
Kharon Is Crucial for Trypanosoma cruzi Morphology but Does Not Impair In Vitro Infection


 , , , ,
, , , ,  , and
, and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Bioinformatic Analysis
2.2. Parasite Maintenance and Growth Curve
2.3. Molecular Cloning and In Vitro Transcription
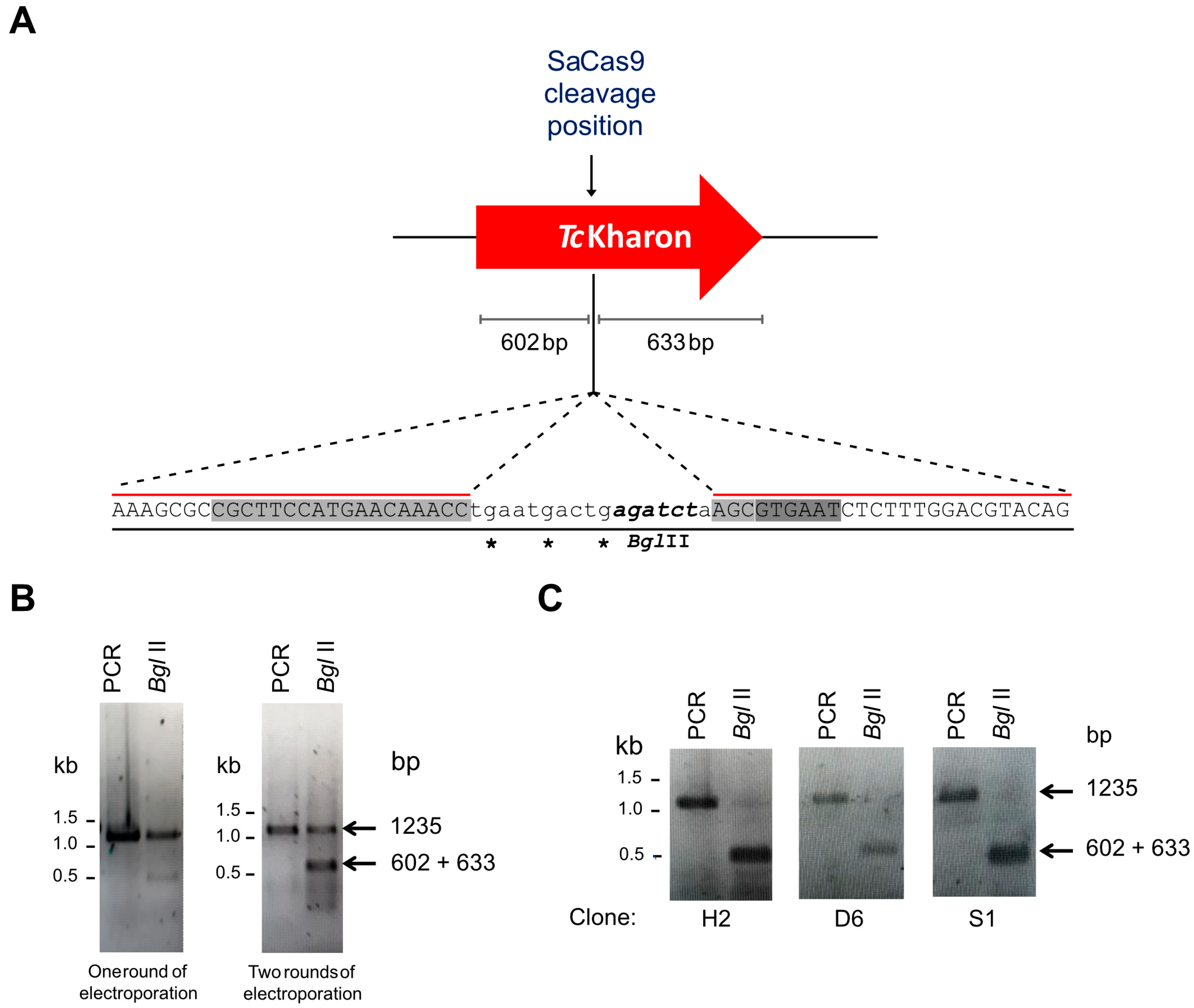
2.4. CRISPR/Cas9 Editing and Genotyping
2.5. Scanning and Transmission Electron Microscopy
2.6. Immunofluorescence Assay and Localization of GFP-Tagged TcKharon
2.7. Western Blot
2.8. Cell Infection Assays and Tissue-Cultured Derived Trypomastigote Counts
2.9. Statistical Analysis
3. Results
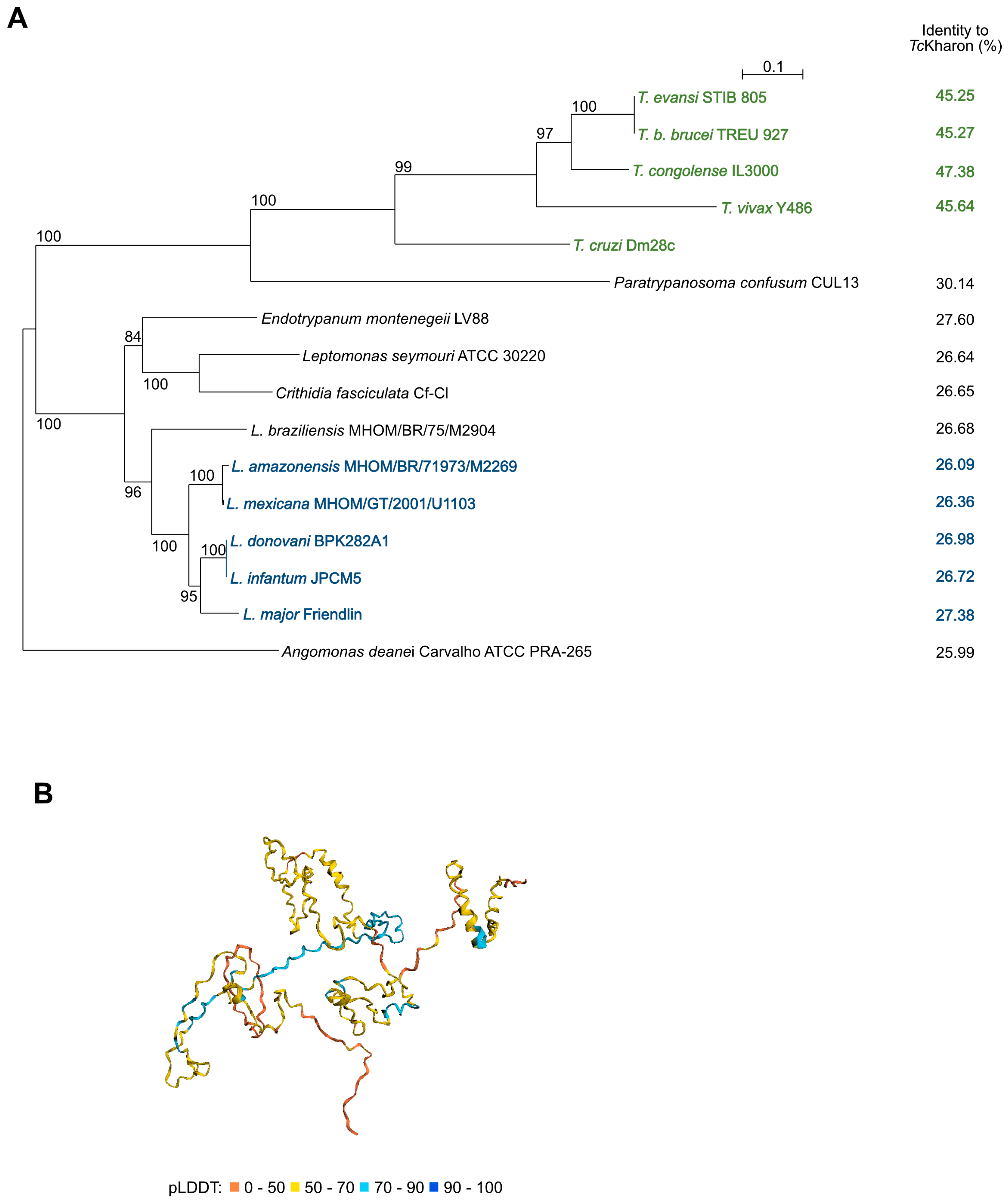
3.1. Kharon Is an Intrinsically Disordered Protein Based on the 3D Structure Prediction
3.2. TcKharon::GFP Is Localized at the Subpellicular Cytoskeleton
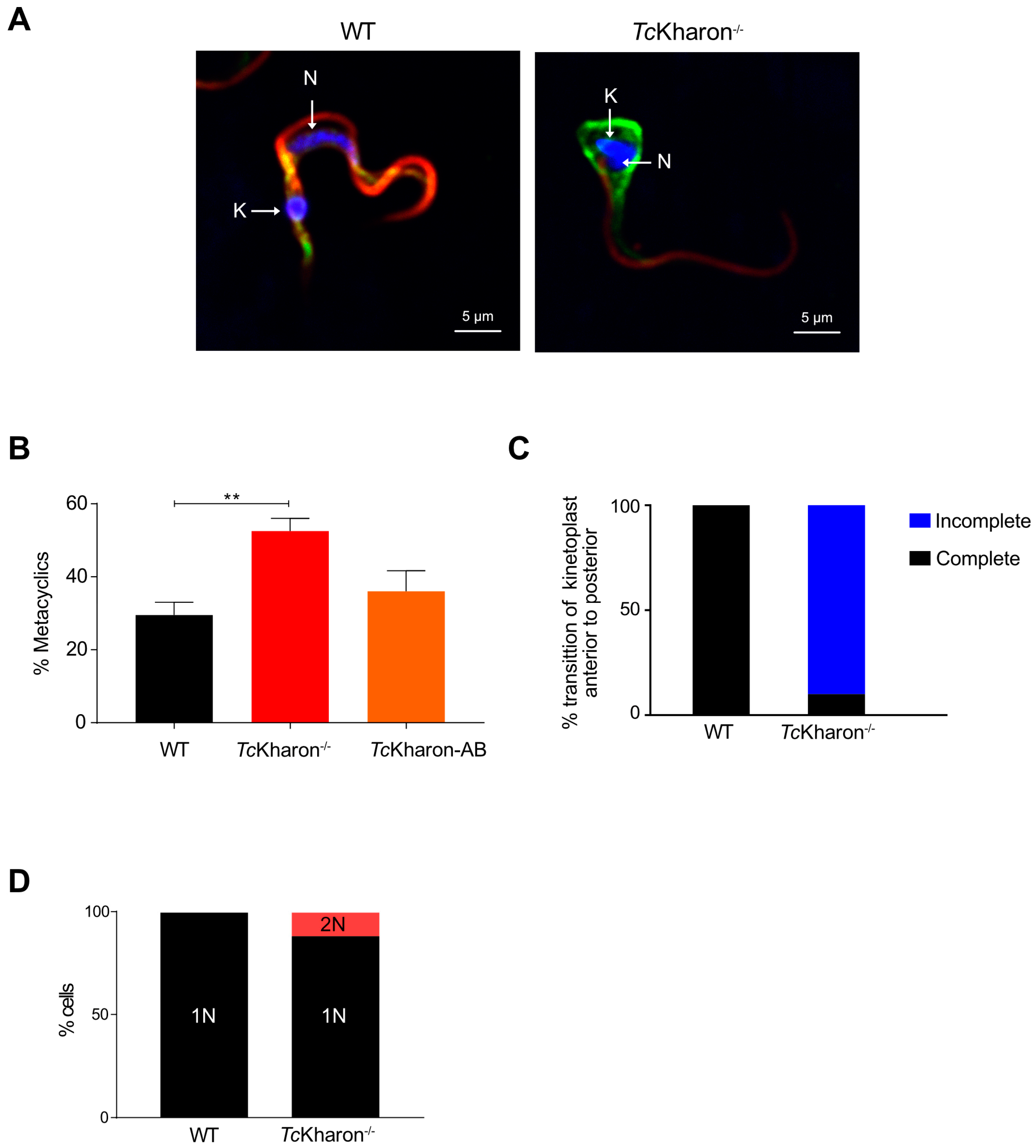
3.3. TcKharon Disruption Affects Cell Growth and Cytokinesis of Epimastigotes
3.4. Morphology of TcKharon-Null Epimastigotes Strongly Affects the Posterior Cell Region
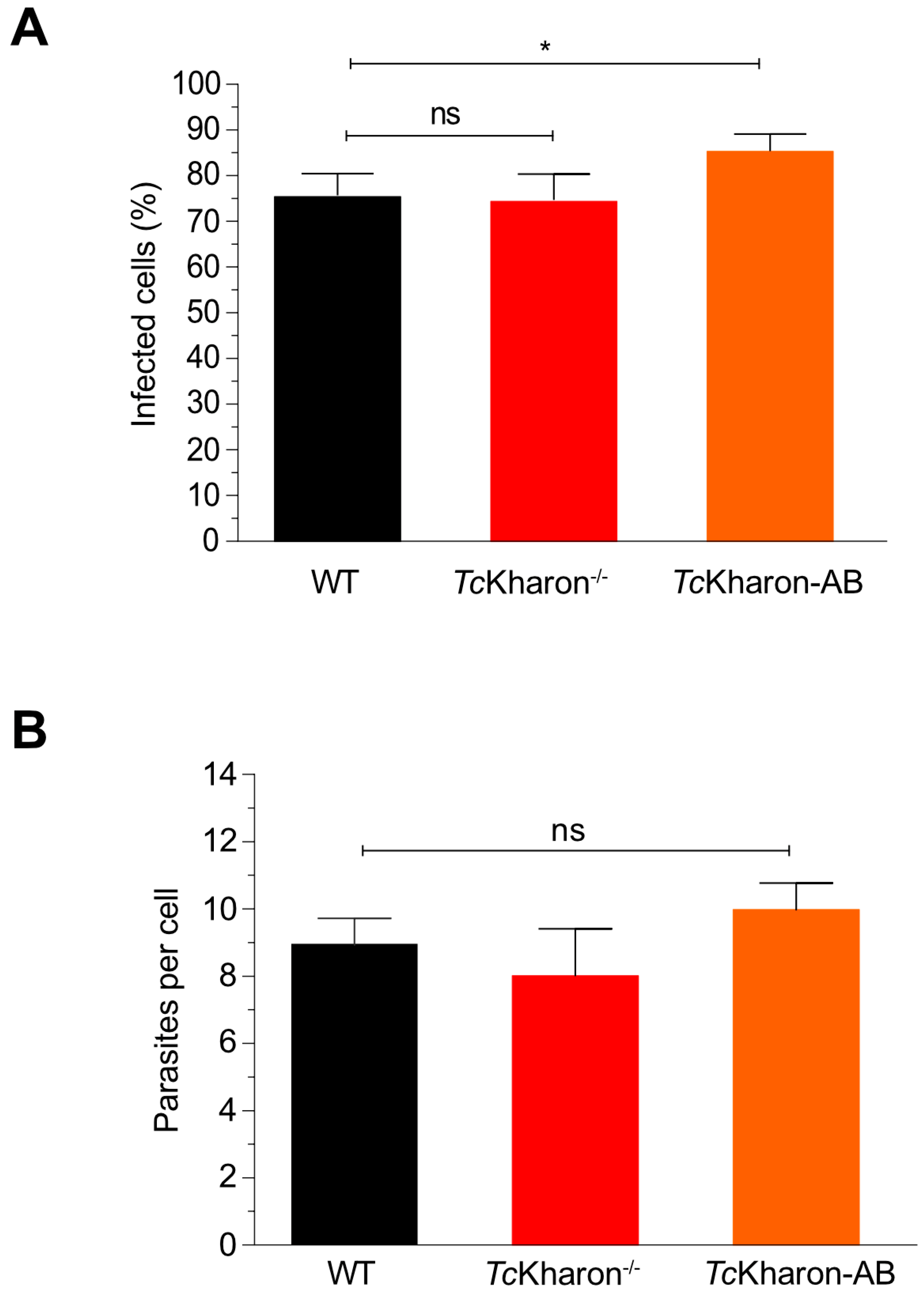
3.5. Metacyclogenesis and Cell Invasion of the TcKharon−/− Mutant Are Not Affected
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gómez-Ochoa, S.A.; Rojas, L.Z.; Echeverría, L.E.; Muka, T.; Franco, O.H. Global, Regional, and National Trends of Chagas Disease from 1990 to 2019: Comprehensive Analysis of the Global Burden of Disease Study. Glob. Heart 2022, 17, 59. [Google Scholar] [CrossRef]
- Kratz, J.M.; Garcia Bournissen, F.; Forsyth, C.J.; Sosa-Estani, S. Clinical and pharmacological profile of benznidazole for treatment of Chagas disease. Expert Rev. Clin. Pharmacol. 2018, 11, 943–957. [Google Scholar] [CrossRef]
