
Insights into Peptidyl-Prolyl cis-trans Isomerases from Clinically Important Protozoans: From Structure to Potential Biotechnological Applications

 , , and
, , and
Abstract
:
1. Introduction
2. Parasite PPIases: Disease, Genome Database, and Structural Characteristics
2.1. Anaerobic or Microaerophilic Protozoan Parasites
2.1.1. Trichomonas vaginalis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt | TrichDB 2 | TrichDB 3 | NCBI | PDB | PPIase Name | Localization 4 | Function 4 | References |
|---|---|---|---|---|---|---|---|---|
| A2FJP1 | TVAG_370440 | TVAGG3_0054050 | XP_001307803 | TvCyP14 | Nucleus | [29] | ||
| A2EC21 | TVAG_137880 | TVAGG3_0269460 | XP_001322019.1 | TvCyP18 | Cytoplasm | [29] | ||
| A2DT06 | TVAG_004440 | TVAGG3_0649370 | XP_001328636.1 | 5YB9 | TvCyP19 (TvCyP1) | Cytoplasm hydrogenosomes, cytoplasm, and membrane | Protein trafficking | [30,31] |
| A2F1H0 | TVAG_027250 | TVAGG3_0947870 | XP_001314072 | TvCyP19.2 | Cytoplasm | [29] | ||
| A2FAA8 | TVAG_047830 | TVAGG3_0485720 | XP_001311112 | TvCyP19.8 | Cytoplasm | [29] | ||
| A2DLL4 | TVAG_062520 | TVAGG3_0580400 | XP_001579633.1 | 6LXO | TvCyP19.9 TvCyP2 | Cytoplasm ER, cytoplasm, and membrane | Protein trafficking | [32,33] |
| A2E5J4 | TVAG_038810 | TVAGG3_0240350 | XP_001324258 | TvCyP20 | Cytoplasm | [29] | ||
| A2FIV3 | TVAG_078570 | TVAGG3_0462310 | XP_001308098 | TvCyP21 | Cytoplasm | [29] | ||
| A2DKZ9 | TVAG_146960 | TVAGG3_0362200 | XP_001579938 | TvCyP22 | Cytoplasm | [29] | ||
| A2FTU8 | TVAG_27739 | TVAGG3_0951420 | XP_001304599 | TvCyP23 | Cytoplasm | [29] | ||
| A2E6H3 | TVAG_106810 | TVAGG3_0040330 | XP_001324009.1 | TvCyP37 | Nucleus | [29] | ||
| A2GDG2 | TVAG_583670 | TVAGG3_0820230 | XP_001297735.1 | TvCyP44 | Cytoplasm and nucleus | [29] | ||
| A2DEW6 | TVAG_172150 | TVAGG3_0530670 | XP_001581944.1 | TvCyP63 | Nucleus | [29] | ||
| A2DA37 | TVAG_476140 | TVAGG3_0266130 | XP_001583671.1 | TvFKBP12 | Cytoplasm | [29] | ||
| A2DYS7 | TVAG_426610 | TVAGG3_0538360 | XP_001326690.1 | TvFKBP15.1 | ER | [29] | ||
| A2G763 | TVAG_062070 | TVAGG3_0922950 | XP_001299933.1 | TvFKBP15.2 | ER | [29] | ||
| A2FYT1 | TVAG_435000 | TVAGG3_0194750 | XP_001302863.1 | TvFKBP19 | Cytoplasm | [29] | ||
| A2F0D0 | TVAG_292580 | TVAGG3_0216440 | XP_001330357.1 | TvFKBP20 | Cytoplasm | [29] | ||
| A2EV02 | TVAG_368970 | TVAGG3_0441630 | XP_001315748.1 | TvFKBP30 | Cytoplasm | [29] | ||
| A2EC50 | TVAG_413760 | TVAGG3_0204900 | XP_001321950.1 | TvFKBP32 | Cytoplasm | [29] | ||
| A2G9L9 | TVAG_428320 | TVAGG3_0107870 | XP_001299079.1 | TvFKBP33 | Cytoplasm | [29] | ||
| A2FER9 | TVAG_140950 | TVAGG3_0603860 | XP_001309536.1 | TvFKBP63 | [29] | |||
| A2ECU0 | TVAG_102340 | TVAGG3_0563910 | XP_001321708.1 | TvPar17.84 | Cytoplasm | [29] | ||
| A2ED59 | TVAG_420360 | TVAGG3_0425040 | XP_001321637.1 | TvPar17.87 | Cytoplasm and nucleus | [29] | ||
| A2EWG2 | TVAG_325610 | TVAGG3_0877000 | XP_001315212.1 | TvPar102 | Cytoplasm and nucleus | [29] |
2.1.2. Entamoeba histolytica
2.1.3. Giardia intestinalis
| Isolate | UniProt | GiardiaDB | NCBI | PDB | PPIase Name | Localization 3 | Function 3 | References |
|---|---|---|---|---|---|---|---|---|
| G. intestinalis WB | GiCyP19 (GiCyP1) | [58] | ||||||
| A8BC67 | GL50803_0017163 | XP_001707838.1 | GiCyP18 | Cytoplasm Secreted | Virulence factor | [29,59,60] | ||
| A8BJP8 | GL50803_0017000 | XP_001706629.1 | GiCyP21 | Cytoplasm Secreted | [29,59] | |||
| G. intestinalis DH | V6TEN6 | DHA2_17000 | GiCyP25 | Membrane | [29] | |||
| G. intestinalis GS | C6LQJ1 | GL50581_1019 | GiCyP18 | Secreted | [29] | |||
| C6LR04 | GL50581_1186 | GiCyP21 | Secreted | [43] | ||||
| G. intestinalis WB | Q8I6M8 | GL50803_10450 | XP_001709141.1 | 2LGO | GiFKBP12 | Secreted | [59,61] | |
| A8B770 | GL50803_7246 | XP_001709155.1 | GiFKBP13 | Cytoplasm | [29] | |||
| A8BHU4 | GL50803_101339 | XP_001706925.1 | GiFKBP24 | Cytoplasm Secreted | [29,59] | |||
| A8BUZ7 | GL50803_42780 | XP_001704692.1 | GiFKBP28 | |||||
| A8BAF3 | GL50803_3643 | XP_001708385.1 | GiFKBP38 | Cytoplasm Secreted | [29,59] | |||
| A8BK50 | GL50803_10570 | XP_001706462.1 | GiFKBP39 | |||||
| G. intestinalis DH | V6TL25 | DHA2_151252 | GiFKBP29 | |||||
| G. intestinalis GS | C6LUS9 | GL50581_2531 | GiFKBP12 | Secreted | [59] | |||
| C6LPP4 | GL50581_711 | GiFKBP13 | ||||||
| C6LXS7 | GL50581_3593 | GiFKBP24 | Secreted | [59] | ||||
| C6LY30 | GL50581_3701 | GiFKBP28 | ||||||
| C6LPE9 | GL50581_614 | GiFKBP38 | Secreted | [59] | ||||
| C6M084 | GL50581_4472 | GiFKBP39 |
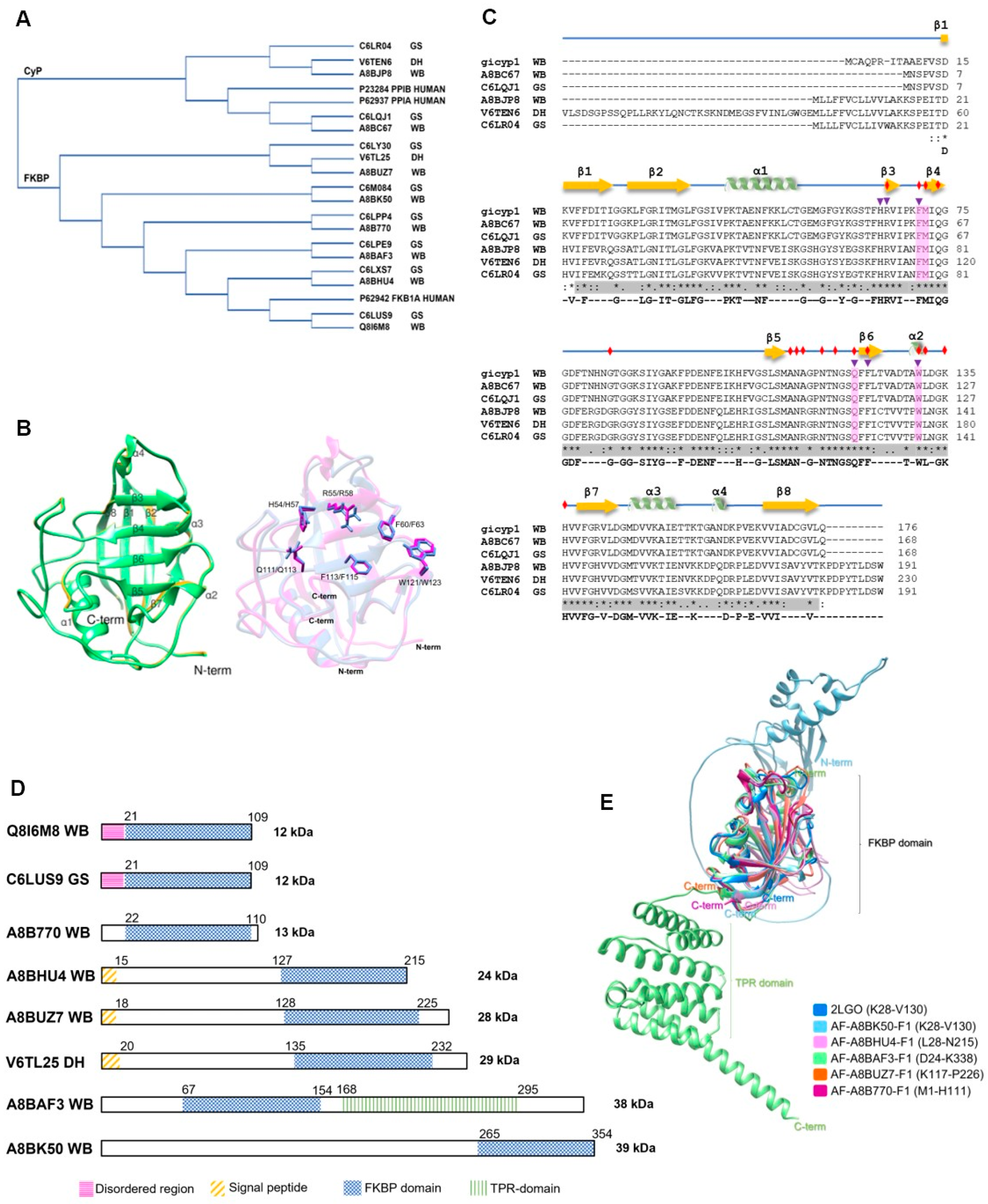
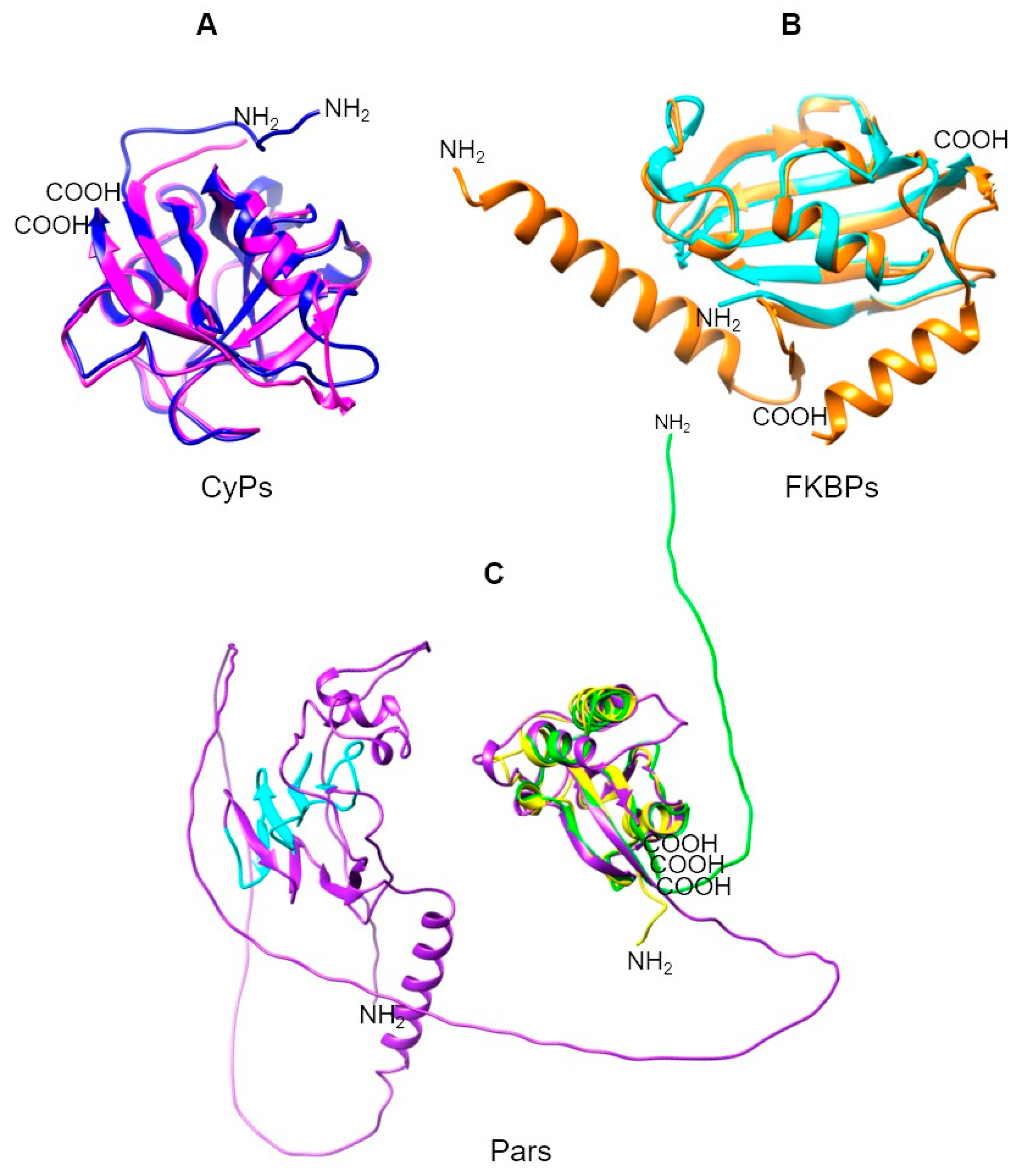
2.2. Trypanosomatid Parasites
2.2.1. Trypanosoma cruzi and Trypanosoma brucei gambiense
| Parasite | UniProt | TriTrypDB | NCBI | PDB | PPIase Name | Localization 2 | Function 2 | References |
|---|---|---|---|---|---|---|---|---|
| T. cruzi CL Brener | Q4E4L9 | TcCLB.506925.300 (CYPA) | XP_821578.1 | TcCyP19 | Extracellular space | Promotes ROS production in host cells | [72,74,75,76,77] | |
| Q4DC03 | TcCLB.507009.100 | XP_811912.1 | TcCyP20 | |||||
| Q4DNC9 | TcCLB.507521.70 | XP_815879.1 | 1XO7 | TcCyP21 | [72,76] | |||
| Q4DI85 | TcCLB.504035.70 | XP_814080.1 | TcCyP22 | Mitochondria | Cell death regulation | [72,78] | ||
| Q4CXV1 | TcCLB.506413.80 | XP_806960.1 | TcCyP24 | |||||
| Q4DFL3 | TcCLB.508323.94 | XP_813175.1 | TcCyP25 | [72,76] | ||||
| Q4D4K3 | TcCLB.503885.40 | XP_809302.1 | TcCyP26 | |||||
| Q4CX88 | TcCLB.509499.10 | XP_806737.1 | TcCyP28 | [72,76] | ||||
| Q4DQI8 | TcCLB.505807.10 | XP_816616.1 | TcCyP29 | |||||
| Q4DNS3 | TcCLB.511589.50 | XP_816007.1 | TcCyP30 | Membrane | [29,72] | |||
| Q4DM35 | TcCLB.511577.40 (CYP35) | XP_815421.1 | TcCyP35 (TcCyP34) | [72,76] | ||||
| Q4DVC9 | TcCLB.511217.120 | XP_818332.1 | TcCyP35.3 (TcCyP35) | [72] | ||||
| Q4E4G0 | TcCLB.506885.400 (CYP40) | XP_821542.1 | TcCyP38 (TcCyP40) | [29,72] | ||||
| Q4DG41 | TcCLB.510761.44 | XP_813344.1 | TcCyP42 | Membrane | [29,72] | |||
| Q4D1M5 | TcCLB.504215.10 | XP_808273.1 | TcCyP103 TcCyP110 | [72] | ||||
| T. cruzi Y | Q09734 | TcYC6_0113560 | CAA49346.1 | 1JVW | TcFKBP22 (TcMIP) | Extracellular space | Host cell entry/invasion | [73,79] |
| T. cruzi CL Brener | Q4D5W5 | TcCLB.508169.69 | XP_809772.1 | TcFKBP12 | ||||
| Q4DFL5 | TcCLB.508323.84 | XP_813174.1 | TcFKBP12.2 | |||||
| Q4D7F5 | TcCLB.511731.89 | XP_810317.1 | TcFKBP35 | |||||
| Q4CZN2 | TcCLB.511353.10 | XP_807578.1 | TcFKBP52 | |||||
| Q4CYE6 | TcCLB.507629.39 | XP_807152.1 | TcFKBP93 | |||||
| T. cruzi CL Brener | Q4D8F7 | TcCLB.508567.70 (Pin1) | XP_810661.1 | TcPar12.6 (TcPin1) | Cytosol | [80,81] | ||
| Q4D394 | TcCLB.506697.50 | XP_808848.1 | TcPar13 (TcPar14) | [82] | ||||
| Q4D9J4 | TcCLB.506857.60 (Par45) | XP_811046.1 | TcPar45 | Nucleus | [82] |
2.2.2. Leishmania major and Leishmania donovani
2.3. Apicomplexan Parasites
2.3.1. Plasmodium falciparum and Plasmodium vivax
| UniProt | PlasmoDB | NCBI | PDB | PPIase Name | Localization 3 | References |
|---|---|---|---|---|---|---|
| Q8IIK3 | PF3D7_1116300 (CYP19C) | XP_001347841.1 | PfCyP18.6 (PfCyP19C) | Nucleus (Spliceosome) | [29,107,110] | |
| Q76NN7 | PF3D7_0322000 (CYP19A) | XP_001351290.1 | 1QNG | PfCyP19 (PfCyP19A) | Cytoplasm | [29,107,110,111,112,113] |
| Q8IIK8 | PF3D7_1115600 (CYP19B) | XP_001347835.1 | PfCyP22 (PfCyP19B) | Cytoplasm Membrane | [29,107,113,114,115] | |
