
Spatial Clustering of Rabies by Animal Species in New Jersey, United States, from 1989 to 2023


Abstract
:1. Introduction
2. Materials and Methods
3. Results
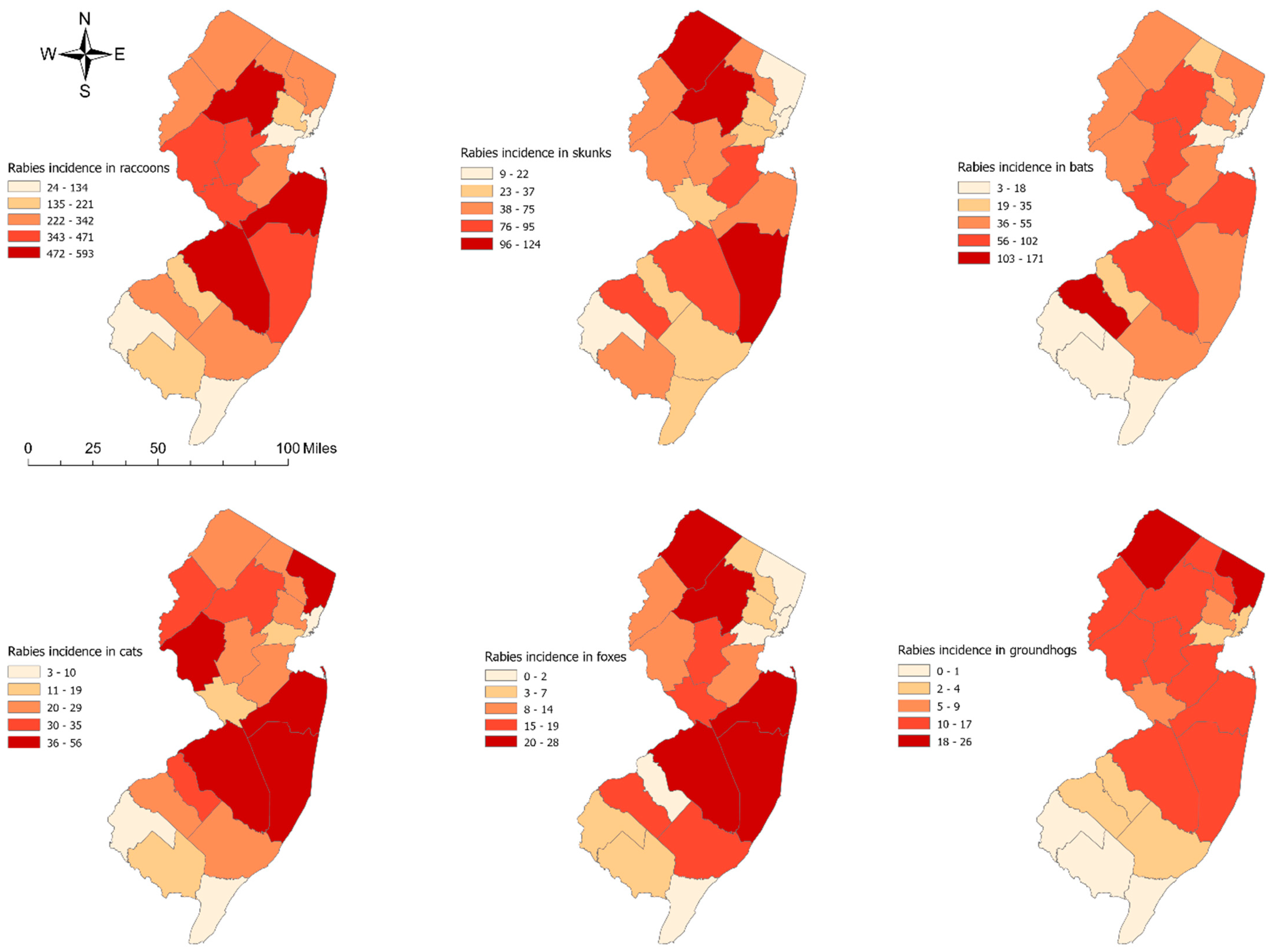
3.1. Spatial Patterns of Rabies Incidence by Animal Species
3.2. Geographic Clusters of Rabies by Animal Species
3.2.1. Global Evidence of Clustering
3.2.2. Local Clusters
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CDC. Rabies in the United States: Protecting Public Health. 2024. Available online: https://www.cdc.gov/rabies/php/protecting-public-health/index.html#:~:text=Testing%3A%20More%20than%20130%20labs%20test%20nearly%20100%2C000,year%2C%20over%204%2C000%20animals%20test%20positive%20for%20rabies (accessed on 10 May 2024).