- Rios, L.; Campos, E.E.; Menon, R.; Zago, M.P.; Garg, N.J. Epidemiology and pathogenesis of maternal-fetal transmission of Trypanosoma cruzi and a case for vaccine development against congenital Chagas disease. Biochim. Biophys. Acta Mol. Basis. Dis. 2020, 1866, 165591. [Google Scholar] [CrossRef]
- Junqueira, C.; Santos, L.I.; Galvão-Filho, B.; Teixeira, S.M.; Rodrigues, F.G.; DaRocha, W.D.; Chiari, E.; Jungbluth, A.A.; Ritter, G.; Gnjatic, S.; et al. Trypanosoma cruzi as an effective cancer antigen delivery vector. Proc. Natl. Acad. Sci. USA 2011, 108, 19695–19700. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.P.M.M.; Machado, L.F.M.; Doro, D.; Nascimento, F.C.; Damasceno, L.; Gazzinelli, R.T.; Fernandes, A.P.; Junqueira, C. New vaccine formulations containing a modified version of the Amastigote 2 antigen and the non-virulent Trypanosoma cruzi CL-14 strain are highly antigenic and protective against Leishmania infantum challenge. Front. Immunol. 2018, 9, 465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.W.; Karmakar, S.; Gannavaram, S.; Dey, R.; Lypaczewski, P.; Ismail, N.; Siddiqui, A.; Simonyan, V.; Oliveira, F.; Coutinho-Abreu, I.V.; et al. A second generation leishmanization vaccine with a markerless attenuated Leishmania major strain using CRISPR gene editing. Nat. Commun. 2020, 11, 3461. [Google Scholar] [CrossRef]
- Burle-Caldas, G.A.; Dos Santos, N.S.A.; de Castro, J.T.; Mugge, F.L.B.; Grazielle-Silva, V.; Oliveira, A.E.R.; Pereira, M.C.A.; Reis-Cunha, J.L.; dos Santos, A.C.; Gomes, D.A.; et al. Disruption of active Trans-sialidase genes impairs egress from mammalian host cells and generates highly attenuated Trypanosoma cruzi parasites. mBio 2022, 13, e0347821. [Google Scholar] [CrossRef]