| Q8I3I0 | PF3D7_0528700 (CYP23) | XP_001351841.1 | PfCyP23 | Nucleus (Spliceosome) | [29,102,107,109,110] | |
| Q8I6S4 | PF3D7_0804800 (CYP24) | XP_001349469.1 | PfCyP25(PfCyP24) | Membrane | [29,107,110,116,117] | |
| Q8I621 | PF3D7_1202400 (CYP26) | XP_001350433.1 | PfCyP263 | Cytoplasm | ||
| Q8I5Q4 | PF3D7_1215200 (CYP32) | XP_001350556.1 | PfCyP32 | Cytoplasm and Mitochondria | [29,107,110] | |
| Q8ILM0 | PF3D7_1423200 (CYP52) | XP_001348397.2 | PfCyP53 (PfCyP52) | Nucleus (Spliceosome) | [29,107,110] | |
| Q8I2K8 | PF3D7_0930600 (CYP72) | XP_001352173.1 | PfCyP72 | Nucleus | [11,29] | |
| Q8IAN0 | PF3D7_0803000 (CYP81) | XP_001349484.1 | PfCyP81 | Nucleus | [11,29] | |
| Q8I402 | PF3D7_0510200 (CYP87) | XP_001351660.1 | 2FU0 | PfCyP87 | Nucleus (Spliceosome) | [29,107,108] |
| C0H5B2 | PF3D7_1313300 | XP_002809009.1 | PfFKBP25.6 | |||
| Q8I4V8 | PF3D7_1247400 | XP_001350859.1 | 2OFN | PfFKBP35 | Cytoplasm and Nucleus | [29,109,118,119,120] |
| UniProt | PlasmoDB | NCBI | PDB | PPIase Name | Localization 3 | References |
|---|---|---|---|---|---|---|
| A0A1G4HCW7 | PVP01_0916900 (CYP19C) | XP_001615280.1 | PvCyP18.5 | |||
| A0A1G4HBM6 | PVP01_0818200 (CYP19A) | XP_001614493.1 | PvCyP19 | |||
| A0A1G4HCM3 | PVP01_0916400 | XP_001615276.1 | PvCyP21 | |||
| A0A1G4HDR7 | PVP01_1005100 (CYP23) | XP_001613671.1 | PvCyP23 | |||
| A0A1G4H2Q1 | PVP01_1301700 (CYP26) | XP_001616500.1 | PvCyP26 | |||
| A0A1G4GR33 | PVP01_0115700 (CYP24) | XP_001608574.1 | PvCyP29 | |||
| A0A1G4H4X8 | PVP01_1434000 (CYP32) | XP_001617250.1 | PvCyP32 | |||
| A0A1G4HIV6 | PVP01_1325800 | CAG9475874.1 | PvCyP52 | |||
| A0A1G4HAY2 | PVP01_0729200 | XP_001614845.1 | PvCyP71 | |||
| A0A1G4GR20 | PVP01_0117200 (CYP81) | CAG9485095.1 | PvCyP65 | Nucleus | [29] | |
| A0A1G4HEA6 | PVP01_1023800 (CYP87) | XP_001613274.1 | PvCyP83 | |||
| A0A1G4H4D0 | PVP01_1414200 | XP_001617060.1 | 4JYS | PvFKBP25 3 | [116] | |
| A0A565A3M9 | PVP01_1464500 | XP_001613999.1 | 2KI3 | PvFKBP34 3 (PvFKBP35) | [120,122] |
2.3.2. Toxoplasma gondii
| UniProt | ToxoDB | NCBI | PDB | PPIase Name | Localization 3 | Function 3 | References |
|---|---|---|---|---|---|---|---|
| A0A125YZ79 | TGME49_289250 | XP_018636397.1 | TgCyP18 | Manipulates host cell responses | [130,131,132] | ||
| S8F7V1 | TGME49_221210 | XP_002369951.1 | TgCyP20 | Secreted | Manipulates host cell responses | [129,133] | |
| A0A125YV51 | TGME49_270560 | XP_002365722.1 | TgCyP21 | ||||
| A0A125YL73 | TGME49_285760 | XP_002369214.1 | TgCyP23 | [132] | |||
| A0A125YLU4 | TGME49_230520 | XP_002367963.2 | TgCyP26 | ||||
| S8FB56 | TGME49_238000 | XP_018637703.1 | TgCyP32 | ||||
| A0A125YQ35 | TGME49_262520 | XP_002365354.1 | TgCyP35 | ||||
| S8F5I7 | TGME49_205700 | XP_002367801.1 | TgCyP38 | Membrane | [29,129] | ||
| A0A125YVH7 | TGME49_241830 | XP_002366733.1 | 3BKP | TgCyP64 | [61] | ||
| A0A125YII8 | TGME49_229940 | XP_002367918.1 | TgCyP66.21 | Nucleus | |||
| S8GFQ1 | TGME49_227850 | XP_002366408.1 | TgCyP66.25 | Nucleus | |||
| A0A125YUW2 | TGME49_305940 | XP_002370366.1 | 3BO7 | TgCyP69 | [61] | ||
| S8FD30 | TGME49_320640 | XP_002369921.1 | TgCyP86 | ||||
| S8GFX3 | TGME49_228360 FKBP-12 | XP_002366458.1 | TgFKBP38 | Membrane | [29] | ||
| S8F5H8 | TGME49_285850 | XP_002369223.1 | TgFKBP46 | ||||
| Q4VKI5 | TGME49_283850 | XP_018637740.1 | TgFCBP57 2 | [134] | |||
| A0A125YIR1 | TGME49_318275 | XP_018637996.1 | TgFKBP64 | ||||
| S8F128 | TGME49_258625 | XP_018637023.1 | TgFKBP66 | ||||
| A0A125YRG0 | TGME49_258930 | XP_002365107.1 | TgPar13 | ||||
| S8EUZ2 | TGME49_228040 | XP_002366427.1 | TgPar96 |
2.3.3. Cryptosporidium parvum and Cryptosporidium hominis
3. Localization and Functions of PPIases in Parasites
4. Recombinant Expression and Purification of PPIases from Clinically Important Protists
5. Assays on the Activity of PPIases from Clinically Important Protists
6. Inhibition Assays of rPPIases from Protozoan Parasites
7. Biotechnological Applications of Protozoan PPIases
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, S.M.; Kim, M.S.; Hayat, F.; Shin, D. Recent advances in the discovery of novel antiprotozoal agents. Molecules 2019, 24, 3886. [Google Scholar] [CrossRef]
- Forrester, S.J.; Hall, N. The revolution of whole genome sequencing to study parasites. Mol. Biochem. Parasitol. 2014, 195, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Fischer, G. Peptidyl-Prolyl cis/trans Isomerases and Their Effectors. Angew. Chem. Int. Ed. Engl. 1994, 33, 1415–1436. [Google Scholar] [CrossRef]
- Fanghanel, J.; Fischer, G. Insights into the catalytic mechanism of peptidyl prolyl cis/trans isomerases. Front. Biosci. 2004, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.P.; Finn, G.; Lee, T.H.; Nicholson, L.K. Prolyl cis-trans isomerization as a molecular timer. Nat. Chem. Biol. 2007, 3, 619–629. [Google Scholar] [CrossRef]
- Bell, A.; Monaghan, P.; Page, A.P. Peptidyl-prolyl cis-trans isomerases (immunophilins) and their roles in parasite biochemistry, host-parasite interaction and antiparasitic drug action. Int. J. Parasitol. 2006, 36, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Pahlke, D.; Freund, C.; Leitner, D.; Labudde, D. Statistically significant dependence of the Xaa-Pro peptide bond conformation on secondary structure and amino acid sequence. BMC Struct. Biol. 2005, 5, 8. [Google Scholar] [CrossRef]
- Hennig, L.; Christner, C.; Kipping, M.; Schelbert, B.; Rücknagel, K.P.; Grabley, S.; Küllertz, G.; Fischer, G. Selective inactivation of parvulin-like peptidyl-prolyl cis/trans isomerases by juglone. Biochemistry 1998, 37, 5953–5960. [Google Scholar] [CrossRef] [PubMed]
- Jordens, J.; Janssens, V.; Longin, S.; Stevens, I.; Martens, E.; Bultynck, G.; Engelborghs, Y.; Lescrinier, E.; Waelkens, E.; Goris, J.; et al. The protein phosphatase 2A phosphatase activator is a novel peptidyl-prolyl cis/trans-isomerase. J. Biol. Chem. 2006, 281, 6349–6357. [Google Scholar] [CrossRef]
- Magnusdottir, A.; Stenmark, P.; Flodin, S.; Nyman, T.; Hammarström, M.; Ehn, M.; Bakali, H.M.A.; Berglund, H.; Nordlund, P. The crystal structure of a human PP2A phosphatase activator reveals a novel fold and highly conserved cleft implicated in protein-protein interactions. J. Biol. Chem. 2006, 281, 22434–22438. [Google Scholar] [CrossRef]
- Handschumacher, R.; Harding, M.; Rice, J.; Drugge, R.; Speicher, D. Cyclophilin: A specific cytosolic binding protein for Cyclosporin A. Science 1984, 226, 544–547. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Heitman, J. The cyclophilins. Genome Biol. 2005, 6, 226. [Google Scholar] [CrossRef]
- Galat, A. Peptidyl prolyl cis/trans isomerases (Immunophilins): Biological diversity-targets-functions. Curr. Top. Med. Chem. 2003, 3, 1315–1347. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.L.; Walker, J.R.; Campagna-Slater, V.; Finerty, P.J.; Finerty, P.J.; Paramanathan, R.; Bernstein, G.; Mackenzie, F.; Tempel, W.; Ouyang, H.; et al. Structural and biochemical characterization of the human cyclophilin family of peptidyl-prolyl isomerases. PLoS Biol. 2010, 8, e1000439. [Google Scholar] [CrossRef] [PubMed]
- Michnick, S.W.; Rosen, M.K.; Wandless, T.L.; Karplus, M.; Schreiber, S.L. Solution structure of FKBP, a rotamase enzyme and receptor for FK506 and rapamycin. Science 1991, 252, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Van Duyne, A.D.; Standaert, R.F.; Karplus, P.A.; Schreiber, S.L.; Clardy, J. Atomic structure of FKBP-FK506, an immunophilin-immunosuppressant complex. Science 1991, 252, 839–842. [Google Scholar] [CrossRef]
