

Lecanicillium psalliotae (Hypocreales: Cordycipitaceae) Exerts Ovicidal and Larvicidal Effects against the Sheep Blood-Feeding Nematode Haemonchus contortus through Its Liquid Culture Filtrates
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location
2.2. Haemonchus contortus Eggs and Infective Larvae
2.3. Fungal Isolation
2.4. Fungal Morphological Identification
2.5. Fungal Molecular Identification
2.6. Fungal Culture in Liquid Media
2.7. Fungal Liquid Culture Filtrates
2.8. Assessment of the In Vitro Nematocidal Activity of Lecanicillium psalliotae against Haemonchus contortus
Assessing the Larvicidal Activity of Lecanicillium psalliotae Liquid Culture Filtrates against Haemonchus contortus
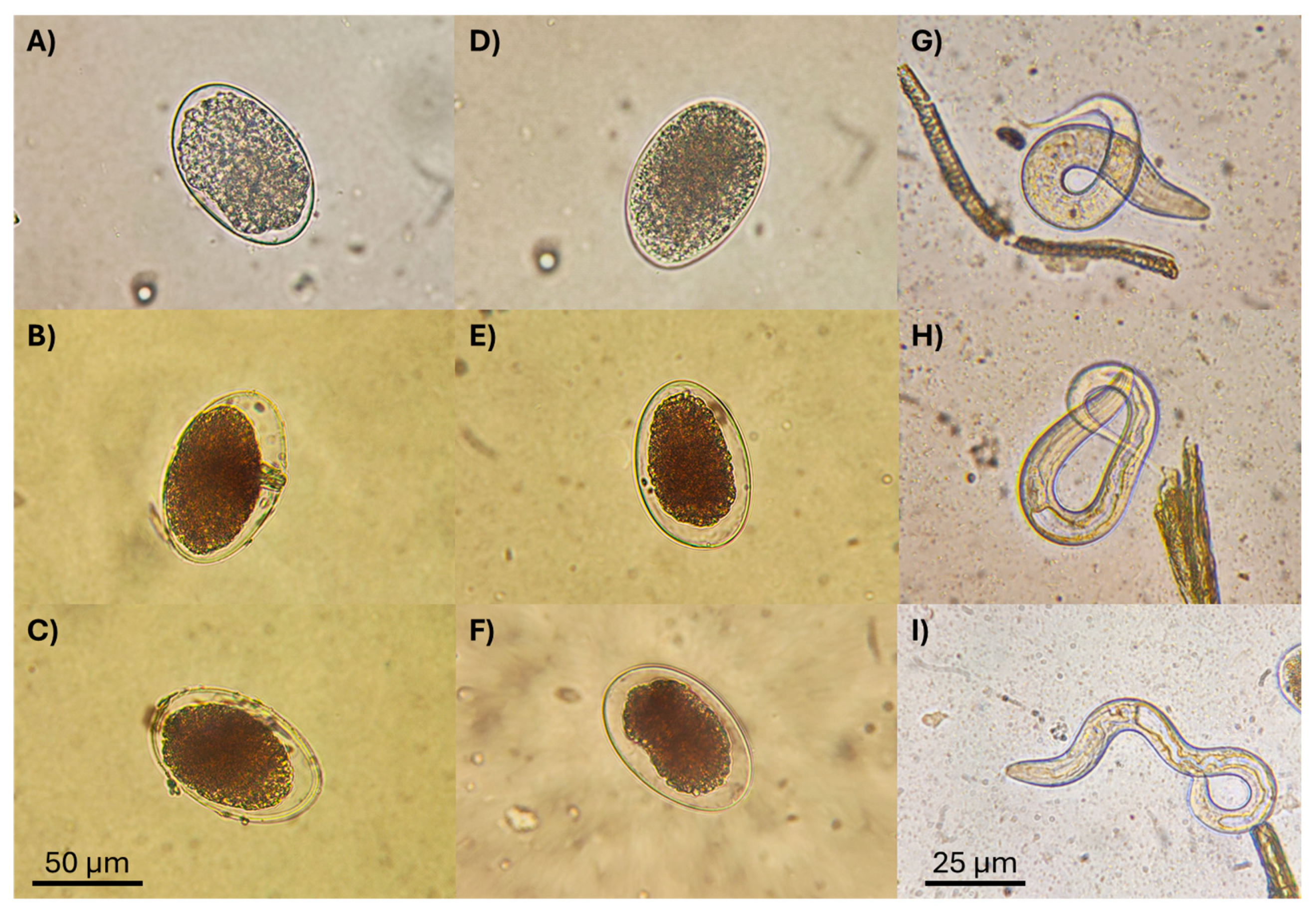
2.9. Microphotographic Record
2.10. Myco-Chemical Profile
2.11. Statistical Analysis
3. Results
3.1. Fungal Morphological Taxonomy
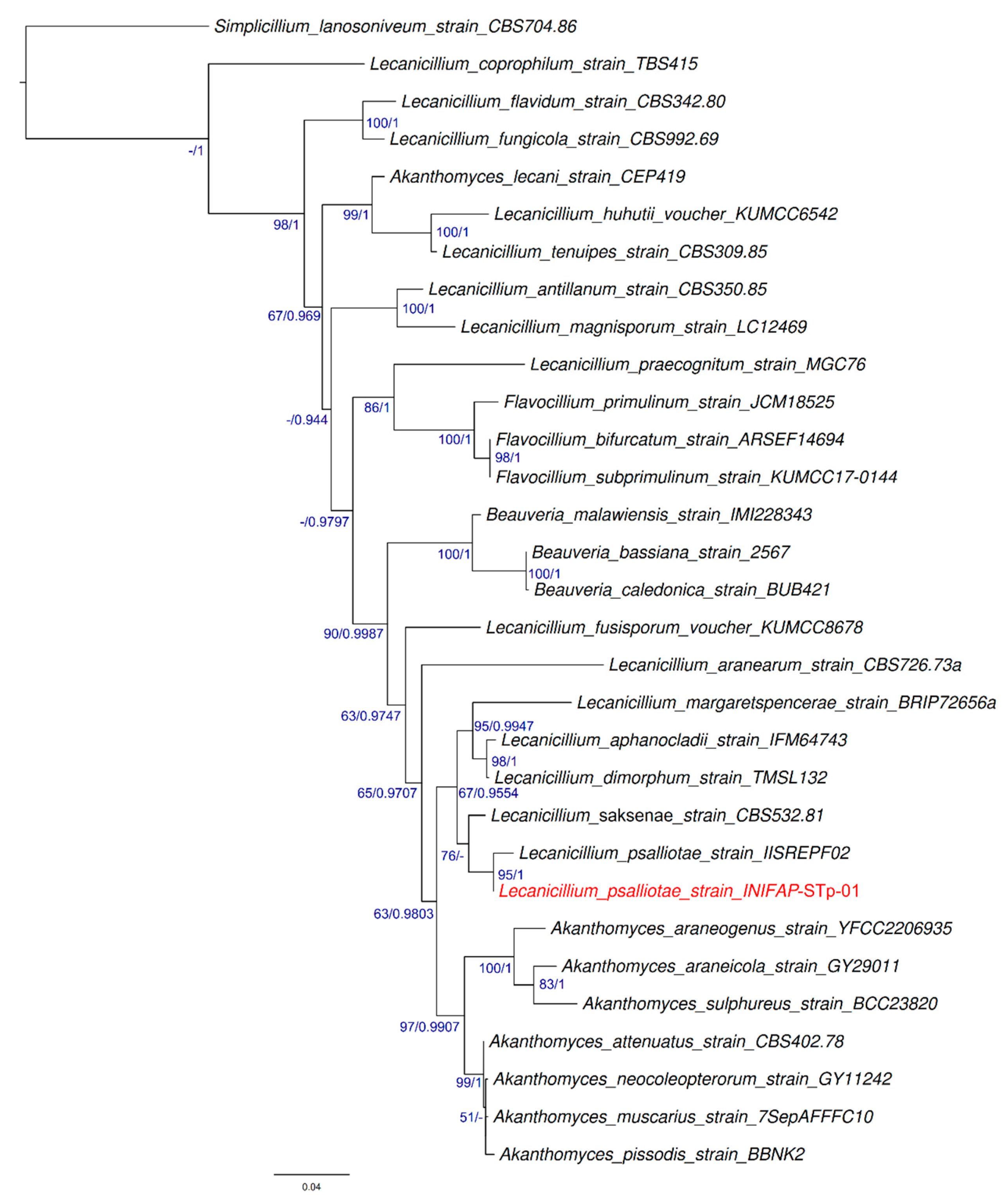
3.2. Molecular Taxonomy
3.3. Ovicidal Activity of Fungal Liquid Culture Filtrates against Haemonchus contortus
3.4. Nematocidal Activity of Fungal Liquid Culture Filtrates against Haemonchus contortus Infective Larvae
3.5. Myco-Chemical Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hannula, S.E.; Morriën, E. Will fungi solve the carbon dilemma? Geoderma 2022, 413, 115767. [Google Scholar] [CrossRef]
- Li, G.; Chen, W.; Xu, S.; Xiong, S.; Zhao, J.; Liu, D.; Ding, G.; Li, J.; Wei, Y. Role of fungal communities and their interaction with bacterial communities on carbon and nitrogen component transformation in composting with different phosphate additives. Environ. Sci. Pollut. Res. 2023, 30, 44112–44120. [Google Scholar] [CrossRef] [PubMed]
- Saccá, M.L.; Barra-Caracciolo, A.; Di-Lenola, M.; Grenni, P. Ecosystem services provided by soil microorganisms. In Soil Biological Communities and Ecosystem Resilience, 1st ed.; Saccá, M.L., Barra-Caracciolo, A., Di-Lenola, M., Grenni, P., Eds.; Springer: Cham, Switzerland, 2017; pp. 9–24. [Google Scholar] [CrossRef]
- Sveen, T.R.; Hannula, S.E.; Bahram, M. Microbial regulation of feedbacks to ecosystem change. Trends Microbiol. 2023, 32, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Samsinakova, A.; Kalalova, S.; Daniel, M.; Dusbabek, F.; Honzakova, E.; Cerný, V. Entomogenous fungi associated with the tick Ixodes ricinus (L.). Folia Parasitol. 1974, 21, 39–48. [Google Scholar]
- Mantzoukas, S.; Kitsiou, F.; Natsiopoulos, D.; Eliopoulos, P.A. Entomopathogenic fungi: Interactions and applications. Encyclopedia 2022, 2, 646–656. [Google Scholar] [CrossRef]