- Perez Brandan, C.; Padilla, A.M.; Xu, D.; Tarleton, R.L.; Basombrio, M.A. Knockout of the DHFR-TS gene in Trypanosoma cruzi generates attenuated parasites able to confer protection against a virulent challenge. PLoS Negl. Trop. Dis. 2011, 5, e1418. [Google Scholar] [CrossRef]
- Rooholamin, Z.; Dianat-Moghadam, H.; Esmaeilifallah, M.; Khanahmad, H. From classical approaches to new developments in genetic engineering of live attenuated vaccine against cutaneous leishmaniasis: Potential and immunization. Front. Public Health 2024, 12, 1382996. [Google Scholar] [CrossRef]
- Moreira, P.O.L.; Nogueira, P.M.; Monte-Neto, R.L. Next-generation leishmanization: Revisiting molecular targets for selecting genetically engineered live-attenuated Leishmania. Microorganisms 2023, 11, 1043. [Google Scholar] [CrossRef]
- Souza, W. Structural organization of Trypanosoma cruzi. Mem. Inst. Oswaldo Cruz. 2009, 104 (Suppl. 1), 89–100. [Google Scholar] [CrossRef]
- Billington, K.; Halliday, C.; Madden, R.; Dyer, P.; Barker, A.R.; Moreira-Leite, F.F.; Carrington, M.; Vaughan, S.; Hertz-Fowler, C.; Dean, S.; et al. Genome-wide subcellular protein map for the flagellate parasite Trypanosoma brucei. Nat. Microbiol. 2023, 8, 533–547. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.N.; Huynh, C.T.; Sladewski, T.E.; Zuromski, J.L.; Ruiz, A.E.; de Graffenried, C.L. The Trypanosoma brucei subpellicular microtubule array is organized into functionally discrete subdomains defined by microtubule associated proteins. PLoS Pathog. 2021, 17, e1009588. [Google Scholar] [CrossRef] [PubMed]