- Galat, A. Sequence diversification of the FK506-binding proteins in several different genomes. Eur. J. Biochem. 2000, 267, 4945–4959. [Google Scholar] [CrossRef] [PubMed]
- Rahfeld, J.-U.; Riicknagel, K.P.; Schelbert, B.; Ludwigb, B.; Hackerb, J.; Mannc, K.; Fischer, G. Confirmation of the existence of a third family among peptidyl-prolylcisltrans isomerases Amino acid sequence and recombinant production of parvulin. FEBS Lett. 1994, 352, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Rahfeld, J.-U.; Schierhornb, A.; Mannc, K.; Fischer, G. A novel peptidyl-prolylcisltrans isomerase from Escherichia coli. FEBS Lett. 1994, 343, 6569. [Google Scholar] [CrossRef]
- Rudd, K.E.; Sofia, H.J.; Koonin, E.V.; Plunkett, G.; Lazar, S.; Rouviere, P.E. A new family of peptidyl-prolyl isomerases. Trends Biochem. Sci. 1995, 20, 12–14. [Google Scholar] [CrossRef]
- Salah, Z.; Alian, A.; Aqeilan, R.I. WW domain-containing proteins: Retrospectives and the future. Front. Biosci.-Landmark 2012, 17, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, R.; Lu, K.P.; Hunter, T.; Noel, J.P. Structural and functional analysis of the mitotic rotamase Pin1 suggests substrate recognition is phosphorylation dependent. Cell 1997, 89, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Flaherty, P.T.; Jain, P. Peptidyl prolyl isomerase inhibitors. Annu. Rep. Med. Chem. 2011, 46, 337–349. [Google Scholar] [CrossRef]
- Trichomoniasis. STI treatment Guidelines. Available online: https://www.cdc.gov/std/treatment-guidelines/trichomoniasis.htm (accessed on 9 July 2024).
- Alvarez-Jarreta, J.; Amos, B.; Aurrecoechea, C.; Bah, S.; Barba, M.; Barreto, A.; Basenko, E.Y.; Belnap, R.; Blevins, A.; Böhme, U.; et al. VEuPathDB: The eukaryotic pathogen, vector and host bioinformatics resource center in 2023. Nucleic Acids Res. 2024, 52, D808–D816. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hsu, H.M.; Chu, C.H.; Wang, Y.T.; Lee, Y.; Wei, S.Y.; Liu, H.W.; Ong, S.J.; Chen, C.; Tai, J.H. Regulation of nuclear translocation of the Myb1 transcription factor by TvCyclophilin 1 in the protozoan parasite Trichomonas vaginalis. J. Biol. Chem. 2014, 289, 19120–19136. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, P.; Livstone, M.S.; Lewis, S.E.; Thomas, P.D. Phylogenetic-based propagation of functional annotations within the Gene Ontology consortium. Brief. Bioinform. 2011, 12, 449–462. [Google Scholar] [CrossRef]
- Hsu, H.M.; Huang, Y.H.; Aryal, S.; Liu, H.W.; Chen, C.; Chen, S.H.; Chu, C.H.; Tai, J.H. Endomembrane Protein Trafficking Regulated by a TvCyP2 Cyclophilin in the Protozoan Parasite, Trichomonas vaginalis. Sci. Rep. 2020, 10, 1275. [Google Scholar] [CrossRef]
- Aryal, S.; Hsu, H.M.; Lou, Y.C.; Chu, C.H.; Tai, J.H.; Hsu, C.H.; Chen, C. N-terminal segment of tvcyp2 cyclophilin from trichomonas vaginalis is involved in self-association, membrane interaction, and subcellular localization. Biomolecules 2020, 10, 1239. [Google Scholar] [CrossRef]
- Martin, T.; Lou, Y.C.; Chou, C.C.; Wei, S.Y.; Sadotra, S.; Cho, C.C.; Lin, M.H.; Tai, J.H.; Hsu, C.H.; Chen, C. Structural basis of interaction between dimeric cyclophilin 1 and Myb1 transcription factor in Trichomonas vaginalis. Sci. Rep. 2018, 8, 5410. [Google Scholar] [CrossRef] [PubMed]
- Graveley, B.R. A protein interaction domain contacts RNA in the prespliceosome. Mol. Cell 2004, 13, 302–304. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: The universal protein knowledge base in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.L.; Walker, J.R.; Ouyang, H.; MacKenzie, F.; Butler-Cole, C.; Newman, E.M.; Eisenmesser, E.Z.; Dhe-Paganon, S. The crystal structure of human WD40 repeat-containing peptidylprolyl isomerase (PPWD1). FEBS J. 2008, 275, 2283–2295. [Google Scholar] [CrossRef]
- Maris, C.; Dominguez, C.; Allain, F.H.T. The RNA recognition motif, a plastic RNA-binding platform to regulate post-transcriptional gene expression. FEBS J. 2005, 272, 2118–2131. [Google Scholar] [CrossRef]
- Hatakeyama, S.; Kei-ichi, I.N. U-box proteins as a new family of ubiquitin ligases. Biochem. Biophys. 2003, 302, 635–645. [Google Scholar] [CrossRef]
- Ohi, M.D.; vander Kooi, C.W.; Rosenberg, J.A.; Chazin, W.J.; Gould, K.L. Structural insights into the U-box, a domain associated with multi-ubiquitination. Nat. Struct. Biol. 2003, 10, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Richly, H.; Rape, M.; Braun, S.; Rumpf, S.; Hoege, C.; Jentsch, S. A series of ubiquitin binding factors connects CDC48/p97 to substrate multiubiquitylation and proteasomal targeting. Cell 2005, 120, 73–84. [Google Scholar] [CrossRef]
- Xu, C.; Min, J. Structure and function of WD40 domain proteins. In Protein and Cell; Higher Education Press Limited Company: Beijing, China, 2011; Volume 2, pp. 202–214. [Google Scholar] [CrossRef]
- D’Andrea, L.D.; Regan, L. TPR proteins: The versatile helix. In Trends in Biochemical Sciences; Elsevier Ltd.: Amsterdam, The Netherlands, 2003; Volume 28, pp. 655–662. [Google Scholar] [CrossRef]
- Zawadzka, K.; Zawadzki, P.; Baker, R.; Rajasekar, K.V.; Wagner, F.; Sherratt, D.J.; Arciszewska, L.K. MukB ATPases are regulated independently by the N-and C-terminal domains of MukFkleisin. eLife 2018, 7, e31522. [Google Scholar] [CrossRef]
- Audibert, A.A.; Simonelig, M. Autoregulation at the level of mRNA 3 end formation of the suppressor of forked gene of Drosophila melanogaster is conserved in Drosophila virilis. Proc. Natl. Acad. Sci. USA 1998, 95, 14302–14307. [Google Scholar] [CrossRef]
- Morán, P.; Serrano-Vázquez, A.; Rojas-Velázquez, L.; González, E.; Pérez-Juárez, H.; Hernández, E.G.; Padilla, M.d.L.A.; Zaragoza, M.E.; Portillo-Bobadilla, T.; Ramiro, M.; et al. Amoebiasis: Advances in Diagnosis, Treatment, Immunology Features and the Interaction with the Intestinal Ecosystem. Int. J. Mol. Sci. 2023, 24, 11755. [Google Scholar] [CrossRef] [PubMed]
- Ostoa-Saloma, P.; Carrero, J.C.; Petrossian, P.; Hérió, P.; Landa, A.; Laclette, J.P. Cloning, characterization and functional expression of a cyclophilin of Entamoeba histolytica. Mol. Biochem. Parasitol. 2000, 107, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Matena, A.; Rehic, E.; Hönig, D.; Kamba, B.; Bayer, P. Structure and function of the human parvulins Pin1 and Par14/17. Biol. Chem. 2018, 399, 101–125. [Google Scholar] [CrossRef] [PubMed]
- Adam, R.D. Giardia duodenalis: Biology and pathogenesis. Clin. Microbiol. Rev. 2021, 34, e0002419. [Google Scholar] [CrossRef] [PubMed]
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium join the “Neglected Diseases Initiative”. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fevre, E.M.; Sripa, B.; et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef]
- European Centre for Disease Prevention and Control. Giardiasis (lambliasis). 2022. In Annual Epidemiological. Report for 2019; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Giardiasis NNDSS Summary Report for 2019. Waterborne Disease and Outbreak Surveillance Reporting. Available online: https://www.cdc.gov/healthy-water-data/documentation/giardiasis-nndss-summary-report-for-2019.html#:~:text=In%202019%2C%20there%20were%2014%2C887,confirmed%20and%203.4%25%20nonconfirmed (accessed on 22 May 2024).
