and genetics mechanism. Heliyon 2022, 8, e11569. [Google Scholar] [CrossRef]
- Grzegorzewska, A.E.; Jodłowska, E.; Mostowska, A.; Sowińska, A.; Jagodziński, P.P. Single nucleotide polymorphisms of vitamin D binding protein, vitamin D receptor and retinoid X receptor alpha genes and response to hepatitis B vaccination in renal replacement therapy patients. Expert Rev. Vaccines 2014, 13, 1395–1403. [Google Scholar] [CrossRef]
- Passanisi, S.; Dipasquale, V.; Romano, C. Vaccinations and immune response in celiac disease. Vaccines 2020, 8, 278. [Google Scholar] [CrossRef]
- Rousseff, T.; Claeys, T.; Vande Vijver, E.; Moes, N.; Vande Velde, S.; Schelstraete, P.; De Bruyne, R.; Van Winckel, M.; Van Biervliet, S. Hepatitis B virus vaccination and revaccination response in children diagnosed with coeliac disease: A multicentre prospective study. Acta Gastro-Enterol. Belg. 2019, 82, 27–30. [Google Scholar]
- Zifman, E.; Zevit, N.; Heshin-Bekenstein, M.; Turner, D.; Shamir, R.; Silbermintz, A. Effect of a gluten free diet on hepatitis B surface antibody concentration in previously immunized pediatric celiac patients. Pediatr. Gastroenterol. Hepatol. Nutr. 2020, 23, 132. [Google Scholar] [CrossRef]
- Zingone, F.; Capone, P.; Tortora, R.; Rispo, A.; Morisco, F.; Caporaso, N.; Imperatore, N.; De Stefano, G.; Iovino, P.; Ciacci, C. Role of gluten intake at the time of hepatitis B virus vaccination in the immune response of celiac patients. Clin. Vaccine Immunol. 2013, 20, 660–662. [Google Scholar] [CrossRef]
- Nemes, E.; Lefler, E.; Szegedi, L.; Kapitány, A.; Kovács, J.B.; Balogh, M.; Szabados, K.; Tumpek, J.; Sipka, S.; Korponay-Szabó, I.R. Gluten intake interferes with the humoral immune response to recombinant hepatitis B vaccine in patients with celiac disease. Pediatrics 2008, 121, e1570–e1576. [Google Scholar] [CrossRef] [PubMed]
- Desombere, I.; Cao, T.; Gijbels, Y.; Leroux-Roels, G. Non-responsiveness to hepatitis B surface antigen vaccines is not caused by defective antigen presentation or a lack of B7 co-stimulation. Clin. Exp. Immunol. 2005, 140, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Ertem, D.; Gonen, I.; Tanidir, C.; Ugras, M.; Yildiz, A.; Pehlivanoglu, E.; Eksioglu-Demiralp, E. The response to hepatitis B vaccine: Does it differ in celiac disease? Eur. J. Gastroenterol. Hepatol. 2010, 22, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.X.; Wang, L.P.; Nie, J.; Jiang, S. Is nonresponsiveness to hepatitis B vaccine due to latent hepatitis B virus infection? J. Infect. Dis. 1992, 165, 777–778. [Google Scholar] [CrossRef] [PubMed]
- Brunetto, M.R.; Giarin, M.M.; Oliveri, F.; Chiaberge, E.; Baldi, M.; Alfarano, A.; Serra, A.; Saracco, G.; Verme, G.; Will, H. Wild-type and e antigen-minus hepatitis B viruses and course of chronic hepatitis. Proc. Natl. Acad. Sci. USA 1991, 88, 4186–4190. [Google Scholar] [CrossRef] [PubMed]
- Paller, A.S.; Mallory, S.B. Acquired forms of immunosuppression. J. Am. Acad. Dermatol. 1991, 24, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Fytili, P.; Bader, C.; Pothakamuri, S.; Schlaphoff, V.; Kuhn, S.; Wedemeyer, H.; Manns, M. Hepatitis B vaccine non-response and occult hepatitis B virus infection. Z. Für Gastroenterol. 2008, 46, K48. [Google Scholar] [CrossRef]
- Colbert, C.; Chavarria, A.; Berkelhammer, C. Fulminant hepatic failure in chronic hepatitis B on withdrawal of corticosteroids, azathioprine and infliximab for Crohn’s disease. Inflamm. Bowel Dis. 2007, 13, 1453–1454. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Chaparro, M.; Esteve, M. prevention and management of hepatitis B and C infection in patients with inflammatory bowel disease. Aliment. Pharmacol. Ther. 2011, 33, 619–633. [Google Scholar] [CrossRef]
- Beaugerie, L.; Rahier, J.-F.; Kirchgesner, J. Predicting, preventing, and managing treatment-related complications in patients with inflammatory bowel diseases. Clin. Gastroenterol. Hepatol. 2020, 18, 1324–1335.e2. [Google Scholar] [CrossRef]
- Manser, C.N.; Maillard, M.H.; Rogler, G.; Schreiner, P.; Rieder, F.; Bühler, S. Vaccination in patients with inflammatory bowel diseases. Digestion 2020, 101, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Baranowska-Nowak, M.; Iwańczak, B.; Szczepanik, M.; Banasiuk, M.; Dembiński, Ł.; Karolewska-Bochenek, K.; Dziekiewicz, M.; Radzikowski, A.; Banaszkiewicz, A. Immune response to hepatitis B vaccination in pediatric patients with inflammatory bowel disease. Cent. Eur. J. Immunol. 2020, 45, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Cossio-Gil, Y.; Martínez-Gómez, X.; Campins-Martí, M.; Rodrigo-Pendás, J.; Borruel-Sainz, N.; Rodríguez-Frías, F.; Casellas-Jordà, F. Immunogenicity of hepatitis B vaccine in patients with inflammatory bowel disease and the benefits of revaccination. J. Gastroenterol. Hepatol. 2015, 30, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Fisman, D.N.; Agrawal, D.; Leder, K. Effect of age on immunologic response to recombinant hepatitis B vaccine: A meta-analysis. Clin. Infect. Dis. 2002, 35, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, J.A.; Habash, N.W.; Ismail, Y.A.; Tremaine, W.J.; Weaver, A.L.; Murray, J.A.; Loftus Jr, E.V.; Absah, I. Effectiveness of Hepatitis B Vaccination for Patients With Inflammatory Bowel and Celiac Disease. Clin. Gastroenterol. Hepatol. 2023, 21, 2901–2907. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.P.; Treat, J.R.; Castelo-Socio, L. Decreased Hepatitis B vaccine response in pediatric patients with atopic dermatitis, psoriasis, and morphea. Vaccine 2017, 35, 4499–4500. [Google Scholar] [CrossRef] [PubMed]