- CDC. About Rabies. 2024. Available online: https://www.cdc.gov/rabies/about/index.html (accessed on 20 April 2024).
- CDC. Clinical Overview of Rabies. 2024. Available online: https://www.cdc.gov/rabies/hcp/clinical-overview/index.html (accessed on 10 May 2024).
- NJDOH. Rabies Background and Technical Information. 2024. Available online: https://www.nj.gov/health/cd/documents/topics/rabies/rabies_background.pdf (accessed on 2 April 2024).
- Ma, X.; Boutelle, C.; Bonaparte, S.; Orciari, L.A.; Condori, R.E.; Kirby, J.D.; Chipman, R.B.; Fehlner-Gardiner, C.; Thang, C.; Cedillo, V.G.; et al. Rabies surveillance in the United States during 2022. J. Am. Veter. Med. Assoc. 2024, 1, 1–8. [Google Scholar] [CrossRef]
- Uhaa, I.J.; Dato, V.M.; Sorhage, F.E.; Beckley, J.W.; Roscoe, D.E.; Gorsky, R.D.; Fishbein, D.B. Benefits and costs of using an orally absorbed vaccine to control rabies in raccoons. J. Am. Veter. Med. Assoc. 1992, 201, 1873–1882. [Google Scholar] [CrossRef]
- Reilly, S.J. Distribution of Wildlife Rabies in Central Appalachia and Analysis of Factors Influencing Human Exposure. Master’s Thesis, University of Kentucky College of Public Health, Lexington, KY, USA, 2014. [Google Scholar]
- Reilly, S.; Sanderson, W.T.; Christian, W.J.; Browning, S.R. Geographical Clusters and Predictors of Rabies in Three Southeastern States. Vector-Borne Zoonotic Dis. 2017, 17, 432–438. [Google Scholar] [CrossRef]
- Jones, M.E.; Curns, A.T.; Krebs, J.W.; Childs, J.E. Environmental and Human Demographic Features Associated with Epizootic Raccoon Rabies in Maryland, Pennsylvania, and Virginia. J. Wildl. Dis. 2003, 39, 869–874. [Google Scholar] [CrossRef]
- Riley, S.P.; Hadidian, J.; Manski, D.A. Population density, survival, and rabies in raccoons in an urban national park. Can. J. Zool. 1998, 76, 1153–1164. [Google Scholar] [CrossRef]
- Recuenco, S.; Eidson, M.; Kulldorff, M.; Johnson, G.; Cherry, B. Spatial and temporal patterns of enzootic raccoon rabies adjusted for multiple covariates. Int. J. Health Geogr. 2007, 6, 14. [Google Scholar] [CrossRef] [PubMed]
- Anthony, J.A.; Childs, J.E.; Glass, G.E.; Korch, G.W.; Ross, L.; Grigor, J.K. Land use associations and changes in population indices of urban raccoons during a rabies epizootic. J. Wildl. Dis. 1990, 26, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Lucey, B.; Russell, C.; Smith, D.; Wilson, M.; Long, A.; Waller, L.; Childs, J.; Real, L. Spatiotemporal Analysis of Epizootic Raccoon Rabies Propagation in Connecticut, 1991–1995. Vector-Borne Zoonotic Dis. 2002, 2, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Hunt, N.; Carroll, A.; Wilson, T.P. Spatiotemporal Analysis and Predictive Modeling of Rabies in Tennessee. J. Geogr. Inf. Syst. 2018, 10, 89–110. [Google Scholar] [CrossRef]
- Wheeler, D.C.; Waller, L.A. Mountains, valleys, and rivers: The transmission of raccoon rabies over a heterogeneous landscape. J. Agric. Biol. Environ. Stat. 2008, 13, 388–406. [Google Scholar] [CrossRef]