- Abouamama, S.; Anis, B.; Abir, S.; Maroua, H.; Sirine, B. Amylolytic and antibacterial activity of filamentous fungi isolated from the rhizosphere of different plants grown in the Tamanghasset region. Heliyon 2023, 9, E14350. [Google Scholar] [CrossRef] [PubMed]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.; Louro, M.; Almeida, C.; Victório, A.C.; Melo, P.; Rodrigues, J.P.; Oliveira, M.; Paz-Silva, A.; Madeira de Carvalho, L. Isolation of saprophytic filamentous fungi from avian fecal samples and assessment of its predatory activity on coccidian oocysts. Sci. Rep. 2023, 13, 8965. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Huang, X.; Tian, B.; Wang, M.; Niu, Q.; Zhang, K. Isolation and characterization of a serine protease from the nematophagous fungus, Lecanicillium psalliotae, displaying nematicidal activity. Biotechnol. Lett. 2005, 27, 1123–1128. [Google Scholar] [CrossRef]
- Mendoza-de Gives, P.; Braga, F.R.; Araújo, J.V.D. Nematophagous fungi, an extraordinary tool for controlling ruminant parasitic nematodes and other biotechnological applications. Biocontrol Sci. Technol. 2022, 32, 777–793. [Google Scholar] [CrossRef]
- Wang, Y.B.; Wang, Y.; Fan, Q.; Duan, D.E.; Zhang, G.D.; Dai, R.Q.; Dai, Y.D.; Zheng, W.B.; Chen, Z.H.; Li, D.D.; et al. Multigene phylogeny of the family Cordycipitaceae (Hypocreales): New taxa and the new systematic position of the Chinese cordycipitoid fungus Paecilomyces hepiali. Fungal Divers. 2020, 103, 1–46. [Google Scholar] [CrossRef]
- Gül, E.; Babaroğlu, N.E.; Demirci, F. Characterization of Lecanicillium psalliotae and Akanthomyces muscarium from Sunn pests (Eurygaster spp.). J. Cent. Eur. Agric. 2022, 23, 526–532. [Google Scholar] [CrossRef]
- Arevalo, J.; Hidalgo-Díaz, L.; Martins, I.; Souza, J.F.; Castro, J.M.C.; Carneiro, R.M.D.; Tigano, M.S. Cultural and morphological characterization of Pochonia chlamydosporia and Lecanicillium psalliotae isolated from Meloidogyne mayaguensis eggs in Brazil. Trop. Plant Pathol. 2009, 34, 158–163. [Google Scholar] [CrossRef]
- Krif, G.; Lahlali, R.; El Aissami, A.; Laasli, S.E.; Mimouni, A.; Dababat, A.A.; Zoubi, B.; Mokrini, F. Potential Effects of Nematophagous Fungi Against Meloidogyne javanica Infection of Tomato Plants Under in vitro and in vivo Conditions. J. Crop Health 2024, 1–11. [Google Scholar] [CrossRef]
- Majdi, N.; Traunspurger, W. Free-living nematodes in the freshwater food web: A review. J. Nematol. 2015, 47, 28. [Google Scholar] [PubMed]