- Elrashidy, H.; Elbahrawy, A.; El-Didamony, G.; Mostafa, M.; George, N.M.; Elwassief, A.; Saeid Mohamed, A.-G.; Elmestikawy, A.; Morsy, M.H.; Hashim, A. Antibody levels against hepatitis B virus after hepatitis B vaccination in Egyptian diabetic children and adolescents. Hum. Vaccines Immunother. 2013, 9, 2002–2006. [Google Scholar] [CrossRef]
- Leonardi, S.; Vitaliti, G.; Garozzo, M.T.; Miraglia del Giudice, M.; Marseglia, G.; La Rosa, M. Hepatitis B vaccination failure in children with diabetes mellitus? The debate continues. Hum. Vaccines Immunother. 2012, 8, 448–452. [Google Scholar] [CrossRef] [PubMed]
- Schillie, S.F.; Spradling, P.R.; Murphy, T.V. Immune response of hepatitis B vaccine among persons with diabetes: A systematic review of the literature. Diabetes Care 2012, 35, 2690. [Google Scholar] [CrossRef]
- Zanoni, G.; Contreas, G.; Valletta, E.; Gabrielli, O.; Mengoli, C.; Veneri, D. Normal or defective immune response to Hepatitis B vaccine in patients with diabetes and celiac disease: An open issue. Hum. Vaccines Immunother. 2015, 11, 58–62. [Google Scholar] [CrossRef]
- Elsharkawy, D.M.A.; El-khaleegy, H.A.E.; Mohamed, S.A.; Mohamed, G.A. Seroprotection Status of Hepatitis B Vaccine in Children with Type 1 Diabetes Mellitus. Int. J. Med. Arts 2021, 3, 1748–1753. [Google Scholar] [CrossRef]
- van den Berg, R.; van Hoogstraten, I.; van Agtmael, M. Non-responsiveness to hepatitis B vaccination in HIV seropositive patients; possible causes and solutions. AIDS Rev. 2009, 11, 157–164. [Google Scholar]
- Bi, X.; Suzuki, Y.; Gatanaga, H.; Oka, S. High frequency and proliferation of CD4+ FOXP3+ Treg in HIV-1-infected patients with low CD4 counts. Eur. J. Immunol. 2009, 39, 301–309. [Google Scholar] [CrossRef]
- Del Mar Del Pozo Balado, M.; Leal, M.; Lagares, G.M.; Mata, R.C.; López-Cortés, L.F.; Viciana, P.; Pacheco, Y.M. Increased regulatory T cell counts in HIV-infected nonresponders to hepatitis B virus vaccine. J. Infect. Dis. 2010, 202, 362–369. [Google Scholar] [CrossRef]
- van der Made, C.I.; Hoischen, A.; Netea, M.G.; van de Veerdonk, F.L. Primary immunodeficiencies in cytosolic pattern-recognition receptor pathways: Toward host-directed treatment strategies. Immunol. Rev. 2020, 297, 247–272. [Google Scholar] [CrossRef]
- Rui-jun, Z.; Lin-zhu, Y.; Jia-xin, W.; Tian-jing, Z.; Yan-di, L.; Ting, W.; Bo, W.; Shu-ying, F.; Yong-liang, F.; Su-ping, W. The effects of TLR3 up-regulation on Th1/Th2 cytokines in CBMCs of infants with non/hypo response to hepatitis B vaccine. Chin. J. Diseas Control. Prev. 2020, 24, 711–715. [Google Scholar]
- Wu, L.; Fu, G.; Xu, X.; Yi, L.; Yao, T.; Wan, H.; Li, J.; Wang, B.; Feng, S.; Feng, Y. Decreased placental TLR3 is associated with hepatitis B virus vaccine responsiveness in infants born to HBsAg-positive mothers. Transl. Pediatr. 2023, 12, 1204. [Google Scholar] [CrossRef]
- Smolnikova, M.V.; Tereshchenko, S.Y. Proteins of the lectin pathway of the complement system activation: Immunobiological functions, genetics and involvement in the pathogenesis of human diseases. Russ. J. Infect. Immun. 2022, 12, 209–221. [Google Scholar] [CrossRef]
- Chong, W.P.; To, Y.F.; Ip, W.K.; Yuen, M.F.; Poon, T.P.; Wong, W.H.; Lai, C.L.; Lau, Y.L. Mannose-binding lectin in chronic hepatitis B virus infection. Hepatology 2005, 42, 1037–1045. [Google Scholar] [CrossRef]
- Tong, H.V.; Toan, N.L.; Song, L.H.; Ouf, E.A.; Bock, C.-T.; Kremsner, P.G.; Kun, J.F.; TP, V. Ficolin-2 levels and FCN2 haplotypes influence hepatitis B infection outcome in Vietnamese patients. PLoS ONE 2011, 6, e28113. [Google Scholar] [CrossRef]
- Xu, H.-d.; Zhao, M.-f.; Wan, T.-h.; Song, G.-z.; He, J.-l.; Chen, Z. Association between Mannose-binding lectin gene polymorphisms and hepatitis B virus infection: A meta-analysis. PLoS ONE 2013, 8, e75371. [Google Scholar] [CrossRef]
- Ruseva, M.; Kolev, M.; Dagnaes-Hansen, F.; Hansen, S.B.; Takahashi, K.; Ezekowitz, A.; Thiel, S.; Jensenius, J.C.; Gadjeva, M. Mannan-binding lectin deficiency modulates the humoral immune response dependent on the genetic environment. Immunology 2009, 127, 279–288. [Google Scholar] [CrossRef]
- Davila, S.; Froeling, F.; Tan, A.; Bonnard, C.; Boland, G.; Snippe, H.; Hibberd, M.; Seielstad, M. New genetic associations detected in a host response study to hepatitis B vaccine. Genes Immun. 2010, 11, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Fourati, S.; Cristescu, R.; Loboda, A.; Talla, A.; Filali, A.; Railkar, R.; Schaeffer, A.K.; Favre, D.; Gagnon, D.; Peretz, Y. Pre-vaccination inflammation and B-cell signalling predict age-related hyporesponse to hepatitis B vaccination. Nat. Commun. 2016, 7, 10369. [Google Scholar] [CrossRef]