- Santi, A.M.M.; Lanza, J.S.; Tunes, L.G.; Fiuza, J.A.; Roy, G.; Orfanó, A.D.S.; De Carvalho, A.T.; Frézard, F.; De Barros, A.L.B.; Murta, S.M.F.; et al. Growth arrested live-attenuated Leishmania infantum KHARON1 null mutants display cytokinesis defect and protective immunity in mice. Sci. Rep. 2018, 8, 11627. [Google Scholar] [CrossRef]
- Kelly, F.D.; Tran, K.D.; Hatfield, J.; Schmidt, K.; Sanchez, M.A.; Landfear, S.M. A cytoskeletal protein complex is essential for division of intracellular amastigotes of Leishmania mexicana. J. Biol. Chem. 2020, 295, 13106–13122. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.D.; Rodriguez-Contreras, D.; Vieira, D.P.; Yates, P.A.; David, L.; Beatty, W.; Elferich, J.; Landfear, S.M. KHARON1 mediates flagellar targeting of a glucose transporter in Leishmania mexicana and is critical for viability of infectious intracellular amastigotes. J. Biol. Chem. 2013, 288, 22721–22733. [Google Scholar] [CrossRef]
- Sanchez, M.A.; Tran, K.D.; Valli, J.; Hobbs, S.; Johnson, E.; Gluenz, E.; Landfear, S.M. KHARON is an essential cytoskeletal protein involved in the trafficking of flagellar membrane proteins and cell division in african trypanosomes. J. Biol. Chem. 2016, 291, 19760–19773. [Google Scholar] [CrossRef]
- Tran, K.D.; Vieira, D.P.; Sanchez, M.A.; Valli, J.; Gluenz, E.; Landfear, S.M. Kharon1 null mutants of Leishmania mexicana are avirulent in mice and exhibit a cytokinesis defect within macrophages. PLoS ONE 2015, 10, e0134432. [Google Scholar] [CrossRef]
- Affolter, M.; Hemphill, A.; Roditi, I.; Müller, N.; Seebeck, T. The repetitive microtubule-associated proteins MARP-1 and MARP-2 of Trypanosoma brucei. J. Struct. Biol. 1994, 112, 241–251. [Google Scholar] [CrossRef]