- Strydom, T.; Lavan, R.P.; Torres, S.; Heaney, K. The Economic Impact of Parasitism from Nematodes, Trematodes and Ticks on Beef Cattle Production. Animals 2023, 13, 1599. [Google Scholar] [CrossRef] [PubMed]
- Tapia-Vázquez, I.; Montoya-Martínez, A.C.; De los Santos-Villalobos, S.; Ek-Ramos, M.J.; Montesinos-Matías, R.; Martínez-Anaya, C. Root-knot nematodes (Meloidogyne spp.) a threat to agriculture in Mexico: Biology, current control strategies, and perspectives. World J. Microbiol. Biotechnol. 2022, 38, 26. [Google Scholar] [CrossRef] [PubMed]
- Sikandar, A.; Jia, L.; Wu, H.; Yang, S. Meloidogyne enterolobii risk to agriculture, its present status and future prospective for management. Front. Plant Sci. 2023, 13, 1093657. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Tokiwa, T.; Sugiyama, H.; Shiroyama, M.; Morishima, Y.; Watanabe, S.; Sasamori, T.; Kondo, M.; Mano, T.; Tsuruga, H. Trichinella T9 in wild bears in Japan: Prevalence, species/genotype identification, and public health implications. Int. J. Parasitol. Parasites Wildl. 2023, 21, 264–268. [Google Scholar] [CrossRef]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. The pathophysiology, ecology and epidemiology of Haemonchus contortus infection in small ruminants. Adv. Parasitol. 2016, 93, 95–143. [Google Scholar]
- Arsenopoulos, K.V.; Fthenakis, G.C.; Katsarou, E.I.; Papadopoulos, E. Haemonchosis: A challenging parasitic infection of sheep and goats. Animals 2021, 11, 363. [Google Scholar] [CrossRef]
- Flay, K.J.; Hill, F.I.; Muguiro, D.H. A Review: Haemonchus contortus infection in pasture-based sheep production systems, with a focus on the pathogenesis of anaemia and changes in haematological parameters. Animals 2022, 12, 1238. [Google Scholar] [CrossRef] [PubMed]
- Soares, V.M.; Pereira, J.G.; Barreto, F.; Jank, L.; Rau, R.B.; Ribeiro, C.B.D.; da Silva, W.P. Residues of veterinary drugs in animal products commercialized in the border region of Brazil, Argentina, and Uruguay. J. Food Prot. 2022, 85, 980–986. [Google Scholar] [CrossRef] [PubMed]
- Goodenough, A.E.; Webb, J.C.; Yardley, J. Environmentally-realistic concentrations of anthelmintic drugs affect survival and motility in the cosmopolitan earthworm Lumbricus terrestris (Linnaeus, 1758). Appl. Soil Ecol. 2019, 137, 87–95. [Google Scholar] [CrossRef]
- Sands, B.; Noll, M. Toxicity of ivermectin residues in aged farmyard manure to terrestrial and freshwater invertebrates. Insect Conserv. Divers. 2022, 15, 9–18. [Google Scholar] [CrossRef]