- Böcher, W.O.; Herzog-Hauff, S.; Schlaak, J.; zum Büschenfelde, K.-H.M.; Löhr, H.F. Kinetics of Hepatitis B Surface Antigen-Specific Immune Responses in Acute and Chronic Hepatitis B or After Hbs Vaccination: Stimulation of The In Vitro Antibody Response by Interferon Gamma. Hepatology 1999, 29, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T follicular helper cell differentiation, function, and roles in disease. Immunity 2014, 41, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Doedée, A.; Kannegieter, N.; Öztürk, K.; van Loveren, H.; Janssen, R.; Buisman, A.-M. Higher numbers of memory B-cells and Th2-cytokine skewing in high responders to hepatitis B vaccination. Vaccine 2016, 34, 2281–2289. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, W.; Liang, Z.; Wu, X.; Zhu, X.; Li, J.; Li, T.; Wang, L.; Li, H.; Liu, Y. Association between polymorphisms of the cytokine and cytokine receptor genes and immune response to hepatitis B vaccination in a Chinese Han population. J. Med. Virol. 2012, 84, 26–33. [Google Scholar] [CrossRef]
- Kardar, G.; Jeddi-Tehrani, M.; Shokri, F. Diminished Th1 and Th2 cytokine production in healthy adult nonresponders to recombinant hepatitis B vaccine. Scand. J. Immunol. 2002, 55, 311–314. [Google Scholar] [CrossRef]
- Sabry, R.; Mohamed, Z.A.Z.; Abdallah, A.M. Relationship between Th1 and Th2 cytokine serum levels and immune response to Hepatitis B vaccination among Egyptian health care workers. J. Immunoass. Immunochem. 2018, 39, 496–508. [Google Scholar] [CrossRef]
- Honorati, M.C.; Dolzani, P.; Mariani, E.; Piacentini, A.; Lisignoli, G.; Ferrari, C.; Facchini, A. Epitope specificity of Th0/Th2 CD4+ T-lymphocyte clones induced by vaccination with rHBsAg vaccine. Gastroenterology 1997, 112, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Wataya, M.; Sano, T.; Kamikawaji, N.; Tana, T.; Yamamoto, K.; Sasazuki, T. Comparative analysis of HLA restriction and cytokine production in hepatitis B surface antigen-specific T cells from low-and high-antibody responders in vaccinated humans. J. Hum. Genet. 2001, 46, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Velu, V.; Saravanan, S.; Nandakumar, S.; Shankar, E.M.; Vengatesan, A.; Jadhav, S.S.; Kulkarni, P.S.; Thyagarajan, S.P. Relationship between T-lymphocyte cytokine levels and sero-response to hepatitis B vaccines. World J. Gastroenterol. 2008, 14, 3534. [Google Scholar] [CrossRef] [PubMed]
- Jogdand, G.M.; Mohanty, S.; Devadas, S. Regulators of Tfh cell differentiation. Front. Immunol. 2016, 7, 520. [Google Scholar] [CrossRef] [PubMed]
- Sage, P.T.; Sharpe, A.H. T follicular regulatory cells in the regulation of B cell responses. Trends Immunol. 2015, 36, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, N.; Jarrossay, D.; Ho, E.; Avery, D.T.; Ma, C.S.; Yu, D.; Sallusto, F.; Tangye, S.G.; Mackay, C.R. CXCR5 expressing human central memory CD4 T cells and their relevance for humoral immune responses. J. Immunol. 2011, 186, 5556–5568. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.; Schmitt, N.; Bentebibel, S.-E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N. Human blood CXCR5+ CD4+ T cells are counterparts of T follicular cells and contain specific subsets that differentially support antibody secretion. Immunity 2011, 34, 108–121. [Google Scholar] [CrossRef]
- Bentebibel, S.-E.; Khurana, S.; Schmitt, N.; Kurup, P.; Mueller, C.; Obermoser, G.; Palucka, A.K.; Albrecht, R.A.; Garcia-Sastre, A.; Golding, H. ICOS+ PD-1+ CXCR3+ T follicular helper cells contribute to the generation of high-avidity antibodies following influenza vaccination. Sci. Rep. 2016, 6, 26494. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, Y.; Liang, Y.; Yin, M.; Yu, Z.; Zhang, Y.; Huang, L.; Ni, J. MiR-18a and miR-17 are positively correlated with circulating PD-1+ICOS+ follicular helper T cells after hepatitis B vaccination in a chinese population. BMC Immunol. 2018, 19, 25. [Google Scholar] [CrossRef]
- Aghbash, P.S.; Hemmat, N.; Nahand, J.S.; Shamekh, A.; Memar, M.Y.; Babaei, A.; Baghi, H.B. The role of Th17 cells in viral infections. Int. Immunopharmacol. 2021, 91, 107331. [Google Scholar] [CrossRef]
- Mills, K.H. IL-17 and IL-17-producing cells in protection versus pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef]
- Zhang, Y.; Cobleigh, M.A.; Lian, J.Q.; Huang, C.X.; Booth, C.J.; Bai, X.F.; Robek, M.D. A proinflammatory role for interleukin-22 in the immune response to hepatitis B virus. Gastroenterology 2011, 141, 1897–1906. [Google Scholar] [CrossRef]
- Pati, N.T.; Hissar, S.; Agrawal, K.; Rani, R.; Sarin, S.K. Decrease in CD4+ T lymphocyte proliferation responses and enhanced CD150 cell expression in health care workers non-responsive to HBV vaccine. Vaccine 2007, 25, 1848–1855. [Google Scholar] [CrossRef]
- Lu, L.-R.; Wang, X.-Y.; Li, D.-C.; Huang, Z.-L.; Liu, J.; Lin, C.-S. A preliminary study of CD150 expression in responders and non-responders to the Hepatitis B vaccine. Biomed. Res. 2014, 25, 149–152. [Google Scholar]
- Borzooy, Z.; Streinu-Cercel, A.; Mirshafiey, A.; Khamseh, A.; Mahmoudie, M.K.; Navabi, S.S.; Nosrati, M.; Najafi, Z.; Hosseini, M.; Jazayeri, S.M. IL-17 and IL-22 genetic polymorphisms in HBV vaccine non-and low-responders among healthcare workers. Germs 2016, 6, 14. [Google Scholar] [CrossRef][Green Version]
- Hsiao, H.-W.; Liu, W.-H.; Wang, C.-J.; Lo, Y.-H.; Wu, Y.-H.; Jiang, S.-T.; Lai, M.-Z. Deltex1 is a target of the transcription factor NFAT that promotes T cell anergy. Immunity 2009, 31, 72–83. [Google Scholar] [CrossRef]
- Xie, B.; Zhang, P.; Liu, M.; Zeng, W.; Yang, J.; Liu, H. Deltex1 Polymorphisms Are Associated with Hepatitis B Vaccination Non-Response in Southwest China. PLoS ONE 2016, 11, e0149199. [Google Scholar] [CrossRef]