- Franzén, O.; Jerlström-Hultqvist, J.; Castro, E.; Sherwood, E.; Ankarklev, J.; Reiner, D.S.; Palm, D.; Andersson, J.O.; Andersson, B.; Svärd, S.G. Draft genome sequencing of Giardia intestinalis assemblage B isolate GS: Is human giardiasis caused by two different species? PLoS Pathog. 2009, 5, e1000560. [Google Scholar] [CrossRef]
- Ankarklev, J.; Franzén, O.; Peirasmaki, D.; Jerlström-Hultqvist, J.; Lebbad, M.; Andersson, J.; Andersson, B.; Svärd, S.G. Comparative genomic analyses of freshly isolated Giardia intestinalis assemblage A isolates. BMC Genom. 2015, 16, 697. [Google Scholar] [CrossRef]
- Zajaczkowski, P.; Lee, R.; Fletcher-Lartey, S.M.; Alexander, K.; Mahimbo, A.; Stark, D.; Ellis, J.T. The controversies surrounding Giardia intestinalis assemblages A and B. Curr. Res. Parasitol. Vector-Borne Dis. 2021, 1, 100055. [Google Scholar] [CrossRef]
- Morrison, H.G.; McArthur, A.G.; Gillin, F.D.; Aley, S.B.; Adam, R.D.; Olsen, G.J.; Best, A.A.; Cande, W.Z.; Chen, F.; Cipriano, M.J.; et al. Genomic minimalism in the early diverging intestinal parasite Giardia lamblia. Science 2007, 317, 1921–1926. [Google Scholar] [CrossRef]
- Adam, R.D.; Dahlstrom, E.W.; Martens, C.A.; Bruno, D.P.; Barbian, K.D.; Ricklefs, S.M.; Hernandez, M.M.; Narla, N.P.; Patel, R.B.; Porcella, S.F.; et al. Genome sequencing of Giardia lamblia genotypes A2 and B isolates (DH and GS) and comparative analysis with the genomes of Genotypes A1 and E (WB and pig). Genome Biol. Evol. 2013, 5, 2498–2511. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef] [PubMed]
- Ma’ayeh, S.Y.; Liu, J.; Peirasmaki, D.; Hörnaeus, K.; Bergström Lind, S.; Grabherr, M.; Bergquist, J.; Svärd, S.G. Characterization of the Giardia intestinalis secretome during interaction with human intestinal epithelial cells: The impact on host cells. PLoS Neglected Trop. Dis. 2017, 11, e0006120. [Google Scholar] [CrossRef]
- Liu, L.; Yang, Y.; Fang, R.; Zhu, W.; Wu, J.; Li, X.; Patankar, J.V.; Li, W. Giardia duodenalis and its secreted ppib trigger inflammasome activation and pyroptosis in macrophages through TLR4-induced ROS signaling and A20-mediated NLRP3 deubiquitination. Cells 2021, 10, 3425. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The Protein Data Bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.G.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-S.; Kong, H.-H.; Chung, D.-I. Cloning and characterization of Giardia intestinalis cyclophilin. Korean J. Parasitol. 2002, 40, 131–138. [Google Scholar] [CrossRef]
- Galat, A. Peptidylprolyl isomerases as in vivo carriers for drugs that target various intracellular entities. Biomolecules 2017, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Buchko, G.W.; Hewitt, S.N.; Van Voorhis, W.C.; Myler, P.J. Solution structure of a putative FKBP-type peptidyl-propyl cis-trans isomerase from Giardia lamblia. J. Biomol. NMR 2013, 57, 369–374. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Trypanosomiasis, Human African (Sleeping Sickness). 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 24 August 2023).
- World Health Organization. Chagas Disease (Also Known as American Trypanosomiasis). 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 24 August 2023).
- Medeiros, C.A.; Silva, M.B.A.; Oliveira, A.L.S.; Alves, S.M.M.; Barros, M.D.N.D.D.S.; Cavalcanti, M.D.G.A.M.; Oliveira, G.M.A.; Carrazzone, C.F.V.; Oliveira, W.A., Jr.; Medeiros, Z.M. Mapping the morbidity and mortality of Chagas disease in an endemic area in Brazil. Rev. Inst. Med. Trop. Sao Paulo 2022, 64, e5. [Google Scholar] [CrossRef] [PubMed]
- Papagni, R.; Novara, R.; Minardi, M.L.; Frallonardo, L.; Panico, G.G.; Pallara, E.; Cotugno, S.; Ascoli Bartoli, T.; Guido, G.; De Vita, E.; et al. Human African Trypanosomiasis (sleeping sickness): Current knowledge and future challenges. Front. Trop. Dis. 2023, 4, 1087003. [Google Scholar] [CrossRef]
- El-Sayed, N.M.; Myler, P.J.; Blandin, G.; Berriman, M.; Crabtree, J.; Aggarwal, G.; Caler, E.; Renauld, H.; Worthey, E.A.; Hertz-Fowler, C.; et al. Comparative genomics of trypanosomatid parasitic protozoa. Science 2005, 309, 404–409. [Google Scholar] [CrossRef] [PubMed]
- Potenza, M.; Galat, A.; Minning, T.A.; Ruiz, A.M.; Duran, R.; Tarleton, R.L.; Marín, M.; Fichera, L.E.; Búa, J. Analysis of the Trypanosoma cruzi cyclophilin gene family and identification of Cyclosporin A binding proteins. Parasitology 2006, 132, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.P.; Sanders, M.; Berry, A.; McQuillan, J.; Aslett, M.A.; Quail, M.A.; Chukualim, B.; Capewell, P.; Macleod, A.; Melville, S.E.; et al. The genome sequence of Trypanosoma brucei gambiense, causative agent of chronic human African Trypanosomiasis. PLoS Negl. Trop. Dis. 2010, 4, e658. [Google Scholar] [CrossRef]
- Búa, J.; Aslund, L.; Pereyra, N.; García, G.A.; Bontempi, E.J.; Ruiz, A.M. Characterisation of a cyclophilin isoform in Trypanosoma cruzi. FEMS Microbiol. Lett. 2001, 200, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.P.; Abukawa, F.M.; Souza-Melo, N.; Alcântara, L.M.; Bittencourt-Cunha, P.; Moraes, C.B.; Jha, B.K.; McGwire, B.S.; Moretti, N.S.; Schenkman, S. Cyclophilin 19 secreted in the host cell cytosol by Trypanosoma cruzi promotes ROS production required for parasite growth. Cell. Microbiol. 2020, 23, e13295. [Google Scholar] [CrossRef]
- Bustos, P.L.; Volta, B.J.; Perrone, A.E.; Milduberger, N.; Búa, J. A homolog of cyclophilin D is expressed in Trypanosoma cruzi and is involved in the oxidative stress–damage response. Cell Death Discov. 2017, 3, 16092. [Google Scholar] [CrossRef]
- Búa, J.; Fichera, L.E.; Fuchs, A.G.; Potenza, M.; Dubin, M.; Wenger, R.O.; Moretti, G.; Scabone, C.M.; Ruiz, A.M. Anti-Trypanosoma cruzi effects of cyclosporin A derivatives: Possible role of a P-glycoprotein and parasite cyclophilins. Parasitology 2008, 135, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Moro, A.; Ruiz-Cabello, F.; Fernández-Cano, A.; Stock, R.P.; Gonzalez, A. Secretion by Trypanosoma cruzi of a peptidyl-prolyl cis-trans isomerase involved in cell infection. EMBO J. 1995, 14, 2483–2490. [Google Scholar] [CrossRef] [PubMed]
- Truebestein, L.; Leonard, T.A. Coiled-coils: The long and short of it. BioEssays 2016, 38, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Bayer-Santos, E.; Aguilar-Bonavides, C.; Rodrigues, S.P.; Cordero, E.M.; Marques, A.F.; Varela-Ramirez, A.; Choi, H.; Yoshida, N.; Da Silveira, J.F.; Almeida, I.C. Proteomic analysis of Trypanosoma cruzi secretome: Characterization of two populations of extracellular vesicles and soluble proteins. J. Proteome Res. 2013, 12, 883–897. [Google Scholar] [CrossRef] [PubMed]
- Erben, E.D.; Daum, S.; Téllez-Iñón, M.T. The Trypanosoma cruzi PIN1 gene encodes a parvulin peptidyl-prolyl cis/trans isomerase able to replace the essential ESS1 in Saccharomyces cerevisiae. Mol. Biochem. Parasitol. 2007, 153, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Erben, E.D.; Valguarnera, E.; Nardelli, S.; Chung, J.; Daum, S.; Potenza, M.; Schenkman, S.; Téllez-Iñón, M.T. Identification of an atypical peptidyl-prolyl cis/trans isomerase from trypanosomatids. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Durocher, D.; Jackson, S.P. The FHA domain. FEBS Lett. 2002, 513, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Erben, E.D.; Nardelli, S.C.; De Jesus, T.C.L.; Schenkman, S.; Tellez-Iñon, M.T. Trypanosomatid pin1-type peptidyl-prolyl isomerase is cytosolic and not essential for cell proliferation. J. Eukaryot. Microbiol. 2013, 60, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Pellé, R.; McOdimba, F.; Chuma, F.; Wasawo, D.; Pearson, T.W.; Murphy, N.B. The African trypanosome cyclophilin A homologue contains unusual conserved central and N-terminal domains and is developmentally regulated. Gene 2002, 290, 181–191. [Google Scholar] [CrossRef]
- Geiger, A.; Hirtz, C.; Bécue, T.; Bellard, E.; Centeno, D.; Gargani, D.; Rossignol, M.; Cuny, G.; Peltier, J.-B. Exocytosis and protein secretion in Trypanosoma. BMC Microbiol. 2010, 10, 20. [Google Scholar] [CrossRef]
- Brasseur, A.; Rotureau, B.; Vermeersch, M.; Blisnick, T.; Salmon, D.; Bastin, P.; Pays, E.; Vanhamme, L.; Pérez-Morgaa, D. Trypanosoma brucei FKBP12 differentially controls motility and cytokinesis in procyclic and bloodstream forms. Eukaryot. Cell 2013, 12, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Desjeux, P. Leishmaniasis: Current situation and new perspectives. Comp. Immunol. Microbiol. Infect. Dis. 2004, 27, 305–318. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Leishmaniasis. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 1 May 2023).