- Fissiha, W.; Kinde, M.Z. Anthelmintic resistance and its mechanism: A review. Infect. Drug Resist. 2021, 14, 5403–5410. [Google Scholar] [CrossRef]
- Jouffroy, S.; Bordes, L.; Grisez, C.; Sutra, J.F.; Cazajous, T.; Lafon, J.; Dumont, N.; Chastel, M.; Vial-Novella, C.; Achard, D.; et al. First report of eprinomectin-resistant isolates of Haemonchus contortus in 5 dairy sheep farms from the Pyrénées Atlantiques département in France. Parasitology 2023, 150, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Cedillo-Borda, M.; López-Arellano, M.E.; Reyes-Guerrero, D.E. In vitro assessment of ivermectin resistance and gene expression profiles of P-glycoprotein genes from Haemonchus contortus (L3). Bio-Protocol 2020, 101, e3851. [Google Scholar] [CrossRef]
- Becerril-Gil, M.M.N.; Estrada-Flores, J.G.; González-Cortazar, M.; Zamilpa, A.; Endara-Agramont, Á.R.; Mendoza-de Gives, P.; López-Arellano, M.E.; Olmedo-Juárez, A. Bioactive compounds from the parasitic plant Arceuthobium vaginatum inhibit Haemonchus contortus egg hatching. Rev. Bras. Parasitol. Vet. 2023, 33, e013223. [Google Scholar] [CrossRef]
- de Jesús-Martínez, X.; Rivero-Pérez, N.; González-Cortazar, M.; Olivares-Pérez, J.; Zamilpa, A.; Zaragoza-Bastida, A.; Mendoza-de Gives, P.; Rojas-Hernández, S.; Flores-Franco, G.; Olmedo-Juárez, A. Ovicidal and larvicidal effects of a hydroalcoholic extract from Cyrtocarpa procera leaves against Haemonchus contortus. Agrociencia 2024, 58, 2957. [Google Scholar] [CrossRef]
- Woo, P.C.; Ngan, A.H.; Chui, H.K.; Lau, S.K.; Yuen, K.Y. Agar block smear preparation: A novel method of slide preparation for preservation of native fungal structures for microscopic examination and long-term storage. J. Clin. Microbiol. 2010, 48, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Zare, R.; Gams, W. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwig. 2001, 73, 1–50. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Stielow, J.B.; Levesque, C.A.; Seifert, K.A.; Meyer, W.; Irinyi, L.; Smits, D.; Robert, V. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia-Mol. Phylogeny Evol. Fungi 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Olmedo-Juárez, A.; Rojo-Rubio, R.; Zamilpa, A.; Mendoza-de Gives, P.; Arece-García, J.; López-Arellano, M.E.; von Son-de Fernex, E. In vitro larvicidal effect of a hydroalcoholic extract from Acacia cochliacantha leaf against ruminant parasitic nematodes. Vet. Res. Commun. 2017, 41, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Núñez, E.J.; Zamilpa, A.; González-Cortazar, M.; Olmedo-Juárez, A.; Cardoso-Taketa, A.; Sánchez-Mendoza, E.; Tapia-Maruri, D.; Salinas-Sánchez, D.O.; Mendoza-de Gives, P. Isorhamnetin: A nematocidal flavonoid from Prosopis laevigata leaves against Haemonchus contortus eggs and larvae. Biomolecules 2020, 10, 773. [Google Scholar] [CrossRef] [PubMed]