- Zhang, X.; Sun, X.; Li, M.; Zhou, Z.; Gao, Y. A promoter polymorphism (rs3806798) of interleukin-15 gene is associated with chronic hepatitis B virus infection in the C hinese H an population. Int. J. Immunogenet. 2014, 41, 298–305. [Google Scholar] [CrossRef]
- Chekol Abebe, E.; Asmamaw Dejenie, T.; Mengie Ayele, T.; Dagnew Baye, N.; Agegnehu Teshome, A.; Tilahun Muche, Z. The role of regulatory B cells in health and diseases: A systemic review. J. Inflamm. Res. 2021, 2021, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Rosser, E.C.; Blair, P.A.; Mauri, C. Cellular targets of regulatory B cell-mediated suppression. Mol. Immunol. 2014, 62, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Rui, K.; Wang, S.; Lu, L. Regulatory B cells in autoimmune diseases. Cell. Mol. Immunol. 2013, 10, 122–132. [Google Scholar] [CrossRef]
- Bolther, M.; Andersen, K.L.D.; Tolstrup, M.; Visvanathan, K.; Woolley, I.; Skinner, N.; Millen, R.; Warner, N.; Østergaard, L.; Jensen-Fangel, S. Levels of regulatory B cells do not predict serological responses to hepatitis B vaccine. Hum. Vaccines Immunother. 2018, 14, 1483–1488. [Google Scholar] [CrossRef]
- Garner-Spitzer, E.; Wagner, A.; Paulke-Korinek, M.; Kollaritsch, H.; Heinz, F.X.; Redlberger-Fritz, M.; Stiasny, K.; Fischer, G.F.; Kundi, M.; Wiedermann, U. Tick-borne encephalitis (TBE) and hepatitis B nonresponders feature different immunologic mechanisms in response to TBE and influenza vaccination with involvement of regulatory T and B cells and IL-10. J. Immunol. 2013, 191, 2426–2436. [Google Scholar] [CrossRef]
- Körber, N.; Pohl, L.; Weinberger, B.; Grubeck-Loebenstein, B.; Wawer, A.; Knolle, P.A.; Roggendorf, H.; Protzer, U.; Bauer, T. Hepatitis B Vaccine Non-Responders Show Higher Frequencies of CD24highCD38high Regulatory B Cells and Lower Levels of IL-10 Expression Compared to Responders. Front. Immunol. 2021, 12, 713351. [Google Scholar] [CrossRef]
- Posteraro, B.; Pastorino, R.; Di Giannantonio, P.; Ianuale, C.; Amore, R.; Ricciardi, W.; Boccia, S. The link between genetic variation and variability in vaccine responses: Systematic review and meta-analyses. Vaccine 2014, 32, 1661–1669. [Google Scholar] [CrossRef]
- Cardell, K.; Lindblom, B.; Frydén, A. Hepatitis B Vaccination in Relatives to Known Non-Responders: A Family Study; Stockholm University: Stockholm, Sweden, 2009. [Google Scholar]
- Qiu, S.; He, P.; Fang, X.; Tong, H.; Lv, J.; Liu, J.; Zhang, L.; Zhai, X.; Wang, L.; Hu, Z. Significant transcriptome and cytokine changes in hepatitis B vaccine non-responders revealed by genome-wide comparative analysis. Hum. Vaccines Immunother. 2018, 14, 1763–1772. [Google Scholar] [CrossRef]
- Xion, Y.; Chen, S.; Chen, R.; Lin, W.; Ni, J. Association between microRNA polymorphisms and humoral immunity to hepatitis B vaccine. Hum. Vaccines Immunother. 2013, 9, 1673–1678. [Google Scholar] [CrossRef][Green Version]
- Bartholomeus, E.; De Neuter, N.; Meysman, P.; Suls, A.; Keersmaekers, N.; Elias, G.; Jansens, H.; Hens, N.; Smits, E.; Van Tendeloo, V. Transcriptome profiling in blood before and after hepatitis B vaccination shows significant differences in gene expression between responders and non-responders. Vaccine 2018, 36, 6282–6289. [Google Scholar] [CrossRef]
- Loureiro, J.; Ploegh, H.L. Antigen presentation and the ubiquitin-proteasome system in host–pathogen interactions. Adv. Immunol. 2006, 92, 225–305. [Google Scholar] [PubMed]
- Pishesha, N.; Harmand, T.J.; Ploegh, H.L. A guide to antigen processing and presentation. Nat. Rev. Immunol. 2022, 22, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.; Kapelus, D.; Singh, P.; Groome, M.; de Assis Rosa, D. ABO blood types, but not Secretor or Lewis blood types, influence strength of antibody response to Hepatitis B vaccine in Black South African children. Vaccine 2023, 41, 3617–3626. [Google Scholar] [CrossRef]
- Alper, C.A.; Kruskall, M.S.; Marcus-Bagley, D.; Craven, D.E.; Katz, A.J.; Brink, S.J.; Dienstag, J.L.; Awdeh, Z.; Yunis, E.J. Genetic Prediction of Nonresponse to Hepatitis B Vaccine. New Engl. J. Med. 1989, 321, 708–712. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, C.; Li, P.; Sun, T.; Wang, J.; Zhang, S.; Ma, Q.; Ma, F.; Shi, W.; Shi, M. Single-cell RNA sequencing reveals the transcriptomic characteristics of peripheral blood mononuclear cells in hepatitis B vaccine non-responders. Front. Immunol. 2023, 14, 1091237. [Google Scholar] [CrossRef]
- Lin, H.H.; Liao, H.W.; Lin, S.K.; Wang, L.Y. HLA and response to booster hepatitis B vaccination in anti-HBs-seronegative adolescents who had received primary infantile vaccination. Vaccine 2008, 26, 3414–3420. [Google Scholar] [CrossRef]
- Wang, L.-Y.; Chen, C.-F.; Wu, T.-W.; Lai, S.-K.; Chu, C.-C.; Lin, H.H. Response to hepatitis B vaccination is co-determined by HLA-DPA1 and-DPB1. Vaccine 2019, 37, 6435–6440. [Google Scholar] [CrossRef]
- Li, Z.-K.; Nie, J.-J.; Li, J.; Zhuang, H. The effect of HLA on immunological response to hepatitis B vaccine in healthy people: A meta-analysis. Vaccine 2013, 31, 4355–4361. [Google Scholar] [CrossRef]
- Mineta, M.; Tanimura, M.; Tana, T.; Yssel, H.; Kashiwagi, S.; Sasazuki, T. Contribution of HLA class I and class II alleles to the regulation of antibody production to hepatitis B surface antigen in humans. Int. Immunol. 1996, 8, 525–531. [Google Scholar] [CrossRef]