- Schock, M.; Schmidt, S.; Ersfeld, K. Novel cytoskeleton-associated proteins in Trypanosoma brucei are essential for cell morphogenesis and cytokinesis. Microorganisms 2021, 9, 2234. [Google Scholar] [CrossRef]
- Cooper, R.; de Jesus, A.R.; Cross, G.A. Deletion of an immunodominant Trypanosoma cruzi surface glycoprotein disrupts flagellum-cell adhesion. J. Cell Biol. 1993, 122, 149–156. [Google Scholar] [PubMed]
- LaCount, D.J.; Barrett, B.; Donelson, J.E. Trypanosoma brucei FLA1 is required for flagellum attachment and cytokinesis. J. Biol. Chem. 2002, 277, 17580–17588. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Camargo, E.P. Growth and differentiation in Trypanosoma cruzi. I. Origin of metacyclic trypanosomes in liquid media. Rev. Inst. Med. Trop. Sao Paulo 1964, 6, 93–100. [Google Scholar] [PubMed]
- Contreras, V.T.; Salles, J.M.; Thomas, N.; Morel, C.M.; Goldenberg, S. In vitro differentiation of Trypanosoma cruzi under chemically defined conditions. Mol. Biochem. Parasitol. 1985, 16, 315–327. [Google Scholar] [CrossRef]
- Saenz-Garcia, J.L.; Borges, B.S.; Souza-Melo, N.; Machado, L.V.; Miranda, J.S.; Pacheco-Lugo, L.A.; Moretti, N.S.; Wheleer, R.; Medeiros, L.C.S.; DaRocha, W.D. Trypanin disruption affects the motility and infectivity of the protozoan Trypanosoma cruzi. Front. Cell. Infect. Microbiol. 2022, 11, 807236. [Google Scholar] [CrossRef]
- Peng, D.; Tarleton, R. EuPaGDT: A web tool tailored to design CRISPR guide RNAs for eukaryotic pathogens. Microb Genom. 2015, 1, e000033. [Google Scholar] [CrossRef]
- Kangussu-Marcolino, M.M.; Cunha, A.P.; Avila, A.R.; Herman, J.P.; DaRocha, W.D. Conditional removal of selectable markers in Trypanosoma cruzi using a site-specific recombination tool: Proof of concept. Mol. Biochem. Parasitol. 2014, 198, 71–74. [Google Scholar] [CrossRef]