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Ivens, A.C.; Peacock, C.S.; Worthey, E.A.; Murphy, L.; Aggarwal, G.; Berriman, M.; Sisk, E.; Rajandream, M.-A.; Adlem, E.; Aert, R.; et al. The genome of the kinetoplastid parasite, Leishmania major. Science 2005, 309, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Downing, T.; Imamura, H.; Decuypere, S.; Clark, T.G.; Coombs, G.H.; Cotton, J.A.; Hilley, J.D.; De Doncker, S.; Maes, I.; Mottram, J.C.; et al. Whole genome sequencing of multiple Leishmania donovani clinical isolates provides insights into population structure and mechanisms of drug resistance. Genome Res. 2011, 21, 2143–2156. [Google Scholar] [CrossRef] [PubMed]
- Soufari, H.; Waltz, F.; Parrot, C.; Durrieu-Gaillard, S.; Bochler, A.; Kuhn, L.; Sissler, M.; Hashem, Y.; Protéomique, P. Structure of the mature kinetoplastids mitoribosome and insights into its large subunit biogenesis. Proc. Natl. Acad. Sci. USA 2020, 117, 29851–29861. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, V.; Xue, Z.; Sherry, B.; Bukrinsky, M. Functional analysis of Leishmania major cyclophilin. Int. J. Parasitol. 2008, 38, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Dutta, M.; Delhi, P.; Sinha, K.M.; Banerjee, R.; Datta, A.K. Lack of abundance of cytoplasmic cyclosporin A-binding protein renders free-living Leishmania donovani resistant to Cyclosporin A. J. Biol. Chem. 2001, 276, 19294–19300. [Google Scholar] [CrossRef]
- Venugopal, V.; Sen, B.; Datta, A.K.; Banerjee, R. Structure of cyclophilin from Leishmania donovani at 1.97 Å resolution. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2007, 63, 60–64. [Google Scholar] [CrossRef]
- Rascher, C.; Pahl, A.; Pecht, A.; Brune, K.; Solbach, W.; Bang, H. Leishmania major parasites express cyclophilin isoforms with an unusual interaction with calcineurin. Biochem. J 1998, 334, 659–667. [Google Scholar] [CrossRef]
- Hoerauf, A.; Rascher, C.; Bang, R.; Pahl, A.; Solbach, W.; Brune, K.; Röllinghoff, M.; Bang, H. Host-cell cyclophilin is important for the intracellular replication of Leishmania major. Mol. Microbiol. 1997, 24, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Yau, W.L.; Blisnick, T.; Taly, J.F.; Helmer-Citterich, M.; Schiene-Fischer, C.; Leclercq, O.; Li, J.; Schmidt-Arras, D.; Morales, M.A.; Notredame, C.; et al. Cyclosporin A treatment of Leishmania donovani reveals stage-specific functions of cyclophilins in parasite proliferation and viability. PLoS Neglected Trop. Dis. 2010, 4, e729. [Google Scholar] [CrossRef] [PubMed]
- Biswas, G.; Ghosh, S.; Raghuraman, H.; Banerjee, R. Probing conformational transitions of PIN1 from L. major during chemical and thermal denaturation. Int. J. Biol. Macromol. 2020, 154, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Das, I.; Datta, R.; Sen, B.; Bhattacharyya, D.; Mandal, C.; Datta, A.K. A single-domain cyclophilin from Leishmania donovani reactivates soluble aggregates of adenosine kinase by isomerase-independent chaperone function. J. Biol. Chem. 2002, 277, 47451–47460. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Basu, S.; Datta, A.K.; Bhattacharyya, D.; Banerjee, R.; Dasgupta, D. Equilibrium unfolding of cyclophilin from Leishmania donovani: Characterization of intermediate states. Int. J. Biol. Macromol. 2014, 69, 353–360. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Malaria. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/malaria (accessed on 24 August 2023).
- Auburn, S.; Böhme, U.; Steinbiss, S.; Trimarsanto, H.; Hostetler, J.; Sanders, M.; Gao, Q.; Nosten, F.; Newbold, C.I.; Berriman, M.; et al. A new Plasmodium vivax reference sequence with improved assembly of the subtelomeres reveals an abundance of pir genes. Wellcome Open Res. 2016, 1, 4. [Google Scholar] [CrossRef]
- Neafsey, D.E.; Galinsky, K.; Jiang, R.H.Y.; Young, L.; Sykes, S.M.; Saif, S.; Gujja, S.; Goldberg, J.M.; Young, S.; Zeng, Q.; et al. The malaria parasite Plasmodium vivax exhibits greater genetic diversity than Plasmodium falciparum. Nat. Genet. 2012, 44, 1046–1050. [Google Scholar] [CrossRef] [PubMed]
- Carlton, J.M.; Das, A.; Escalante, A.A. Genomics, population genetics and evolutionary history of Plasmodium vivax. Adv. Parasitol. 2013, 81, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Marín-Menéndez, A.; Bell, A. Overexpression, purification and assessment of cyclosporin binding of a family of cyclophilins and cyclophilin-like proteins of the human malarial parasite Plasmodium falciparum. Protein Expr. Purif. 2011, 78, 225–234. [Google Scholar] [CrossRef]
- Vedadi, M.; Lew, J.; Artz, J.; Amani, M.; Zhao, Y.; Dong, A.; Wasney, G.A.; Gao, M.; Hills, T.; Brokx, S.; et al. Genome-scale protein expression and structural biology of Plasmodium falciparum and related Apicomplexan organisms. Mol. Biochem. Parasitol. 2007, 151, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.R.; Hall, D.R.; Berriman, M.; Nunes, J.A.; Leonard, G.A.; Fairlamb, A.H.; Hunter, W.N. The three-dimensional structure of a Plasmodium falciparum cyclophilin in complex with the potent anti-malarial cyclosporin A. J. Mol. Biol. 2000, 298, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.R.; Kang, C.B.; Chia, J.; Tang, K.; Yoon, H.S. Expression, purification, and molecular characterization of Plasmodium falciparum FK506-binding protein 35 (PfFKBP35). Protein Expr. Purif. 2007, 53, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Adams, B.; Musiyenko, A.; Shulyayeva, O.; Barik, S. The FK506-binding protein of the malaria parasite, Plasmodium falciparum, is a FK506-sensitive chaperone with FK506-independent calcineurin-inhibitory activity. Mol. Biochem. Parasitol. 2005, 141, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Berriman, M.; Fairlamb, A.H. Detailed characterization of a cyclophilin from the human malaria parasite Plasmodium falciparum. Biochem. J. 1998, 334, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Gavigan, C.S.; Kiely, S.P.; Hirtzlin, J.; Bell, A. Cyclosporin-binding proteins of Plasmodium falciparum. Int. J. Parasitol. 2003, 33, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Hirtzlin, J.; Farber, P.M.; Franklin, R.M.; Bell, A. Molecular and biochemical characterization of a Plasmodium falciparum cyclophilin containing a cleavable signal sequence. Eur. J. Biochem. 1995, 232, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Cong, B.K.; Ye, H.; Hye, R.Y.; Ho, S.Y. Solution structure of FK506 binding domain (FKBD) of Plasmodium falciparum FK506 binding protein 35 (PfFKBP35). Proteins Struct. Funct. Genet. 2008, 70, 300–302. [Google Scholar] [CrossRef]
- Wu, Y.; Craig, A. Comparative proteomic analysis of metabolically labelled proteins from Plasmodium falciparum isolates with different adhesion properties. Malar. J. 2006, 5, 67. [Google Scholar] [CrossRef]
- Rajan, S.; Yoon, H.S. Structural insights into Plasmodium PPIases. Front. Cell. Infect. Microbiol. 2022, 12, 931635. [Google Scholar] [CrossRef]
- Marín-Menéndez, A.; Monaghan, P.; Bell, A. A family of cyclophilin-like molecular chaperones in Plasmodium falciparum. Mol. Biochem. Parasitol. 2012, 184, 44–47. [Google Scholar] [CrossRef]
- Reddy, G.R. Cloning and characterization of a Plasmodium falciparum cyclophilin gene that is stage-specifically expressed. Mol. Biochem. Parasitol. 1995, 73, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Alag, R.; Shin, J.; Yoon, H.S. NMR assignments of the FK506-binding domain of FK506-binding protein 35 from Plasmodium vivax. Biomol. NMR Assign. 2009, 3, 243–245. [Google Scholar] [CrossRef] [PubMed]
- Rajan, S.; Austin, D.; Harikishore, A.; Nguyen, Q.T.; Baek, K.; Yoon, H.S. Crystal structure of Plasmodium vivax FK506-binding protein 25 reveals conformational changes responsible for its noncanonical activity. Proteins Struct. Funct. Bioinform. 2014, 82, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Alag, R.; Qureshi, I.A.; Bharatham, N.; Shin, J.; Lescar, J.; Yoon, H.S. NMR and crystallographic structures of the FK506 binding domain of human malarial parasite Plasmodium vivax FKBP35. Protein Sci. 2010, 19, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Toxoplasmosis. 2023. Available online: https://www.cdc.gov/dpdx/toxoplasmosis/index.html (accessed on 23 August 2023).
- About Toxoplasmosis. Toxoplasmosis. Available online: https://www.cdc.gov/toxoplasmosis/about/index.html (accessed on 22 April 2024).
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. World Health Organ. 2013, 91, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Congenital Toxoplasmosis—Annual Epidemiological Report for 2020. European Centre for Disease Prevention and Control. Available online: https://www.ecdc.europa.eu/en/publications-data/congenital-toxoplasmosis-annual-epidemiological-report-2020 (accessed on 29 August 2023).
- Kim, K.; Weiss, L.M. Toxoplasma: The next 100 years. Microbes Infect. 2008, 10, 978–984. [Google Scholar] [CrossRef] [PubMed]
- Langosch, D.; Arkin, I.T. Interaction and conformational dynamics of membrane-spanning protein helices. Protein Sci. 2009, 18, 1343–1358. [Google Scholar] [CrossRef] [PubMed]
- Adams, B.; Musiyenko, A.; Kumar, R.; Barik, S. A novel class of dual-family immunophilins. J. Biol. Chem. 2005, 280, 24308–24314. [Google Scholar] [CrossRef] [PubMed]
- Aliberti, J.; Valenzuela, J.G.; Carruthers, V.B.; Hieny, S.; Andersen, J.; Charest, H.; Sousa, C.R.; Fairlamb, A.; Ribeiro, J.M.; Sher, A. Molecular mimicry of a CCR5 binding-domain in the microbial activation of dendritic cells. NaturenImmunology 2003, 4, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Favretto, F.; Jiménez-Faraco, E.; Conter, C.; Dominici, P.; Hermoso, J.A.; Astegno, A. Structural basis for cyclosporin isoform-specific inhibition of cyclophilins from Toxoplasma gondii. ACS Infect. Dis. 2023, 9, 365–377. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Bannai, H.; Xuan, X.; Nishikawa, Y. Toxoplasma gondii cyclophilin 18-mediated production of nitric oxide induces bradyzoite conversion in a CCR5-dependent manner. Infect. Immun. 2009, 77, 3686–3695. [Google Scholar] [CrossRef] [PubMed]
- High, K.P.; Joiner, K.A.; Handschumacher, R.E. Isolation, cDNA sequences, and biochemical characterization of the major cyclosporin-binding proteins of Toxoplasma gondii. J. Biochem. Chem. 1994, 269, 9105–9112. [Google Scholar] [CrossRef]
- Cryptosporidiosis|CDC Yellow Book 2024. Available online: https://wwwnc.cdc.gov/travel/yellowbook/2024/infections-diseases/cryptosporidiosis (accessed on 9 July 2024).