- Wang, C.; Tang, J.; Song, W.; Lobashevsky, E.; Wilson, C.M.; Kaslow, R.A. HLA and cytokine gene polymorphisms are independently associated with responses to hepatitis B vaccination. Hepatology 2004, 39, 978–988. [Google Scholar] [CrossRef]
- Desombere, I.; Willems, A.; Leroux-Roels, G. Response to hepatitis B vaccine: Multiple HLA genes are involved. Tissue Antigens 1998, 51, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Amirzargar, A.A.; Mohseni, N.; Shokrgozar, M.A.; Arjang, Z.; Ahmadi, N.; Yousefi Behzadi, M.; Amanzadeh, A.; Shokri, F. HLA-DRB1, DQA1 and DQB1 Alleles and Hap-lotypes Frequencies in Iranian Healthy Adult Responders and Non-Responders to Recombi-nant Hepatitis B Vaccine. Iran. J. Immunol. 2008, 5, 92–99. [Google Scholar] [PubMed]
- Yoon, J.H.; Shin, S.; won In, J.; Chang, J.Y.; Song, E.Y.; Roh, E.Y. Association of HLA alleles with the responsiveness to hepatitis B virus vaccination in Korean infants. Vaccine 2014, 32, 5638–5644. [Google Scholar] [CrossRef] [PubMed]
- Höhler, T.; Reuss, E.; Evers, N.; Dietrich, E.; Rittner, C.; Freitag, C.M.; Vollmar, J.; Schneider, P.M.; Fimmers, R. Differential genetic determination of immune responsiveness to hepatitis B surface antigen and to hepatitis A virus: A vaccination study in twins. Lancet 2002, 360, 991–995. [Google Scholar] [CrossRef] [PubMed]
- Jilin, Z.; Yuguo, S. The effect of HLA-DRB1 alleles in immune response to hepatitis B vaccine. In Proceedings of the 2011 International Conference on Human Health and Biomedical Engineering, Jilin, China, 19–22 August 2011; pp. 554–557. [Google Scholar]
- Milich, D.R.; Leroux-Roels, G.G. Immunogenetics of the response to HBsAg vaccination. Autoimmun. Rev. 2003, 2, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Sakai, A.; Noguchi, E.; Fukushima, T.; Tagawa, M.; Iwabuchi, A.; Kita, M.; Kakisaka, K.; Miyasaka, A.; Takikawa, Y.; Sumazaki, R. Identification of amino acids in antigen-binding site of class II HLA proteins independently associated with hepatitis B vaccine response. Vaccine 2017, 35, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Höhler, T.; Stradmann-Bellinghausen, B.; Starke, R.; Sänger, R.; Victor, A.; Rittner, C.; Schneider, P.M. C4A deficiency and nonresponse to hepatitis B vaccination. J. Hepatol. 2002, 37, 387–392. [Google Scholar] [CrossRef] [PubMed]
- De Silvestri, A.; Pasi, A.; Martinetti, M.; Belloni, C.; Tinelli, C.; Rondini, G.; Salvaneschi, L.; Cuccia, M. Family study of non-responsiveness to hepatitis B vaccine confirms the importance of HLA class III C4A locus. Genes Immun. 2001, 2, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhu, D.; Bi, Y.; Wang, Y.; Hu, Y.; Zhou, Y.-H. Minimal association of alleles of human leukocyte antigen class II gene and long-term antibody response to hepatitis B vaccine vaccinated during infancy. Vaccine 2017, 35, 2457–2462. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.W.; Chen, C.F.; Lai, S.K.; Lin, H.H.; Chu, C.C.; Wang, L.Y. SNP rs7770370 in HLA-DPB 1 loci as a major genetic determinant of response to booster hepatitis B vaccination: Results of a genome-wide association study. J. Gastroenterol. Hepatol. 2015, 30, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Uno, N.; Sato, S.; Mori, S.; Sasaki, D.; Kaku, N.; Kosai, K.; Morinaga, Y.; Hasegawa, H.; Yanagihara, K. Strong influence of human leukocyte antigen-DP variants on response to hepatitis B vaccine in a Japanese population. Vaccine 2017, 35, 5662–5665. [Google Scholar] [CrossRef]
- Yu, L.; Cheng, Y.-j.; Cheng, M.-l.; Yao, Y.-m.; Zhang, Q.; Zhao, X.-k.; Liu, H.-j.; Hu, Y.-x.; Mu, M.; Wang, B. Quantitative assessment of common genetic variations in HLA-DP with hepatitis B virus infection, clearance and hepatocellular carcinoma development. Sci. Rep. 2015, 5, 14933. [Google Scholar] [CrossRef][Green Version]
- Pan, L.; Zhang, L.; Zhang, W.; Wu, X.; Li, Y.; Yan, B.; Zhu, X.; Liu, X.; Yang, C.; Xu, J. A genome-wide association study identifies polymorphisms in the HLA-DR region associated with non-response to hepatitis B vaccination in Chinese Han populations. Hum. Mol. Genet. 2014, 23, 2210–2219. [Google Scholar] [CrossRef]
- Roh, E.Y.; Yoon, J.H.; In, J.W.; Lee, N.; Shin, S.; Song, E.Y. Association of HLA-DP variants with the responsiveness to Hepatitis B virus vaccination in Korean Infants. Vaccine 2016, 34, 2602–2607. [Google Scholar] [CrossRef]
- Duan, Z.; Chen, X.; Liang, Z.; Zeng, Y.; Zhu, F.; Long, L.; McCrae, M.A.; Zhuang, H.; Shen, T.; Lu, F. Genetic polymorphisms of CXCR5 and CXCL13 are associated with non-responsiveness to the hepatitis B vaccine. Vaccine 2014, 32, 5316–5322. [Google Scholar] [CrossRef]
- Zhuo, Y.; Yang, Y.; Zhang, M.; Xu, Y.; Chen, Z.; Mu, L.; Tang, X.; Zhong, Z.; Chen, J.; Zhou, L. Single nucleotide polymorphisms in IFN-γ signaling pathway associated with risk of hepatitis B virus infection in Chinese children. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 8121659. [Google Scholar] [CrossRef]
- Amirpour-Rostami, S.; Kazemi Arababadi, M. IL-18 and IL-1β gene polymorphisms: The plausible risk factors for chronic hepatitis B. Viral Immunol. 2019, 32, 208–213. [Google Scholar] [CrossRef]
- Chen, J.; Liang, Z.; Lu, F.; Fang, X.; Liu, S.; Zeng, Y.; Zhu, F.; Chen, X.; Shen, T.; Li, J. Toll-like receptors and cytokines/cytokine receptors polymorphisms associate with non-response to hepatitis B vaccine. Vaccine 2011, 29, 706–711. [Google Scholar] [CrossRef]