- Pacheco-Lugo, L.; Díaz-Olmos, Y.; Sáenz-García, J.; Probst, C.M.; DaRocha, W.D. Effective gene delivery to Trypanosoma cruzi epimastigotes through nucleofection. Parasitol. Int. 2017, 66, 236–239. [Google Scholar] [CrossRef]
- Souza-Melo, N.; Alcantara, C.L.; Vidal, J.C.; Rocha, G.M.; de Souza, W. Implications of flagellar attachment zone proteins TcGP72 and TcFLA-1BP in morphology, proliferation, and intracellular dynamics in Trypanosoma cruzi. Pathogens 2023, 12, 1367. [Google Scholar] [CrossRef]
- de Almeida, J.M.; Nunes, F.O.; Ceole, L.F.; Klimeck, T.D.F.; da Cruz, L.A.; Tófoli, D.; Borges, B.S.; Garcez, W.S.; Tozetti, I.A.; Medeiros, L.C.S.; et al. Synergistic effect and ultrastructural changes in Trypanosoma cruzi caused by isoobtusilactone A in short exposure of time. PLoS ONE 2021, 16, e0245882. [Google Scholar] [CrossRef] [PubMed]
- Ramos, T.C.P.; Freymüller-Haapalainen, E.; Schenkman, S. Three-dimensional reconstruction of Trypanosoma cruzi epimastigotes and organelle distribution along the cell division cycle: 3D Electron Microscopy of Trypanosoma cruzi. Cytom. Part A 2011, 79A, 538–544. [Google Scholar] [CrossRef]
- DaRocha, W.D.; Silva, R.A.; Bartholomeu, D.C.; Pires, S.F.; Freitas, J.M.; Macedo, A.M.; Vazquez, M.P.; Levin, M.J.; Teixeira, S.M.R. Expression of exogenous genes in Trypanosoma cruzi: Improving vectors and electroporation protocols. Parasitol. Res. 2004, 92, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.M.; Pappu, R.V. AlphaFold and implications for intrinsically disordered proteins. J. Mol. Biol. 2021, 433, 167208. [Google Scholar] [CrossRef]