- Guerra, M.A.; Curns, A.T.; Rupprecht, C.E.; Hanlon, C.A.; Krebs, J.W.; Childs, J.E. Skunk and Raccoon Rabies in the Eastern United States: Temporal and Spatial Analysis. Emerg. Infect. Dis. 2003, 9, 1143–1150. [Google Scholar] [CrossRef] [PubMed]
- Lawson, A.B. Statistical Methods in Spatial Epidemiology; John Wiley & Sons: Hoboken, NJ, USA, 2013; ISBN 9780470009673. [Google Scholar]
- Rupprecht, C.E.; Buchanan, T.; Cliquet, F.; King, R.; Müller, T.; Yakobson, B.; Yang, D.-K. A global perspective on oral vaccination of wildlife against rabies. J. Wildl. Dis. 2024, 60, 241–284. [Google Scholar] [CrossRef] [PubMed]
- Willsey, A.; Laniewicz, B.R.; Eidson, M.; Trimarchi, C.V.; Rudd, R.; Safford, M. Oral rabies vaccination of wildlife in the eastern Adirondack region of New York State. In Proceedings of the Conference Program and Abstracts of the 12th Annual Rabies in the Americas, Peterborough, ON, Canada, 12–16 November 2001. [Google Scholar]
- Sidwa, T.J.; Wilson, P.J.; Moore, G.M.; Oertli, E.H.; Hicks, B.N.; Rohde, R.E.; Johnston, D.H. Evaluation of oral rabies vaccination programs for control of rabies epizootics in coyotes and gray foxes: 1995–2003. J. Am. Veter. Med. Assoc. 2005, 227, 785–792. [Google Scholar] [CrossRef] [PubMed]
- USDA. Oral Rabies Vaccination. 2024. Available online: https://www.aphis.usda.gov/national-wildlife-programs/rabies/vaccine (accessed on 25 August 2024).
- National Association of State Public Health Veterinarians; Compendium of Animal Rabies Prevention and Control Committee; Brown, C.M.; Slavinski, S.; Ettestad, P.; Sidwa, T.J.; Sorhage, F.E.; Prevention, C.O.A.R. Compendium of Animal Rabies Prevention and Control, 2016. J. Am. Veter. Med. Assoc. 2016, 248, 505–517. [Google Scholar] [CrossRef] [PubMed]
- USDA. U.S. National Plan for Wildlife Rabies Management 2023–2027. 2023. Available online: https://www.aphis.usda.gov/sites/default/files/national-plan-wildlife-rabies-management-23-27.pdf (accessed on 25 August 2024).
- USDA. ORV Information by State. 2024. Available online: https://www.aphis.usda.gov/national-wildlife-programs/rabies/orv-state (accessed on 26 August 2024).
- MapsOfWorld. New Jersey Counties. 2024. Available online: https://www.mapsofworld.com/usa/states/new-jersey/counties/#:~:text=The%20US%20state%20of%20New%20Jersey%20is%20divided,cities%2C%20244%20townships%2C%2015%20towns%2C%20and%20four%20villages (accessed on 14 March 2024).
- NJGOV. State of New Jersey Profile. 2024. Available online: https://www.nj.gov/njoem/mitigation/pdf/2019/mit2019_section4_State_Profile.pdf#:~:text=New%20Jersey%20is%20located%20in%20the%20Mid-Atlantic%20region,a%20population%20of%208%2C904%2C413%20%28ACS%205yr%20Estimates%2C%202015%29 (accessed on 15 March 2024).
- University, R. Population Density and New Jersey’s Two Largest Racial/Ethnic Minority Groups in 2020. 2020. Available online: https://policylab.rutgers.edu/population-density-and-new-jerseys-two-largest-racial-ethnic-minority-groups-in-2020/ (accessed on 21 March 2024).