- Çiftdoǧan, D.Y.; Onay, H.; Tosun, S.; Özdemir, T.R.; Özkinay, F.; Vardar, F. IFNG and IFNGR1 gene polymorphisms in children with nonresponse to the hepatitis B vaccine. Future Virol. 2014, 9, 123–130. [Google Scholar] [CrossRef]
- Burton, A.R.; Maini, M.K. Human antiviral B cell responses: Emerging lessons from hepatitis B and COVID-19. Immunol. Rev. 2021, 299, 108–117. [Google Scholar] [CrossRef]
- Pondé, R.A.d.A. Expression and detection of anti-HBs antibodies after hepatitis B virus infection or vaccination in the context of protective immunity. Arch. Virol. 2019, 164, 2645–2658. [Google Scholar] [CrossRef]
- Khodadadi, L.; Cheng, Q.; Radbruch, A.; Hiepe, F. The maintenance of memory plasma cells. Front. Immunol. 2019, 10, 721. [Google Scholar] [CrossRef]
- Van Damme, P.; Dionne, M.; Leroux-Roels, G.; Van Der Meeren, O.; Di Paolo, E.; Salaun, B.; Surya Kiran, P.; Folschweiller, N. Persistence of HB sAg-specific antibodies and immune memory two to three decades after hepatitis B vaccination in adults. J. Viral Hepat. 2019, 26, 1066–1075. [Google Scholar] [CrossRef] [PubMed]
- Rosado, M.M.; Scarsella, M.; Pandolfi, E.; Cascioli, S.; Giorda, E.; Chionne, P.; Madonne, E.; Gesualdo, F.; Romano, M.; Ausiello, C.M. Switched memory B cells maintain specific memory independently of serum antibodies: The hepatitis B example. Eur. J. Immunol. 2011, 41, 1800–1808. [Google Scholar] [CrossRef]
- Samandari, T.; Fiore, A.E.; Negus, S.; Williams, J.L.; Kuhnert, W.; McMahon, B.J.; Bell, B.P. Differences in response to a hepatitis B vaccine booster dose among Alaskan children and adolescents vaccinated during infancy. Pediatrics 2007, 120, e373–e381. [Google Scholar] [CrossRef] [PubMed]
- Coursaget, P.; Leboulleux, D.; Soumare, M.; le Cann, P.; Yvonnet, B.; Chiron, J.-P.; Coll-Seck, A.-M.; Diop-Mar, I. Twelve-year follow-up study of hepatitis B immunization of Senegalese infants. J. Hepatol. 1994, 21, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Zaffina, S.; Marcellini, V.; Santoro, A.P.; Scarsella, M.; Camisa, V.; Vinci, M.R.; Musolino, A.M.; Nicolosi, L.; Rosado, M.M.; Carsetti, R. Repeated vaccinations do not improve specific immune defenses against Hepatitis B in non-responder health care workers. Vaccine 2014, 32, 6902–6910. [Google Scholar] [CrossRef] [PubMed]
- Valats, J.; Tuaillon, E.; Funakoshi, N.; Hoa, D.; Brabet, M.; Bollore, K.; Ducos, J.; Vendrell, J.; Blanc, P. Investigation of memory B cell responses to hepatitis B surface antigen in health care workers considered as non-responders to vaccination. Vaccine 2010, 28, 6411–6416. [Google Scholar] [CrossRef]
- Karimi, S.; Mehdipour, F.; Sarvari, J.; Ataollahi, M.R.; Ramezani, A.; Meri, S.; Kalantar, K. Investigation of the frequencies of various B cell populations in non-responder healthcare workers in comparison with responders to hepatitis B virus vaccination. Trans. R. Soc. Trop. Med. Hyg. 2023, 117, 628–636. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, J.; Lu, J.; Yan, B.; Song, L.; Li, L.; Cui, F.; Zhang, G.; Wang, F.; Liang, X. Antibody response to revaccination among adult non-responders to primary Hepatitis B vaccination in China. Hum. Vaccines Immunother. 2015, 11, 2716–2722. [Google Scholar] [CrossRef]
- Raven, S.F.; Hoebe, C.J.; Vossen, A.C.; Visser, L.G.; Hautvast, J.L.; Roukens, A.H.; van Steenbergen, J.E. Serological response to three alternative series of hepatitis B revaccination (Fendrix, Twinrix, and HBVaxPro-40) in healthy non-responders: A multicentre, open-label, randomised, controlled, superiority trial. Lancet Infect. Dis. 2020, 20, 92–101. [Google Scholar] [CrossRef]
- Hoebe, C.J.; Vermeiren, A.P.; Dukers-Muijrers, N.H. Revaccination with Fendrix® or HBVaxPro® results in better response rates than does revaccination with three doses of Engerix-B® in previous non-responders. Vaccine 2012, 30, 6734–6737. [Google Scholar] [CrossRef]
- Grazzini, M.; Arcangeli, G.; Mucci, N.; Bonanni, P.; Bini, C.; Bechini, A.; Boccalini, S.; Tiscione, E.; Paolini, D. High chance to overcome the non-responder status to hepatitis B vaccine after a further full vaccination course: Results from the extended study on healthcare students and workers in Florence, Italy. Hum. Vaccines Immunother. 2020, 16, 949–954. [Google Scholar] [CrossRef]
- Li, J.; Meng, F.; Zheng, J.; Liang, Q.; Li, H.; Li, J.; Zhang, L.; Gan, J.; Zhu, F. Immune persistence induced by three doses of 60 μg hepatitis B vaccine in non-responders following standard primary vaccination in Chinese adults. Hum. Vaccines Immunother. 2021, 17, 2762–2767. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.-X.; Zeng, Y.; Song, X.-F.; Zhang, Y.-J.; Xu, K.; Liang, Z.-L.; Zhu, F.-C. Immune response to hepatitis B vaccine with high antigen content in non-responders after standard primary vaccination in Chinese adults. Vaccine 2014, 32, 3706–3712. [Google Scholar] [CrossRef] [PubMed]
- Shooshtari, M.H.S.; Makvandi, M.; Rasti, M.; Neisi, N.; Rastegarvand, N.; Pouremamali, A.; Haj, M.S.; Ghaedi, F. Evaluation of hepatitis B surface antibody and specific gamma interferon response in health care workers after vaccination. Jundishapur J. Microbiol. 2015, 8, e13801. [Google Scholar] [CrossRef] [PubMed]
- Saraceni, C.; Birk, J. A review of hepatitis B virus and hepatitis C virus immunopathogenesis. J. Clin. Transl. Hepatol. 2021, 9, 409. [Google Scholar] [CrossRef] [PubMed]