- Wheeler, R.J. A resource for improved predictions of Trypanosoma and Leishmania protein three-dimensional structure. PLoS ONE 2021, 16, e0259871. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Gadelha, C.; Wickstead, B.; de Souza, W.; Gull, K.; Cunha-e-Silva, N. Cryptic paraflagellar rod in endosymbiont-containing kinetoplastid protozoa. Eukaryot. Cell. 2005, 4, 516–525. [Google Scholar] [CrossRef]
- Sousa, M.A.D. A simple method to purify biologically and antigenically preserved bloodstream trypomastigotes of Trypanosoma cruzi using Deae-cellulose columns. Mem. Inst. Oswaldo Cruz 1983, 78, 317–333. [Google Scholar] [CrossRef]
- Elias, M.C.; da Cunha, J.P.; de Faria, F.P.; Mortara, R.A.; Freymüller, E.; Schenkman, S.; de Faria, F.P. Morphological events during the Trypanosoma cruzi cell cycle. Protist 2007, 158, 147–157. [Google Scholar] [CrossRef]
- Wheeler, R.J.; Gull, K.; Sunter, J.D. Coordination of the cell cycle in Trypanosomes. Annu. Rev. Microbiol. 2019, 73, 133–154. [Google Scholar] [CrossRef]
- Robinson, D.R.; Sherwin, T.; Ploubidou, A.; Byard, E.H.; Gull, K. Microtubule polarity and dynamics in the control of organelle positioning, segregation, and cytokinesis in the trypanosome cell cycle. J. Cell Biol. 1995, 128, 1163–1172. [Google Scholar] [CrossRef]
- Sinclair, A.N.; de Graffenried, C.L. More than microtubules: The structure and function of the subpellicular array in Trypanosomatids. Trends Parasitol. 2019, 35, 760–777. [Google Scholar] [CrossRef] [PubMed]