- Gilbert, I.H.; Vinayak, S.; Striepen, B.; Manjunatha, U.H.; Khalil, I.A.; Van Voorhis, W.C.; Cryptosporidiosis Therapeutics Advocacy Group CTAG; Cryptosporidiosis Therapeutics Advocacy CTAG. Safe and effective treatments are needed for cryptosporidiosis, a truly neglected tropical disease. BMJ Glob. Health 2023, 8, e012540. [Google Scholar] [CrossRef] [PubMed]
- Sow, S.O.; Muhsen, K.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.H.; Panchalingam, S.; Sur, D.; Zaidi, A.K.M.; Faruque, A.S.G. The burden of Cryptosporidium Diarrheal disease among children < 24 months of age in moderate/high mortality regions of sub-Saharan Africa and South Asia, utilizing data from the global Enteric multicenter study (GEMS). PLoS Negl. Trop. Dis. 2016, 10, e0004729. [Google Scholar] [CrossRef]
- Krücken, J.; Greif, G.; von Samson-Himmelstjerna, G. In silico analysis of the cyclophilin repertoire of apicomplexan parasites. Parasites Vectors 2009, 2, 27. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Dubourg, A.; Xia, D.; Winpenny, J.P.; Al Naimi, S.; Bouzid, M.; Sexton, D.W.; Wastling, J.M.; Hunter, P.R.; Tyler, K.M. Giardia secretome highlights secreted tenascins as a key component of pathogenesis. GigaScience 2018, 7, giy003. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, M.M.; Karafova, A.; Kamysz, W.; Schenkman, S.; Pelle, R.; McGwire, B.S. Secreted trypanosome cyclophilin inactivates lytic insect defense peptides and induces parasite calcineurin activation and infectivity. J. Biol. Chem. 2013, 288, 8772–8784. [Google Scholar] [CrossRef] [PubMed]
- Rêgo, J.V.; Duarte, A.P.; Liarte, D.B.; de Carvalho Sousa, F.; Barreto, H.M.; Búa, J.; Romanha, A.J.; Rádis-Baptista, G.; Murta, S.M.F. Molecular characterization of cyclophilin (TcCyP19) in Trypanosoma cruzi populations susceptible and resistant to benznidazole. Exp. Parasitol. 2015, 148, 73–80. [Google Scholar] [CrossRef]
- Jha, B.K.; Varikuti, S.; Verma, C.; Shivahare, R.; Bishop, N.; Dos Santos, G.P.; McDonald, J.; Sur, A.; Myler, P.J.; Schenkman, S.; et al. Immunization with a Trypanosoma cruzi cyclophilin-19 deletion mutant protects against acute Chagas disease in mice. Npj Vaccines 2023, 8, 63. [Google Scholar] [CrossRef]
- Perrone, A.E.; Pinillo, M.; Rial, M.S.; Fernández, M.; Milduberger, N.; González, C.; Bustos, P.L.; Fichera, L.E.; Laucella, S.A.; Albareda, M.C.; et al. Trypanosoma cruzi secreted cyclophilin TcCyP19 as an early marker for trypanocidal treatment efficiency. Int. J. Mol. Sci. 2023, 24, 11875. [Google Scholar] [CrossRef]
- Bustos, P.L.; Perrone, A.E.; Milduberger, N.A.; Búa, J. Improved immuno-detection of a low-abundance cyclophilin allows the confirmation of its expression in a protozoan parasite. Immunochem. Immunopathol. 2015, 1, 103. [Google Scholar] [CrossRef]
- Stoller, G.; Rücknagel, K.P.; Nierhaus, K.H.; Schmid, F.X.; Fischer, G.; Rahfeld, J.U. A ribosome-associated peptidyl-prolyl cis/trans isomerase identified as the trigger factor. EMBO J. 1995, 14, 4939–4948. [Google Scholar] [CrossRef] [PubMed]
- Goh, J.Y.; Lai, C.Y.; Tan, L.C.; Yang, D.; He, C.Y.; Liou, Y.C. Functional characterization of two novel parvulins in Trypanosoma brucei. FEBS Lett. 2010, 584, 2901–2908. [Google Scholar] [CrossRef] [PubMed]
- Ludlam, A.V.; Moore, B.A.; Xu, Z. The crystal structure of ribosomal chaperone trigger factor from Vibrio cholerae. Proc. Natl. Acad. Sci. USA 2004, 101, 13436–13441. [Google Scholar] [CrossRef]
- Sen, B.; Venugopal, V.; Chakraborty, A.; Datta, R.; Dolai, S.; Banerjee, R.; Datta, A.K. Amino acid residues of Leishmania donovani cyclophilin key to interaction with its adenosine kinase: Biological implications. Biochemistry 2007, 46, 7832–7843. [Google Scholar] [CrossRef]
- Chakraborty, A.; Sen, B.; Datta, R.; Datta, A.K. Isomerase-independent chaperone function of cyclophilin ensures aggregation prevention of adenosine kinase both in vitro and under in vivo conditions. Biochemistry 2004, 43, 11862–11872. [Google Scholar] [CrossRef]
- Morales, M.A.; Watanabe, R.; Laurent, C.; Lenormand, P.; Rousselle, J.C.; Namane, A.; Späth, G.F. Phosphoproteomic analysis of Leishmania donovani pro- and amastigote stages. Proteomics 2008, 8, 350–363. [Google Scholar] [CrossRef]
- Morales, M.A.; Watanabe, R.; Dacher, M.; Chafey, P.; Osorio, Y.; Fortéa, J.; Scott, D.A.; Beverley, S.M.; Ommen, G.; Clos, J.; et al. Phosphoproteome dynamics reveal heat-shock protein complexes specific to the Leishmania donovani infectious stage. Proc. Natl. Acad. Sci. USA 2010, 107, 8381–8386. [Google Scholar] [CrossRef]
- Leneghan, D.; Bell, A. Immunophilin-protein interactions in Plasmodium falciparum. Parasitology 2015, 142, 1404–1414. [Google Scholar] [CrossRef]
- Yarovinsky, F.; Andersen, J.F.; King, L.R.; Caspar, P.; Aliberti, J.; Golding, H.; Sher, A. Structural determinants of the anti-HIV activity of a CCR5 antagonist derived from Toxoplasma gondii. J. Biol. Chem. 2004, 279, 53635–53642. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, D.S.; Lee, E.J.; Mabon, S.A.; Misteli, T. A cyclophilin functions in pre-mRNA splicing. EMBO J. 2002, 21, 470–480. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fernández-Robledo, J.A.; Vasta, G.R. Production of recombinant proteins from protozoan parasites. Trends Parasitol. 2010, 26, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Guerra, Á.P.; Calvo, E.P.; Wasserman, M.; Chaparro-Olaya, J. Production of recombinant proteins from Plasmodium falciparum in Escherichia coli. Biomedica 2016, 36, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.J.; Vega, M.C.; González-Rey, E.; Fernández-Carazo, R.; Macedo-Ribeiro, S.; Gomis-Rüth, F.X.; González, A.; Coll, M. Trypanosoma cruzi macrophage infectivity potentiator has a rotamase core and a highly exposed alpha-helix. EMBO Rep. 2002, 3, 88–94. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Monaghan, P.; Bell, A. A Plasmodium falciparum FK506-binding protein (FKBP) with peptidyl-prolyl cis-trans isomerase and chaperone activities. Mol. Biochem. Parasitol. 2005, 139, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Harikishore, A.; Niang, M.; Rajan, S.; Preiser, P.R.; Yoon, H.S. Small molecule plasmodium FKBP35 inhibitor as a potential antimalaria agent. Sci. Rep. 2013, 3, srep02501. [Google Scholar] [CrossRef]
- Fischer, G.; Bang, H.; Mech, C. NachweiseinerEnzymkatalyse für die cis-trans-Isomerisierung der Peptidbindung in prolinhaltigenPeptiden. Biomed Biochim Acta 1984, 43, 1101–1111. [Google Scholar] [PubMed]
- Kofron, J.L.; Kuzmic, P.; Kishore, V.; Colón-Bonilla, E.; Rich, D.H. Determination of kinetic constants for peptidyl prolyl cis-trans isomerase by an improved spectrophotometric assay. Biochemistry 1991, 30, 6127–6134. [Google Scholar] [CrossRef]
- Harrison, R.K.; Stein, R.L. Substrate specificities of the peptidyl prolyl cis-trans isomerase activities of distinct enzymes cyclophilin and FK-506 binding protein: Evidence for the existence of a family of distinct enzymes. Biochemistry 1990, 29, 3813–3816. [Google Scholar] [CrossRef]
- Siekierka, J.; Hung, S.; Poe, M.; Lin, C.; Sigal, N. A cytosolic binding protein for the immunosuppressant FK506 has peptidyl-prolyl isomerase activity but is distinct from cyclophilin. Lett. Nat. 1989, 341, 755–757. [Google Scholar] [CrossRef]
- Ünal, C.M.; Steinert, M. Microbial peptidyl-prolyl cis/trans isomerases (PPIases): Virulence factors and potential alternative drug targets. Microbiol. Mol. Biol. Rev. 2014, 78, 544–571. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, K.; Kakalis, L.T.; Anderson, K.S.; Armitage, I.M.; Kandschumacher, R.E.; Handschumacher, R.E. Expression of human cyclophilin-40 and the effect of the His141+Trp mutation on catalysis and cyclosporin A binding. Eur. J. Biochem. 1995, 229, 188–193. [Google Scholar] [CrossRef]
- Connern, C.P.; Halestrap, A.P. Purification and N-terminal sequencing of peptidyl-prolyl cis-trans-isomerase from rat liver mitochondrial matrix reveals the existence of a distinct mitochondrial cyclophilin. Biochem. J. 1992, 284, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Dunyak, B.M.; Gestwicki, J.E. Peptidyl-proline isomerases (PPIases): Targets for natural products and natural product-inspired compounds. J. Med. Chem. 2016, 59, 9622–9644. [Google Scholar] [CrossRef] [PubMed]