- Fabrizi, F.; Cerutti, R.; Dixit, V.; Ridruejo, E. Hepatitis B virus vaccine and chronic kidney disease. The advances. Nefrologia 2021, 41, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Koc, Ö.M.; Damoiseaux, J.; van Loo, I.H.; Masquillier, H.I.; Lashof, A.M.O. Case report of delayed seroprotection rather than non-response after primary three-dose hepatitis B vaccination. Vaccine 2020, 38, 112–114. [Google Scholar] [CrossRef] [PubMed]
- Madhavan, A.; Palappallil, D.S.; Balakrishnapanicker, J.; Asokan, A. Immune response to hepatitis B vaccine: An evaluation. Perspect. Clin. Res. 2021, 12, 209. [Google Scholar] [PubMed]
- Hanif, F.; Mehmood, N.; Majid, Z.; Luck, N.; Laeeq, S.; Tasneem, A.; Ul Haque, M. Successful response of intradermal hepatitis B vaccine in nonresponders of intramuscular hepatitis B vaccine in general and hemodialysis population. Saudi J. Gastroenterol. 2020, 26, 306–311. [Google Scholar] [CrossRef]
- Leonardi, S.; Praticò, A.D.; Lionetti, E.; Spina, M.; Vitaliti, G.; La Rosa, M. Intramuscular vs intradermal route for hepatitis B booster vaccine in celiac children. World J. Gastroenterol. 2012, 18, 5729. [Google Scholar] [CrossRef]
- Dhillon, S.; Moore, C.; Li, S.; Aziz, A.; Kakar, A.; Dosanjh, A.; Beesla, A.; Murphy, L.; Van Thiel, D. Efficacy of high-dose intra-dermal hepatitis B virus vaccine in previous vaccination non-responders with chronic liver disease. Dig. Dis. Sci. 2012, 57, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Filippelli, M.; Lionetti, E.; Gennaro, A.; Lanzafame, A.; Arrigo, T.; Salpietro, C.; La Rosa, M.; Leonardi, S. Hepatitis B vaccine by intradermal route in non responder patients: An update. World J. Gastroenterol. 2014, 20, 10383–10394. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, P.; Tchitchek, N.; Joly, C.; Rodriguez Pozo, A.; Stimmer, L.; Langlois, S.; Hocini, H.; Gosse, L.; Pejoski, D.; Cosma, A. Vaccine inoculation route modulates early immunity and consequently antigen-specific immune response. Front. Immunol. 2021, 12, 645210. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Chu, L.Y.; Burton, S.A.; Hansen, K.J.; Panyam, J. Intradermal delivery of vaccine nanoparticles using hollow microneedle array generates enhanced and balanced immune response. J. Control. Release 2019, 294, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Menon, I.; Bagwe, P.; Gomes, K.B.; Bajaj, L.; Gala, R.; Uddin, M.N.; D’souza, M.J.; Zughaier, S.M. Microneedles: A new generation vaccine delivery system. Micromachines 2021, 12, 435. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Aggarwal, A.; Aggarwal, R. Hepatitis B vaccine: Using skin when muscle does not work. J. Gastroenterol. Hepatol. 2016, 31, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Erstad, B.L.; Barletta, J.F. Implications of obesity for drug administration and absorption from subcutaneous and intramuscular injections: A primer. Am. J. Health-Syst. Pharm. 2022, 79, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z. Effect of Aluminum Adjuvant on the Structure of Recombinant Hepatitis B Core Antigen. Front. Soc. Sci. Technol. 2020, 2, 27–38. [Google Scholar]
- Wang, Z.; Li, S.; Shan, P.; Wei, D.; Hao, S.; Zhang, Z.; Xu, J. Improved aluminum adjuvants eliciting stronger immune response when mixed with hepatitis B virus surface antigens. ACS Omega 2022, 7, 34528–34537. [Google Scholar] [CrossRef]
- Tajiri, K.; Shimizu, Y. Unsolved problems and future perspectives of hepatitis B virus vaccination. World J. Gastroenterol. 2015, 21, 7074. [Google Scholar] [CrossRef]
- Tanwar, S.; Thursz, M. Granulocyte colony-stimulating factor as a novel adjunct to improve hepatitis B vaccination. World J. Hepatol. 2010, 2, 136. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Mandal, S.; Thakur, A.; Kapoor, D.; Anuradha, S.; Prakash, A.; Kohli, R.; Agarwal, S. Efficacy of GM-CSF as an adjuvant to hepatitis B vaccination in patients with chronic renal failure—Results of a prospective, randomized trial. Ren. Fail. 2003, 25, 255–266. [Google Scholar] [CrossRef] [PubMed]
- das Graças Sasaki, M.; Foccacia, R.; de Messias-Reason, I.J. Efficacy of granulocyte-macrophage colony-stimulating factor (GM-CSF) as a vaccine adjuvant for hepatitis B virus in patients with HIV infection. Vaccine 2003, 21, 4545–4549. [Google Scholar]
- Lin, C.; Zhu, J.; Zheng, Y.; Chen, Y.; Wu, Z.; Chong, Y.; Gao, Z. Effect of GM-CSF in combination with hepatitis B vaccine on revacination of healthy adult non-responders. J. Infect. 2010, 60, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.Y.; Lin, X.Z.; Pan, W.Y.; Wu, P.Y.; Chang, C.M.; Lin, T.Y.; Shen, H.H.; Tao, M.H. Hydrogel-delivered GM-CSF overcomes nonresponsiveness to hepatitis B vaccine through the recruitment and activation of dendritic cells. J. Immunol. 2010, 185, 5468–5475. [Google Scholar] [CrossRef]
- Brunel, F.; Darbouret, A.; Ronco, J. Cationic lipid DC-Chol induces an improved and balanced immunity able to overcome the unresponsiveness to the hepatitis B vaccine. Vaccine 1999, 17, 2192–2203. [Google Scholar] [CrossRef]