- Sukarno, N.; Kurihara, Y.; Park, J.Y.; Inaba, S.; Ando, K.; Harayama, S.; Ilyas, M.; Mangunwardoyo, W.; Sjamsuridzal, W.; Yuniarti, E.; et al. Lecanicillium and Verticillium species from Indonesia and Japan including three new species. Mycoscience 2009, 50, 369–379. [Google Scholar] [CrossRef]
- Sreeja, P.; Rani, R.O.; Chellappan, M. Characterization of insecticidal metabolite oosporein in Lecanicillium saksenae (Kushwaha) Kurihara and Sukarno, a geographically distinct isolate from Kerala, India. Toxicon 2023, 230, 107176. [Google Scholar]
- Nagaoka, T.; Nakata, K.; Kouno, K. Antifungal activity of oosporein from an antagonistic fungus against Phytophthora infestans. Z. Naturforschung C 2004, 59, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal biosynthesis of the bibenzoquinone oosporein to evade insect immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef]
- Mc Namara, L.; Dolan, S.K.; Walsh, J.M.; Stephens, J.C.; Glare, T.R.; Kavanagh, K.; Griffin, C.T. Oosporein, an abundant metabolite in Beauveria caledonica, with a feedback induction mechanism and a role in insect virulence. Fungal Biol. 2019, 123, 601–610. [Google Scholar] [CrossRef]
- Hofstätter, B.D.M.; da Silva-Fonseca, A.O.; de Souza-Maia-Filho, F.; de Souza-Silveira, J.; Persici, B.M.; Pötter, L.; Silveira, A.; Antoniolli, Z.I.; Pereira, D.I.B. Effect of Paecilomyces lilacinus, Trichoderma harzianum and Trichoderma virens fungal extracts on the hatchability of Ancylostoma eggs. Rev. Iberoamer. Micol. 2017, 34, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Braga, F.R.; Araújo, J.V.; Carvalho, R.O.; Silva, A.R.; Araujo, J.M.; Soares, F.E.F.; Geniêr, H.L.A.; Ferreira, S.R.; Queiroz, J.H. Ovicidal action of a crude enzymatic extract of the fungus Pochonia chlamydosporia against cyathostomin eggs. Vet. Parasitol. 2010, 172, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, T.M.; Pervaz, I.P. In vitro evaluation of ovicidal and larvicidal effects of culture filtrate of Verticillium chlamydosporium against Meloidogyne javanica. Int. J. Agric. Biol. 2003, 5, 576–579. [Google Scholar]