- Golding, H.; Aliberti, J.; King, L.R.; Manischewitz, J.; Andersen, J.; Valenzuela, J.; Landau, N.R.; Sher, A. Inhibition of HIV-1 infection by a CCR5-binding cyclophilin from Toxoplasma gondii. Blood 2003, 102, 3280–3286. [Google Scholar] [CrossRef] [PubMed]
- Golding, H.; Khurana, S.; Yarovinsky, F.; King, L.R.; Abdoulaeva, G.; Antonsson, L.; Owman, C.; Platt, E.J.; Kabat, D.; Andersen, J.F.; et al. CCR5 N-terminal region plays a critical role in HIV-1 inhibition by Toxoplasma gondii-derived cyclophilin-18. J. Biol. Chem. 2005, 280, 29570–29577. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Huang, X.; Yu, Q.; Li, Y.; Huang, J.; Li, J.; Yang, J.; Li, H.; Zhang, G.; Ren, W.; et al. The protective effect of a DNA vaccine encoding the Toxoplasma gondii cyclophilin gene in BALB/c mice. Parasite Immunol. 2013, 35, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Huang, X.; Gong, P.; Zhang, Q.; Li, J.; Zhang, G.; Yang, J.; Li, H.; Wang, N.; Zhang, X. Protective immunity induced by a recombinant BCG vaccine encoding the cyclophilin gene of Toxoplasma gondii. Vaccine 2013, 31, 6065–6071. [Google Scholar] [CrossRef] [PubMed]
- Perkins, M.E.; Wu, T.W.; Le Blancq, S.M. Cyclosporin analogs inhibit in vitro growth of Cryptosporidium parvum. Antimicrob. Agents Chemother. 1998, 42, 843–848. [Google Scholar] [CrossRef]
- Yang, H.-P.; Zhong, H.-N.; Zhou, H.-M. Catalysis of the refolding of urea denatured creatine kinase by peptidyl-prolyl cis-trans isomerase. Biochim. Et Biophys. Acta 1997, 1338, 147–150. [Google Scholar] [CrossRef] [PubMed]
- de la Cruz, J.J.; Villanueva-Lizama, L.; Dzul-Huchim, V.; Ramírez-Sierra, M.-J.; Martinez-Vega, P.; Rosado-Vallado, M.; Ortega-Lopez, J.; Flores-Pucheta, C.I.; Gillespie, P.; Zhan, B.; et al. Production of recombinant TSA-1 and evaluation of its potential for the immuno-therapeutic control of Trypanosoma cruzi infection in mice. Hum. Vaccines Immunother. 2018, 15, 210–219. [Google Scholar] [CrossRef] [PubMed]






| UniProt | AmoebaDB | NCBI | PPIase Name | Localization 3 | References |
|---|---|---|---|---|---|
| C4LYX1 | EHI_117870 | XP_656069.1 | EhCyP10 | ||
| O15729 | EHI_125840 | XP_656494.1 | EhCyP18 (EhCyP) | Cytoplasm | [29,45] |
| C4M7U6 | EHI_020340 | XP_654585.1 | EhCyP20 | Cytoplasm | [29] |
| C4M525 | EHI_128100 | XP_648283.1 | EhCyP21 | Cytoplasm | [29] |
| C4M942 | EHI_083580 | XP_654418.1 | EhCyP22 | Cytoplasm | [29] |
| C4M2J5 | EHI_054760 | XP_654797.2 | EhCyP40 | Nucleus | [29] |
| C4LTN0 | EHI_012390 | XP_655852.2 | EhFKBP18 | ER | [29] |
| C4M276 | EHI_180160 | XP_653822.1 | EhFKBP29 | ||
| C4LTA4 | EHI_044850 | XP_657211.1 | EhFKBP35 | ||
| B1N302 | EHI_051870 | XP_001913568.1 | EhFKBP43 | ||
| C4LUU9 | EHI_178850 | XP_656239.1 | EhFKBP46 | ||
| C4M181 | EHI_188070 | XP_653673.2 | EhPar13 | Cytoplasm and nucleus | [29] |
| C4LT92 | EHI_044730 | XP_657226.1 | EhPar13.25 | Nucleus | [29] |
| UniProt | TriTrypDB | NCBI | PPIase Name 2 | Localization 3 | Function 3 | References |
|---|---|---|---|---|---|---|
| D0A5M6 | Tbg972.11.920(CYPA) | XP_011779241.1 | TbgCyP19 (TbgCyPA) | Cytoplasm, flagellum, and extracellular space | [85,86] | |
| C9ZYX4 | Tbg972.9.6990 | XP_011776889.1 | TbgCyP20.3 | |||
| D0A8E1 | Tbg972.11.10610 | XP_011780206.1 | TbgCyP20.5 | |||
| C9ZIV0 | Tbg.972.2.170 | XP_011771617.1 | TbgCyP21.1 | |||
| C9ZT99 | Tbg972.7.5450 | XP_011774914.1 | TbgCyP21.2 | Extracellular space | [86] | |
| C9ZWH7 | Tbg972.8.7100 | XP_011776042.1 | TbgCyP21.4 | |||
| C9ZRQ0 | Tbg972.7.160 | XP_011774319.1 | TbgCyP24 | |||
| C9ZSQ5 | Tbg972.7.3760 | XP_011774720.1 | TbgCyP25.55 | |||
| C9ZNS2 | Tbg972.5.1880 | XP_011773337.1 | TbgCyP25.56 | |||
| C9ZWA7 | Tbg972.8.6340 | XP_011775972.1 | TbgCyP27.1 | |||
| C9ZXF5 | Tbg972.9.1740 | XP_011776370.1 | TbgCyP27.4 | |||
| C9ZQE6 | Tbg972.6.1040 | XP_011773911.1 | TbgCyP29 | |||
| C9ZVY5 | Tbg972.8.5140 | XP_011775850.1 | TbgCyP30 | |||
| C9ZUX8 | Tbg972.8.1650 | XP_011775493.1 | TbgCyP33 | |||
| C9ZYI8 | Tbg972.9.5630 | XP_011776753.1 | TbgCyP38 | Extracellular space | [86] | |
| C9ZZI1 | Tbg972.9.9060 | XP_011777096.1 | TbgCyP43 | Membrane | [29] | |
| C9ZIB2 | Tbg972.1.930 | XP_011771345.1 | TbgCyP46 | |||
| C9ZPQ4 | Tbg972.5.5220 | XP_011773669.1 | TbgCyP58 | Nucleus | [29] | |
| C9ZZU0 | Tbg972.10.15980 | XP_011778762.1 | TbgCyP100 | |||
| D0A2I5 | Tbg972.10.5640 | XP_011777743.1 | TbgFKBP12 | |||
| C9ZSQ4 | Tbg972.7.3750 | XP_011774719.1 | TbgFKBP12.3 (TbgFKBP12) | Flagellar pocket | Motility and cytokinesis | [87] |
| D0A0P0 | Tbg972.10.19020(MIP) | XP_011779062.1 | TbgFKBP21 | |||
| D0A0P1 | Tbg972.10.19030 | XP_011779063.1 | TbgFKBP36 | |||
| D0A0V5 | Tbg972.10.19710 | XP_011779127.1 | TbgFKBP48 | Extracellular space | [86] | |
| D0A6H9 | Tbg972.11.3980 | XP_011779544.1 | TbgFKBP92 | |||
| C9ZUI9 | Tbg972.8.300(Pin1) | XP_011775354.1 | TbgPar12 (TbgPin1) | Cytoplasm | [88] | |
| C9ZKX9 | Tbg972.3.3260 | XP_011772278.1 | TbgPar13 (TbgPar14) | [88] | ||
| C9ZRL7 | Tbg972.7.2770(Par45) | XP_011774600.1 | TbgPar42 | Nucleus | Cell growth | [88] |
| UniProt | TriTrypDB | NCBI | PDB | PPIase Name | Localization 3 | References |
|---|---|---|---|---|---|---|
| O02614 | LmjF.25.0910 (CYPA) | XP_001683845.1 | LmCyP19 (LmaCyP1) | Cilium, Cytoplasm, and Nucleus | [29,94,95] | |
| Q4QJ67 | LmjF.06.0120 (CYP2) | XP_001680781.1 | LmCyP20.3 (LmaCyP2) | Cytoplasm | [29,96] | |
| Q4QBG3 | LmjF.23.0125 (CyP3) | XP_001683335.1 | LmCyP20.4 (LmaCyP3) | Nucleus | [29,96] | |
| Q4Q424 | LmjF.33.1630 (CYP4) | XP_001685924.1 | LmCyP24 (LmaCyP4) | Cytoplasm | [29,96] | |
| Q4Q6Q9 | LmjF.31.0050 (CYP5) | XP_001684989.1 | LmCyP24.6 (LmaCyP5) | Cytoplasm | [29,96] | |
| Q4QBK2 | LmjF.22.1450 (CYP6) | XP_001683296.1 | 7AIH | LmCyP25 (LmaCyP6) | Cilium, cytoplasm, and nucleus | [29,94,96] |
| E9AFI5 | LmjF.35.3610 (CYP7) | XP_003722755.1 | LmCyP26 (LmaCyP7) | Cilium, cytoplasm, and nucleus | [29,96] | |
| Q4QAK0 | LmjF.24.1315 (CYP8) | XP_001683648.1 | LmCyP26.5 (LmaCyP8) | Cytoplasm | [29,96] | |
| Q4Q7V7 | LmjF.30.0020 (CYP9) | XP_001684591.1 | LmCyP27 (LmaCyP9) | Axoneme and cytoplasm | [29,96] | |
| Q4Q1A6 | LmjF.36.3130 (CYP10) | XP_001686892.1 | 7AM2 | LmCyP29 (LmaCyP10) | Cytoplasm | [29,94,96,97] |
| Q4QBH1 | LmjF.23.0050 (CYP11) | XP_001683327.1 | 2HQJ | LmCyP32 (LmaCyP11) | Cytoplasm and nucleolus | [29,61,96] |
| E9AC11 | LmjF.01.0220 (CYP12) | XP_003721542.1 | LmCyP36 (LmaCyP12) | Axoneme and cytoplasm | [29,96] | |
| E9AFV2 | LmjF.35.4770 (CYP40) | XP_003722872.1 | LmCyP38 (LmaCyp40) | Cytoplasm | [29,96] | |
| Q4QEP7 | LmjF.16.1200 (CYP13) | XP_001682201.1 | LmCyP39 (LmaCyP13) | Axoneme and cytoplasm | [29,96] | |
| E9AEZ3 | LmjF.35.1720 (CYP14) | XP_003722563.1 | LmCyP48 (LmaCyP14) | Cytoplasm and membranes | [29,96] | |
| Q4QCV2 | LmjF.20.0940 (CYP15) | XP_001682846.1 | LmCyP49 LmaCyP15 | [96] | ||
| Q4QDV4 | LmjF.18.0880 (CYP16) | XP_001682494.1 | LmCyP108 LmaCyP16 | Nucleoplasm | [29,96] | |
| Q4QBK4 | LmjF.22.1430 | XP_001683294.1 | LmFKBP-11.8 (maFKBPL1) | Axoneme and cytoplasm | [29,96] | |
| Q4Q255 | LmjF.36.0230 | XP_001686593.1 | LmFKBP-11.9 (LmaFKBPL2) | [96] | ||
| Q4QHC5 | LmjF.10.0890 | XP_001681423.1 | LmFKBP-17.3 (LmaFKBPL3) | [96] | ||
| Q4QDB9 | LmjF.19.0970 | XP_001682679.1 | LmFKBP-23 (LmaFKBPL4) | [96] | ||
| Q4QD56 | LmjF.19.1530 | XP_001682742.1 | LmFKBP-48 (LmaFKBPL5) | [96] | ||
| Q4QII4 | LmjF.07.1030 (PIN1) | XP_001681014.1 | LmPar13 (LmaPPICL1) (LmPIN1) | Cytosol and nucleus | [29,96,98] | |
| Q4QBU3 | LmjF.22.0530 (PAR45) | XP_001683205.1 | LmPar47 (LmaPPICL2) | Nucleus | [29,96] |
| UniProt | TriTrypDB | NCBI | PDB | PPIase Name | Localization 3 | Function 3 | References |
|---|---|---|---|---|---|---|---|
| E9BHJ8 | LdBPK_250940.1 (CYPA) | XP_003861424.1 | LdCyP19 | ||||
| A0A3S7WXE3 | LdBPK_230140.1 | XP_003860915.1 | LdCyP20.3 | ||||
| Q9U9R3 | LdBPK_060120.1 | XP_003858320.1 | 2HAQ | LdCyP20.4 (LdCyP) | Cytoplasm and ER | Disaggregation and aggregation prevention | [99,100,101,102,103] |
| A0A3S7X410 | LdBPK_310060.1 | XP_003863096.1 | LdCyP24 | ||||
| A0A3Q8ICB3 | LdBPK_221300.1 | XP_003860876.1 | LdCyP25 | ||||
| E9BSN7 | LdBPK_353660.1 | XP_003864946.1 | LdCyP26 | ||||
| E9BGZ8 | LdBPK_241350.1 | XP_003861226.1 | LdCyP27 | ||||
| A0A3S7X325 | LdBPK_300020.1 | XP_003862718.1 | LdCyP28 | ||||
| E9BQA4 | LdBPK_331730.1 | XP_003863999.1 | LdCyP28.6 | Membrane | [100] | ||
| E9BU37 | LdBPK_363280.1 | XP_003865443.1 | LdCyP29 | ||||