- Worldpopulationreview. New Jersey Population 2024. 2024. Available online: https://worldpopulationreview.com/states/new-jersey-population (accessed on 15 March 2024).
- NJDOH. Rabies Testing. 2024. Available online: https://www.nj.gov/health/phel/public-health-lab-testing/rabies/ (accessed on 20 May 2024).
- NJDOH. Animal Rabies Statistics 2024. Available online: https://www.nj.gov/health/cd/statistics/rabies-stats/ (accessed on 13 March 2024).
- NJOGIS. County Boundaries of New Jersey. 2024. Available online: https://njogis-newjersey.opendata.arcgis.com/datasets/newjersey::county-boundaries-of-nj/explore/ (accessed on 15 March 2024).
- Jenks, G.F. The data model concept in statistical mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Auchincloss, A.H.; Gebreab, S.Y.; Mair, C.; Roux, A.V.D. A Review of Spatial Methods in Epidemiology, 2000–2010. Annu. Rev. Public Health 2012, 33, 107–122. [Google Scholar] [CrossRef]
- Mitchell, A. The ESRI Guide to GIS Analysis: Geographic Patterns & Relationships; ESRI, Inc.: Redlands, CA, USA, 1999; Volume 1. [Google Scholar]
- Anselin, L.; Syabri, I.; Kho, Y. GeoDa: An introduction to spatial data analysis. In Handbook of Applied Spatial Analysis: Software tools, Methods and Applications; Springer: Berlin/Heidelberg, Germany, 2009; pp. 73–89. [Google Scholar]
- Kulldorff, M.; Nagarwalla, N. Spatial disease clusters: Detection and inference. Stat. Med. 1995, 14, 799–810. [Google Scholar] [CrossRef]
- Anselin, L. Local indicators of spatial association—LISA. Geogr. Anal. 1995, 27, 93–115. [Google Scholar] [CrossRef]
- Ord, J.K.; Getis, A. Testing for Local Spatial Autocorrelation in the Presence of Global Autocorrelation. J. Reg. Sci. 2001, 41, 411–432. [Google Scholar] [CrossRef]
- Anselin, L. Exploring spatial data with GeoDaTM: A workbook. Cent. Spatially Integr. Soc. Sci. 2005, 1963, 157. [Google Scholar]
- Desktop, E.A. ArcGIS Pro; Environmental Systems Research Institute: Redlands, CA, USA, 2019. [Google Scholar]
- Recuenco, S.; Eidson, M.; Cherry, B.; Kulldorff, M.; Johnson, G. Factors associated with endemic raccoon (Procyon lotor) rabies in terrestrial mammals in New York State, USA. Prev. Veter. Med. 2008, 86, 30–42. [Google Scholar] [CrossRef]
- Recuenco, S.E. Use of Raccoon Rabies Spatial Patterns to Optimize Efficiency of Oral Rabies Vaccine Interventions. Ph.D. Thesis, State University of New York at Albany, New York, NY, USA, 2006. [Google Scholar]
- Smith, D.; Waller, L.; Russell, C.; Childs, J.; Real, L. Assessing the role of long-distance translocation and spatial heterogeneity in the raccoon rabies epidemic in Connecticut. Prev. Veter. Med. 2005, 71, 225–240. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Dyer, J.L.; Blanton, J.D.; Kuzmin, I.V.; Rupprecht, C.E. Rabies in rodents and lagomorphs in the United States, 1995–2010. J. Am. Veter. Med. Assoc. 2014, 245, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.R.; Winkler, W.G. Descriptive epidemiology from an epizootic of raccoon rabies in the middle atlantic states, 1982–1983. Am. J. Epidemiol. 1987, 126, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.W.; Strine, T.W.; Smith, J.S.; Rupprecht, C.E.; Childs, J.E. Rabies surveillance in the United States during 1993. J. Am. Veter. Med. Assoc. 1994, 205, 1695–1709. [Google Scholar] [CrossRef]