- Regaieg, H.; Ciancio, A.; Raouani, N.H.; Grasso, G.; Rosso, L. Effects of culture filtrates from the nematophagous fungus Verticillium leptobactrum on viability of the root-knot nematode Meloidogyne incognita. World J. Microbiol. Biotechnol. 2010, 26, 2285–2289. [Google Scholar] [CrossRef]
- Cortes-Morales, J.A.; Olmedo-Juárez, A.; Trejo-Tapia, G.; González-Cortazar, M.; Domínguez-Mendoza, B.E.; Mendoza-de Gives, P.; Zamilpa, A. In vitro ovicidal activity of Baccharis conferta Kunth against Haemonchus contortus. Exp. Parasitol. 2019, 197, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Patidar, P.; Prasad, L.; Sagar, S.; Sirohi, A.; Saharan, M.S.; Dhillon, M.K.; Singh, V.K.; Bag, T.K. Chemo-profiling of Purpureocillium lilacinum and Paecilomyces variotii isolates using GC-MS analysis, and evaluation of their metabolites against M. incognita. PLoS ONE 2024, 19, e0297925. [Google Scholar] [CrossRef] [PubMed]
- Ocampo-Gutiérrez, A.Y.; Hernández-Velázquez, V.M.; Aguilar-Marcelino, L.; Cardoso-Taketa, A.; Zamilpa, A.; López-Arellano, M.E.; González-Cortázar, M.; Hernández-Romano, J.; Reyes-Esteban, M.; Mendoza-de Gives, P. Morphological and molecular characterization, predatory behaviour and effect of organic extracts of four nematophagous fungi from Mexico. Fungal Ecol. 2021, 49, 101004. [Google Scholar] [CrossRef]
- Angeles-Hernández, S.; Torres-Hernández, G.; Alonso-Díaz, M.A.; von Son-de-Fernex, E.; Aguilar-Marcelino, L.; González-Garduño, R.; Becerril-Pérez, C.M.; Alcántara-Carbajal, J.L.; Vargas-López, S.; Olmedo-Juárez, A.; et al. Effect of an Arthrobotrys musiformis (Fungi: Orbiliales) culture filtrate on the population of gastrointestinal parasitic nematode eggs in faeces of grazing lambs. Vet. Parasitol. Reg. Stud. Rep. 2021, 24, 100565. [Google Scholar] [CrossRef] [PubMed]
- Bloch, P.; Tamm, C.; Bollinger, P.; Petcher, T.J.; Weber, H.P. Pseurotin, a new metabolite of Pseudeurotium ovalis STOLK having an unusual hetero-spirocyclic system. (Preliminary Communication). Helv. Chim. Acta 1976, 59, 133–137. [Google Scholar] [CrossRef]
- Niu, X.M. Secondary metabolites from Pochonia chlamydosporia and other species of Pochonia. In Perspectives in Sustainable Nematode Management through Pochonia chlamydosporia Applications for Root and Rhizosphere Health, 1st ed.; Manzanilla-López, R., Lopez-Llorca, L., Eds.; Springer: Cham, Switzerland, 2017; pp. 131–168. [Google Scholar]
- de Souza-Gouveia, A.; Monteiro, T.S.A.; Balbino, H.M.; de Magalhães, F.C.; Ramos, M.E.S.; Moura, V.A.S.; Luiz, P.H.D.; de Almeida-Oliveira, M.G.; Freitas, L.G.; de Oliveira Ramos, H.J. Inoculation of Pochonia chlamydosporia triggers a defense response in tomato roots, affecting parasitism by Meloidogyne javanica. Microbiol. Res. 2023, 266, 127242. [Google Scholar] [CrossRef]