| E9BG26 A0A504XWA0 | LdBPK_230060.1 | XP_003860907.1 | LdCyP32 | ||||
| A0A451EJ79 | LdBPK_010220.1 | XP_003857835.1 | LdCyP36 | ||||
| A0A3Q8IIG9 | LdBPK_354830.1 | XP_003865060.1 | LdCyP38.4 (LdCyP40) | [96] | |||
| A0A504WZ51 | LdBPK_161250.1 | XP_003859812.1 | LdCyP39 | ||||
| E9BS46 | LdBPK_351710.1 (CYP14) | XP_003864755.1 | LdCyP48.5 | Membrane | [29] | ||
| E9BEP2 | LdBPK_200950.1 | XP_003860435.1 | LdCyP49 | ||||
| E9BDR8 | LdBPK_180880.1 | XP_003860101.1 | LdCyP108 | ||||
| E9BFZ3 | LdBPK_221280.1 | XP_003860874.1 | LdFKBP11.8 | ||||
| E9BT84 | LdBPK_360250.1 | XP_003865142.1 | LdFKBP11.9 | ||||
| E9BAD9 | LdBPK_100940.1 | XP_003858930.1 | LdFKBP17 | ||||
| E9BE85 | LdBPK_190920.1 | XP_003860268.1 | LdFKBP22 | ||||
| E9BEE5 | LdBPK_191560.1 | XP_003860328.1 | LdFKBP47 | ||||
| E9B9B2 | LdBPK_071180.1 | XP_003858557.1 | LdPar12 | ||||
| E9BFR0 | LdBPK_220410.1 | XP_003860791.1 | LdPar17 |
| UniProt | CryptoDB | NCBI | PDB | PPIase Name | Localization 2 | Reference |
|---|---|---|---|---|---|---|
| A3FQA7 | cgd2_4120 | XP_001388243.1 | 2PLU 2POY | CpCyP18 | Cytosol and plasma membrane | [29,61] |
| Q9Y0F5 | cgd2_1660 | XP_001388204.1 | 2POE 2QER | CpCyP19 | Spliceosome | [29,61] |
| F0X4J7 | cgd5_3350 | XP_001388285.1 | CpCyP21.1 | Cytosol and plasma membrane | [29] | |
| Q5CW56 | cgd8_1560 | XP_627063.1 | CpCyP21.2 | Cytosol and plasma membrane | [29] | |
| Q5CSY2 | cgd1_870 | XP_627935.1 | CpCyP23 | Cytosol and plasma membrane | [29] | |
| K9ME21 | cgd8_2350 | XP_001388428.1 | CpCyP34 | Nucleus | [29] | |
| Q5CYY7 | cgd7_520 | XP_628243.1 | CpCyP89 | Spliceosome | [29] | |
| Q5CZ15 | cgd7_210 | XP_628215.1 | CpFKBP-34 | Nucleolus | [29] | |
| Q5CX33 | cgd6_2690 | XP_627621.1 | CpFKBP-37 | Nucleolus | [29] |
| UniProt | CryptoDB | NCBI | PPIase Name | Localization 2 | Reference |
|---|---|---|---|---|---|
| A0A0S4TC12 | Chro.20441 | XP_667665.1 | ChCyp18.4 ChCyP18 | Cytosol and plasma membrane | [29,138] |
| A0A0S4TBF4 | Chro.20180 | XP_667461.1 | ChCyp18.9 ChCyP19 | Spliceosome | [29,138] |
| A0A0S4TFY9 | Chro.50038 | XP_665956.1 | ChCyp17.9 ChCyP21 | Cytosol and plasma membrane | [29,138] |
| A0A0S4TJX7 | Chro.80184 | XP_665525.1 | ChCyP21.2 | Cytosol and plasma membrane | [29,138] |
| A0A0S4TA19 | Chro.10107 | XP_667336.1 | ChCyp22.9 ChCyP23 | Cytosol and plasma membrane | [29,138] |
| A0A0S4TLJ2 | Chro.80276 | XP_666493.1 | ChCyp34.5 ChCyP40 | Nucleus | [29,138] |
| A0A0S4THZ0 | Chro.70067 | XP_666650.1 | ChCyp88.9 ChCyP89 | Spliceosome | [29,138] |
| A0A0S4TJS1 | Chro.70034 | XP_668351.1 | ChFKBP-34 | Nucleolus | [29] |
| A0A0S4TGW0 | Chro.60310 | XP_667889.1 | ChFKBP-37 | Nucleolus | [29] |
| Parasite | PPIase | UniProt | kDa | pI | Expression System | Purification | Catalytic Efficiency 3 | Inhibition | References | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain | Vector | kcat/Km | Inhibitor | IC50 nM 4 | |||||||
| T. cruzi | TcCyP19 | Q9U664 | 18.9 | 8.4 | M15, XL1Blue | pQE30 | IMAC | CsA H-7-94 F-7-62 MeVal-4 | 14.4-18.4 12.5 13.3 15.3 | [75,78,81] | |
| TcCyP21 | Q4DPB9 | 21.1 | 9.1 | BL21 pLysS | pET14 | IMAC | CsA H-7-94 F-7-62 MeVal-4 | 28.7 23.6 25.2 30.0 | [73,78] | ||
| TcCyP25 | Q9NAT5 | 25.6 | 8.5 | Origami | pRSETA | IMAC | CsA H-7-94 F-7-62 MeVal-4 | 31.7 17.2 17.8 30.0 | [73,78] | ||
| TcCyP28 | O76990 | 28.4 | 9.7 | BL21 RIL | pET41b | IMAC | CsA H-7-94 F-7-62 MeVal-4 | 13.1 9.2 10.1 13.5 | [73,78] | ||
| TcCyP34 | K2NAL4 | 33.4 | 9.0 | BL21(DE3) | pRSETA | IMAC | CsA H-7-94 F-7-62 MeVal-4 | >200 5 | [73,78] | ||
| TcCyP38 (TcCyP40) | Q6V7K6 | 38.4 | 5.7 | M15 | pQE30 | IMAC | CsA H-7-94 F-7-62 MeVal-4 | >200 5 | [73] | ||
| T. brucei | TbgCyP19 (TbCypA) | D0A5M6 | 18.7 | 8.3 | M15 | pQE30 | IMAC | [86] | |||
| T. vaginalis | TvCyP19 (TvCyP1) | A2DT06 | 19 | 7.7 | BL21 | pET32a | IMAC, IEX, AC | 7.1 μM−1s−1 4.0 μM−1s−1 | CsA | 7.5 | [27,32] |
| TvCyP19.9 (TvCyP2) | A2DLL4 | 20.0 | 9.1 | BL21 | pET, pGEX2t pET29b | IMAC, IEX, AC | 4.5 μM−1s−1 | [30,31] | |||
| L. major | LmaCyP19 | O02614 | 19.0 | 7.7 | M15 | pQE30, pREP4 pET14b, pTYB1 pGEX4T-3 | IMAC, HIC, AC | 1.5 × 106 M−1s−1 2.6 × 106 M−1s−1 | CsA | Ki = 0.5 3 | [95,98] |
| LmaCyP38 (LmaCyp40) | E9AFV2 | 38.4 | 5.6 | BL21 | pGEX-5X-Strep | AC | [100] | ||||
| L. donovani | LdCyP20.4 (LdCyP) | Q9U9R3 | 17.7 | 6.9 | BL21 pLysS | pET3a, pQE32 | IMAC | [96,97,102,103] | |||
| T. gondii | TgCyP18 | A0A125YZ79 | 18.3 | 6.9 | BL21 | pET28a | IMAC, AC, SEC, RPC | 1.0 × 104 M−1s−1 | [132] | ||
| TgCyP20 | S8F7V1 | 19.6 | 6.0 | AC | [131,134] | ||||||
| TgCyP23 | A0A125YL73 | 22.9 | 7.0 | BL21 | pET28a | IMAC, SEC | 3.8 × 106 M−1s−1 | [132] | |||
| P. falciparum | PfCyP19 (PfCyP19A) | Q76NN7 | 19.0 | 8.2 | BL21 | pET-3a, pET22b+ | IMAC | 6.3 × 106 M−1s−1 1.2 × 107 M−1s−1 | CsA CsC CsD Rapamycin FK506 | 10 581 238 >5000 >10,000 | [108,110,113,114,119] |
| PfCyP22 (PfCyP19B) | Q8IIK8 | 22.0 | 7.1 | BL21 | pET22b+ | IMAC | 2.3 × 106 M−1s−1 5.7 × 106 M−1s−1 | CsA | 10 | [108,113,114,115,117] | |
| PfCyP18.6 (PfCyP19C) | Q8IIK3 | 18.6 | 5.9 | BL21 | pET22b+ | IMAC | [108,119] | ||||
| PfCyP23 | Q8I3I0 | 23.2 | 5.3 | BL21 | pET22b+ | IMAC | [108,119] | ||||
| PfCyP25 (PfCyP24) | Q8I6S4 | 24.9 | 6.7 | BL21 | pET22b+ | IMAC | [108,119,120,122] | ||||
| PfCyP26 | Q8I621 | 26.4 | 8.5 | BL21 | pET22b+ | IMAC | [108,119] | ||||
| PfCyP32 | Q8I5Q4 | 32.3 | 9.8 | Rosetta | pET22b+ | IMAC | [108,119] | ||||
| PfCyP53 (PfCyP52) | Q8ILM0 | 52.7 | 7.0 | BL21 | pET22b+ | IMAC | [108,119] | ||||
| E. histolytica | EhCyP18 (EhCyP) | O15729 | 18.1 | XL1Blue | pTrcHis A | IMAC | CsA | 10 | [45] | ||
| G. intestinalis | GiCyP19 (GiCyP1) | 19.0 | BL21 | pGEX 4T-1 | AC | CsA | 500 | [65] | |||
| GiCyP18 | A8BC67 | 18.0 | 8.4 | BL21 | pColdI | IMAC | [60] | ||||
| T. cruzi | TcFKBP22 (TcMIP) | Q09734 | 22.1 | 6.8 | XL1 Blue | pGEX-2T | AC | 0.745 M−1s−1 | FK506 | 410 | [79,158] |
| T. gondii | TgFCBP57 | Q4VKI5 | 57.2 | 5.5 | BL21(DE3) | pET15b | IMAC | FK506 CsA | 70 750 | [130] | |
| P. falciparum | PfFKBP35 | Q8I4V8 | 34.8 | 5.4 | BL21, TB1 | pMALc2X, pSUMO | IMAC, SEC, AC | 1.7 × 104 M−1s−1 1.0 × 105 M−1s−1 | FK506 Rapamycin D44 | 320, 260 480 125 | [111,112,116,159] |
| P. vivax | PvFKBP25 | A0A1G4H4D0 | 25.2 | 9.5 | BL21(DE3) | pNIC28-Bsa4 | IMAC, SEC | [122] | |||
| PvFKBP34 (PvFKBP35) | A0A565A3M9 | 34.0 | 6.1 | BL21(DE3) | pSUMO | IMAC, SEC | 1.0 × 105 M−1s−1 | FK506 D44 | 160 125 | [122,159] | |
| G. intestinalis | GiFKBP12 | Q8I6M8 | 12.0 | 9.2 | BL21(DE3)-R3-RARE | AVA0421 | IMAC, SEC | [67] | |||
| T. cruzi | TcPar12.6 (TcPin1) | Q4D8F7/ Q4DKA4 | 12.6 | 7.7 | JM109 | pQE30 | IMAC | 3.97 × 105 M−1s−1 1.54 × 104 M−1s−1 | [82,85] | ||
| TcPar13 (TcPar14) | Q4D394/ Q4E641 | 13.3 | 9.4 | BL21(DE3)-CodonPlus RIL | pET-22b+ | IMAC, SEC | 0.194 M−1s−1 | [83] | |||
| TcPar45 | Q4D9J4/ Q4DH56 | 45.5 | 8.7 | BL21(DE3)-CodonPlus RIL | PET28a | IMAC, SEC | 7.1 × 103 M−1s−1 | [83] | |||
| T. brucei | TbgPar12 (TbPin1) | C9ZUI9 | 12.5 | 6 | pET28b | SEC | [147] | ||||
| TbPar42 | C9ZRL7 | 41.7 | 7.1 | pET28b | SEC | [147] | |||||
| L. major | LmaPar13 (LmPIN1) | Q4QII4 | 12.6 | 7.2 | BL21 | IMAC, SEC | [100,101] | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aranda-Chan, V.; Cárdenas-Guerra, R.E.; Otero-Pedraza, A.; Pacindo-Cabrales, E.E.; Flores-Pucheta, C.I.; Montes-Flores, O.; Arroyo, R.; Ortega-López, J. Insights into Peptidyl-Prolyl cis-trans Isomerases from Clinically Important Protozoans: From Structure to Potential Biotechnological Applications. Pathogens 2024, 13, 644. https://doi.org/10.3390/pathogens13080644
Aranda-Chan V, Cárdenas-Guerra RE, Otero-Pedraza A, Pacindo-Cabrales EE, Flores-Pucheta CI, Montes-Flores O, Arroyo R, Ortega-López J. Insights into Peptidyl-Prolyl cis-trans Isomerases from Clinically Important Protozoans: From Structure to Potential Biotechnological Applications. Pathogens. 2024; 13(8):644. https://doi.org/10.3390/pathogens13080644
Chicago/Turabian StyleAranda-Chan, Verónica, Rosa Elena Cárdenas-Guerra, Alejandro Otero-Pedraza, Esdras Enoc Pacindo-Cabrales, Claudia Ivonne Flores-Pucheta, Octavio Montes-Flores, Rossana Arroyo, and Jaime Ortega-López. 2024. "Insights into Peptidyl-Prolyl cis-trans Isomerases from Clinically Important Protozoans: From Structure to Potential Biotechnological Applications" Pathogens 13, no. 8: 644. https://doi.org/10.3390/pathogens13080644