- Shouval, D.; Roggendorf, H.; Roggendorf, M. Enhanced immune response to hepatitis B vaccination through immunization with a Pre-S1/Pre-S2/S vaccine. Med. Microbiol. Immunol. 2015, 204, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, A.; Ludwig, C.; Jochum, C.; Fiedler, M.; Heinemann, F.M.; Shouval, D.; Roggendorf, M.; Roggendorf, H.; Lindemann, M. Induction of a robust T- and B-cell immune response in non- and low-responders to conventional vaccination against hepatitis B by using a third generation PreS/S vaccine. Vaccine 2014, 32, 5077–5082. [Google Scholar] [CrossRef] [PubMed]
- Rendi-Wagner, P.; Shouval, D.; Genton, B.; Lurie, Y.; Rümke, H.; Boland, G.; Cerny, A.; Heim, M.; Bach, D.; Schroeder, M. Comparative immunogenicity of a PreS/S hepatitis B vaccine in non-and low responders to conventional vaccine. Vaccine 2006, 24, 2781–2789. [Google Scholar] [CrossRef] [PubMed]
- Mizrahi, M.; Lalazar, G.; Ben Ya’acov, A.; Livovsky, D.M.; Horowitz, Y.; Zolotarov, L.; Adler, R.; Shouval, D.; Ilan, Y. Beta-glycoglycosphingolipid-induced augmentation of the anti-HBV immune response is associated with altered CD8 and NKT lymphocyte distribution: A novel adjuvant for HBV vaccination. Vaccine 2008, 26, 2589–2595. [Google Scholar] [CrossRef]
- Lindemann, M.; Zaslavskaya, M.; Fiedler, M.; Wilde, B.; Heinemann, F.; Heinold, A.; Horn, P.; Witzke, O. Humoral and Cellular Responses to a Single Dose of Fendrix in Renal Transplant Recipients with Non-response to Previous Hepatitis B Vaccination. Scand. J. Immunol. 2017, 85, 51–57. [Google Scholar] [CrossRef]
- Babu, T.M.; Kotton, C.N. Immunizations in chronic kidney disease and kidney transplantation. Curr. Treat. Options Infect. Dis. 2021, 13, 47–65. [Google Scholar] [CrossRef]
- Bauer, T.; Günther, M.; Bienzle, U.; Neuhaus, R.; Jilg, W. Vaccination against hepatitis B in liver transplant recipients: Pilot analysis of cellular immune response shows evidence of HBsAg-specific regulatory T cells. Liver Transplant. 2007, 13, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Michaelides, K.; Prasanna, M.; Badhan, R.; Mohammed, A.-U.-R.; Walters, A.; Howard, M.K.; Dulal, P.; Al-Khattawi, A. Single administration vaccines: Delivery challenges, in vivo performance, and translational considerations. Expert Rev. Vaccines 2023, 22, 579–595. [Google Scholar] [CrossRef] [PubMed]
- Cardell, K.; Akerlind, B.; Sällberg, M.; Frydén, A. Excellent response rate to a double dose of the combined hepatitis A and B vaccine in previous nonresponders to hepatitis B vaccine. J. Infect. Dis. 2008, 198, 299–304. [Google Scholar] [CrossRef]
- Shahidi, S.; Ghareghani, N.N.; Mortazavi, M.; Sadeghi, S.; Adeli, R. The evaluation of Tetanus-diphtheria (Td) vaccine impacts on immune response to hepatitis B (HB) vaccine in non-responder dialysis patients. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2011, 16, 598. [Google Scholar]
- Haghighat, A.; Moafi, M.; Sharifian, J.; Salehi, H.; Taleban, R.; Kalbasi, N.; Salehi, M.; Salehi, M.M.; Salehi, M. Effect of tetanus-diphtheria vaccine on immune response to Hepatitis b vaccine in low-responder individuals. Int. J. Prev. Med. 2016, 7, 94. [Google Scholar]
- Sönmez, E.; Sönmez, A.S.; Bayindir, Y.; Coskun, D.; Aritürk, S. Antihepatitis B response to hepatitis B vaccine administered simultaneously with tetanus toxoid in nonresponder individuals. Vaccine 2002, 21, 243–246. [Google Scholar] [CrossRef]
- Fabrizi, F.; Ganeshan, S.; Dixit, V.; Martin, P. Meta-analysis: The adjuvant role of granulocyte macrophage-colony stimulating factor on immunological response to hepatitis B virus vaccine in end-stage renal disease. Aliment. Pharmacol. Ther. 2006, 24, 789–796. [Google Scholar] [CrossRef]
- Saade, F.; Honda-Okubo, Y.; Trec, S.; Petrovsky, N. A novel hepatitis B vaccine containing Advax™, a polysaccharide adjuvant derived from delta inulin, induces robust humoral and cellular immunity with minimal reactogenicity in preclinical testing. Vaccine 2013, 31, 1999–2007. [Google Scholar] [CrossRef]
- Ramezani, A.; Eslamifar, A.; Banifazl, M.; Ahmadi, F.; Maziar, S.; Razeghi, E.; Kalantar, E.; Amirkhani, A.; Aghakhani, A. Efficacy and long-term immunogenicity of hepatitis B vaccine in haemodialysis patients. Int. J. Clin. Pract. 2009, 63, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Fabrizi, F.; Dixit, V.; Messa, P.; Martin, P. Intradermal vs intramuscular vaccine against hepatitis B infection in dialysis patients: A meta-analysis of randomized trials. J. Viral Hepat. 2011, 18, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Christodoulou, M.; Aspray, T.J.; Schoenmakers, I. Vitamin D supplementation for patients with chronic kidney disease: A systematic review and meta-analyses of trials investigating the response to supplementation and an overview of guidelines. Calcif. Tissue Int. 2021, 109, 157–178. [Google Scholar] [CrossRef] [PubMed]
- Vingerhoets, J.; Vanham, G.; Kestens, L.; Penne, G.; Leroux-Roels, G.; Gigase, P. Deficient T-cell responses in non-responders to hepatitis B vaccination: Absence of TH1 cytokine production. Immunol. Lett. 1994, 39, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Jarrosson, L.; Kolopp-Sarda, M.; Aguilar, P.; Bene, M.; Lepori, M.; Vignaud, M.; Faure, G.; Kohler, C. Most humoral non-responders to hepatitis B vaccines develop HBV-specific cellular immune responses. Vaccine 2004, 22, 3789–3796. [Google Scholar] [CrossRef]
- Hsu, Y.-C.; Huang, D.Q.; Nguyen, M.H. Global burden of hepatitis B virus: Current status, missed opportunities and a call for action. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 524–537. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).