- Ibragimova, Z.B.; Anan’ko, G.G.; Kostina, N.E.; Teplyakova, T.V.; Mazurkova, N.A. Toxicity and antiviral activity of the extracts of submerged mycelium of nematophagous Duddingtonia flagrans fungus in vero cell culture. Bull. Exp. Biol. Med. 2015, 160, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Guo, Y.; Wang, X.; Gao, M.; Liu, T.; Ma, Y.; Zhang, Y.; Li, Q.; Wang, R.; You, X. Preparation and application of biocontrol formulation of nematode-trapping fungus Duddingtonia flagrans. Vet. Parasitol. 2024, 327, 110119. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Pineda, M.C.; Bulla-Castañeda, D.M.; Garcia-Corredor, D.J.; Pulido-Medellín, M.O. Tannins as a Biological Control of Gastrointestinal Nematodes in Sheep. Rev. Electron. Vet. 2020, 21, 13–22. [Google Scholar]
- Fomum, S.W.; Nsahlai, I.V. In vitro nematicidal activity of plant species possessing alkaloids and tannins. Cogent Food Agric. 2017, 3, 1334295. [Google Scholar] [CrossRef]
- da Silva, G.D.; de Lima, H.G.; de Sousa, N.B.; de Jesus-Genipapeiro, I.L.; Uzêda, R.S.; Branco, A.; Botura, M.B. In vitro anthelmintic evaluation of three alkaloids against gastrointestinal nematodes of goats. Vet. Parasitol. 2021, 296, 109505. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Mean | Range |
|---|---|---|
| Phialide length (µm) | 21.75 | 15.6–33.8 |
| Phialide width (µm) | 1.78 | 1.26–2.52 |
| Macroconidia length (µm) | 9.06 | 6.74–11.94 |
| Macroconidia width (µm) | 2.08 | 1.56–2.61 |
| Microconidia length (µm) | 4.94 | 4.27–5.73 |
| Microconidia width (µm) | 1.67 | 1.25–2.04 |
| Phialides/whorl | 3 | 1–5 |
| Media | Concentration (mg/mL) | Larvae (Mean) | Eggs (Mean) | Total Count (Mean) | Egg-Hatching Inhibition (Mean ± SE) * |
|---|---|---|---|---|---|
| SPDB | 0 | 127.56 | 2.45 | 130.25 | 1.88 ± 1.14 a |
| 10 | 53.34 | 71.23 | 125.00 | 56.98 ± 15.40 b | |
| 15 | 19.78 | 99.55 | 119.00 | 83.66 ± 6.03 bc | |
| 20 | 6.13 | 128.68 | 134.75 | 95.49 ± 1.80 c | |
| 25 | 2.67 | 99.51 | 102.38 | 97.20 ± 0.80 c | |
| CzDoxB | 0 | 115.32 | 1.63 | 117.00 | 1.39 ± 0.60 a |
| 10 | 36.09 | 54.70 | 90.75 | 60.28 ± 4.00 b | |
| 15 | 33.54 | 67.86 | 101.38 | 66.94 ± 4.45 b | |
| 20 | 13.76 | 94.58 | 108.57 | 87.11 ± 3.05 c | |
| 25 | 0.97 | 103.27 | 104.25 | 99.06 ± 0.20 c |
| Media | Concentration (mg/mL) | Dead/Total Larvae (Mean) | Larval Mortality (%) (Mean ± SE) * |