- Baines, A.; Gull, K. WCB is a C2 domain protein defining the plasma membrane-sub-pellicular microtubule corset of kinetoplastid parasites. Protist 2008, 159, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Vedrenne, C.; Giroud, C.; Robinson, D.R.; Besteiro, S.; Bosc, C.; Bringaud, F.; Baltz, T. Two related subpellicular cytoskeleton-associated proteins in Trypanosoma brucei stabilize microtubules. Mol. Biol. Cell 2002, 13, 1058–1070. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saenz-Garcia, J.L.; Souza-Melo, N.; Miranda, J.S.; Borges, B.; Pacheco-Lugo, L.A.; Quintero-Solano, J.M.; Moretti, N.; Wheeler, R.; Soares-Medeiros, L.C.; DaRocha, W.D. Kharon Is Crucial for Trypanosoma cruzi Morphology but Does Not Impair In Vitro Infection. Pathogens 2025, 14, 312. https://doi.org/10.3390/pathogens14040312
Saenz-Garcia JL, Souza-Melo N, Miranda JS, Borges B, Pacheco-Lugo LA, Quintero-Solano JM, Moretti N, Wheeler R, Soares-Medeiros LC, DaRocha WD. Kharon Is Crucial for Trypanosoma cruzi Morphology but Does Not Impair In Vitro Infection. Pathogens. 2025; 14(4):312. https://doi.org/10.3390/pathogens14040312
Chicago/Turabian StyleSaenz-Garcia, Jose Luis, Normanda Souza-Melo, Juliana Severo Miranda, Beatriz Borges, Lisandro A. Pacheco-Lugo, Jose M. Quintero-Solano, Nilmar Moretti, Richard Wheeler, Lia C. Soares-Medeiros, and Wanderson D. DaRocha. 2025. "Kharon Is Crucial for Trypanosoma cruzi Morphology but Does Not Impair In Vitro Infection" Pathogens 14, no. 4: 312. https://doi.org/10.3390/pathogens14040312
APA StyleSaenz-Garcia, J. L., Souza-Melo, N., Miranda, J. S., Borges, B., Pacheco-Lugo, L. A., Quintero-Solano, J. M., Moretti, N., Wheeler, R., Soares-Medeiros, L. C., & DaRocha, W. D. (2025). Kharon Is Crucial for Trypanosoma cruzi Morphology but Does Not Impair In Vitro Infection. Pathogens, 14(4), 312. https://doi.org/10.3390/pathogens14040312