- Smith, D.L.; Lucey, B.; Waller, L.A.; Childs, J.E.; Real, L.A. Predicting the spatial dynamics of rabies epidemics on heterogeneous landscapes. Proc. Natl. Acad. Sci. USA 2002, 99, 3668–3672. [Google Scholar] [CrossRef]
- Roscoe, D.E.; Holste, W.C.; Sorhage, F.E.; Campbell, C.; Niezgoda, M.; Buchannan, R.; Diehl, D.; Niu, H.S.; Rupprecht, C.E.; Hong Douglas, E.; et al. Efficacy of an oral vaccinia-rabies glycoprotein recombinant vaccine in controlling epidemic raccoon rabies in New Jersey. J. Wildl. Dis. 1998, 34, 752–763. [Google Scholar] [CrossRef]
- Ma, X.; Blanton, J.D.; Rathbun, S.L.; Recuenco, S.; Rupprecht, C.E. Time Series Analysis of the Impact of Oral Vaccination on Raccoon Rabies in West Virginia, 1990–2007. Vector-Borne Zoonotic Dis. 2010, 10, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Sterner, R.T.; Meltzer, M.I.; Shwiff, S.A.; Slate, D. Tactics and Economics of Wildlife Oral Rabies Vaccination, Canada and the United States. Emerg. Infect. Dis. 2009, 15, 1176–1184. [Google Scholar] [CrossRef]
- Buzzelli, M. Modifiable areal unit problem. Int. Encycl. Hum. Geogr. 2020, 169–173. [Google Scholar] [CrossRef]
- Elmore, S.A.; Chipman, R.B.; Slate, D.; Huyvaert, K.P.; VerCauteren, K.C.; Gilbert, A.T. Management and modeling approaches for controlling raccoon rabies: The road to elimination. PLoS Neglected Trop. Dis. 2017, 11, e0005249. [Google Scholar] [CrossRef]
- Brunker, K.; Hampson, K.; Horton, D.L.; Biek, R. Integrating the landscape epidemiology and genetics of RNA viruses: Rabies in domestic dogs as a model. Parasitology 2012, 139, 1899–1913. [Google Scholar] [CrossRef] [PubMed]
- Elser, J.L.; Bigler, L.L.; Anderson, A.M.; Maki, J.L.; Lein, D.H.; Shwiff, S.A. The Economics of a Successful Raccoon Rabies Elimination Program on Long Island, New York. PLoS Neglected Trop. Dis. 2016, 10, e0005062. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Bonaparte, S.; Corbett, P.; Orciari, L.A.; Gigante, C.M.; Kirby, J.D.; Chipman, R.B.; Fehlner-Gardiner, C.; Thang, C.; Cedillo, V.G.; et al. Rabies surveillance in the United States during 2021. J. Am. Veter. Med. Assoc. 2023, 261, 1045–1053. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.H.; Tinline, R.R. Rabies control in wildlife. In Rabies; Elsevier: Amsterdam, The Netherlands, 2003; pp. 445–471. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Animal Species | Global Moran’s I (p-Value) |
|---|---|
| Raccoons | 0.32 (0.012) |
| Foxes | 0.29 (0.011) |
| Groundhogs | 0.37 (0.005) |
| Cats | 0.17 (0.071) |
| Bats | −0.09 (0.375) |
| Skunks | 0.10 (0.132) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarkar, S.; Meliker, J.R. Spatial Clustering of Rabies by Animal Species in New Jersey, United States, from 1989 to 2023. Pathogens 2024, 13, 742. https://doi.org/10.3390/pathogens13090742
Sarkar S, Meliker JR. Spatial Clustering of Rabies by Animal Species in New Jersey, United States, from 1989 to 2023. Pathogens. 2024; 13(9):742. https://doi.org/10.3390/pathogens13090742
Chicago/Turabian StyleSarkar, Shamim, and Jaymie R. Meliker. 2024. "Spatial Clustering of Rabies by Animal Species in New Jersey, United States, from 1989 to 2023" Pathogens 13, no. 9: 742. https://doi.org/10.3390/pathogens13090742