|---|---|---|---|
| SPDB | NF | 4.42/90.67 | 4.87 ± 1.05 a |
| 25 | 3.98/101.09 | 3.94 ± 0.77 a | |
| 50 | 19.96/96.63 | 20.66 ± 1.53 b | |
| 100 | 58.77/108.29 | 54.27 ± 4.56 c | |
| CzDoxB | NF | 3.32/83.38 | 3.98 ± 0.74 a |
| 25 | 44.23/94.45 | 46.82 ± 7.15 b | |
| 50 | 69.77/76.50 | 91.20 ± 2.80 c | |
| 100 | 85.27/88.09 | 96.80 ± 0.68 c |
| Metabolite and Reagent | Colorimetric Reaction | Lecanicillium psalliotae | |
|---|---|---|---|
| SPDB | CzDoxB | ||
| Alkaloids Dragendorff Mayer Wagner | Turbidity or precipitate (red to orange, white to cream, and brown) | +++ + ++ | +++ + + |
| Coumarins Bornträger | Yellow fluorescence (U.V) | - | - |
| Flavonoids Mg2+ and HCL | Red, orange, and violet colour | - | - |
| Tannins Ferric chloride (FeCl3) | Hydrolysable tannins (blue) Condensed tannins (green) | +++ + | + ++ |
| Confirmation | |||
| Solution of gelatine | Precipitate white | + | + |
| Gelatine and saline solution | Precipitate white | + | + |
| Saline solution | Precipitate white | - | - |
| Triterpenes/Sterols Liebermann–Buchard | Colour blue, blue-green (sterols) | - | - |
| Salkowski | Red to purple (triterpene) | - | - |
| Saponins Water | Foam formation | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Anzúrez, G.; Mendoza-de Gives, P.; Alonso-Díaz, M.Á.; von Son-de Fernex, E.; Paz-Silva, A.; López-Arellano, M.E.; Olmedo-Juárez, A. Lecanicillium psalliotae (Hypocreales: Cordycipitaceae) Exerts Ovicidal and Larvicidal Effects against the Sheep Blood-Feeding Nematode Haemonchus contortus through Its Liquid Culture Filtrates. Pathogens 2024, 13, 588. https://doi.org/10.3390/pathogens13070588
Pérez-Anzúrez G, Mendoza-de Gives P, Alonso-Díaz MÁ, von Son-de Fernex E, Paz-Silva A, López-Arellano ME, Olmedo-Juárez A. Lecanicillium psalliotae (Hypocreales: Cordycipitaceae) Exerts Ovicidal and Larvicidal Effects against the Sheep Blood-Feeding Nematode Haemonchus contortus through Its Liquid Culture Filtrates. Pathogens. 2024; 13(7):588. https://doi.org/10.3390/pathogens13070588
Chicago/Turabian StylePérez-Anzúrez, Gustavo, Pedro Mendoza-de Gives, Miguel Ángel Alonso-Díaz, Elke von Son-de Fernex, Adolfo Paz-Silva, María Eugenia López-Arellano, and Agustín Olmedo-Juárez. 2024. "Lecanicillium psalliotae (Hypocreales: Cordycipitaceae) Exerts Ovicidal and Larvicidal Effects against the Sheep Blood-Feeding Nematode Haemonchus contortus through Its Liquid Culture Filtrates" Pathogens 13, no. 7: 588. https://doi.org/10.3390/pathogens13070588