Persistence of Babesia microti Infection in Humans

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
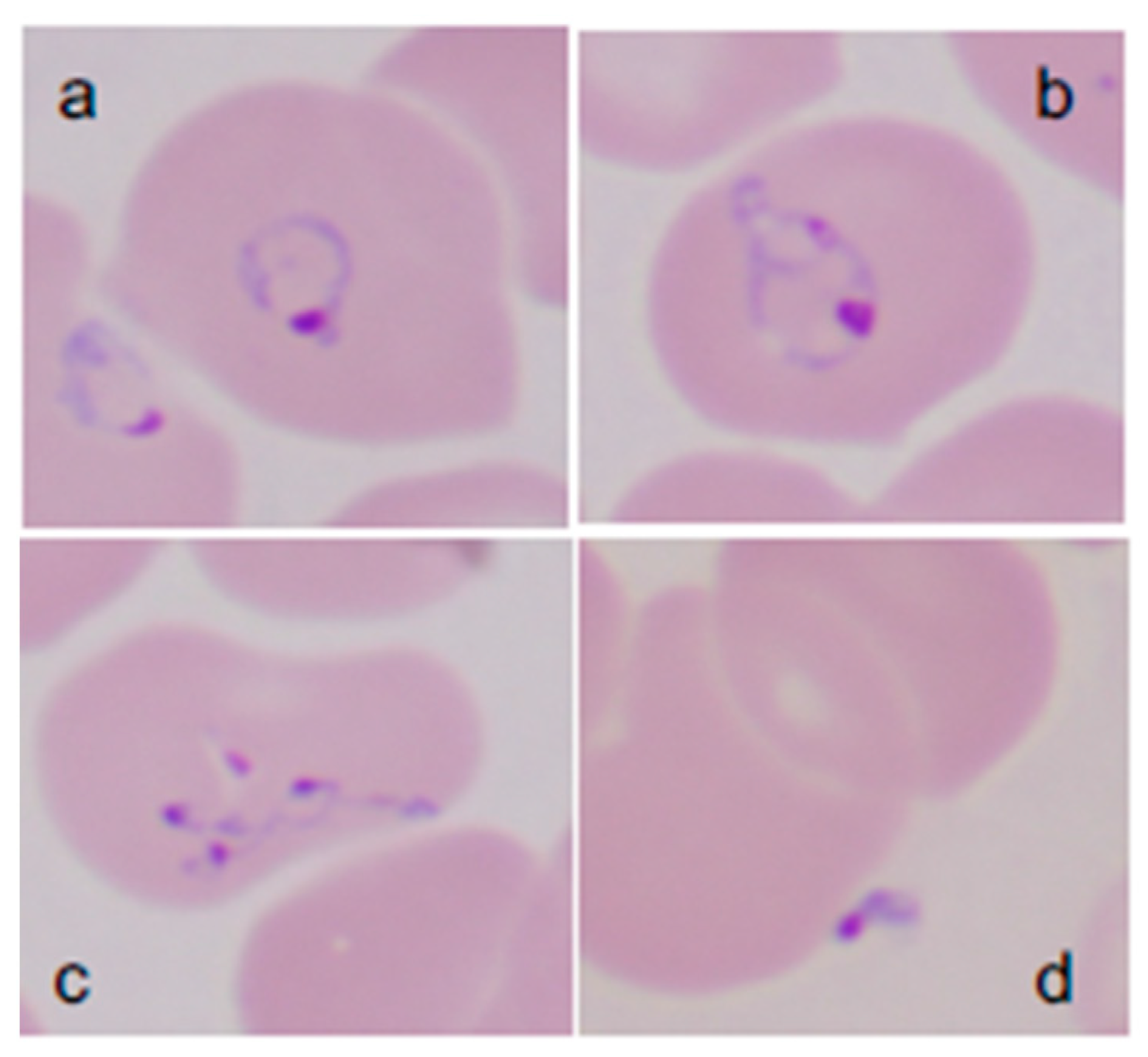
2. Babesia microti Detection Tools
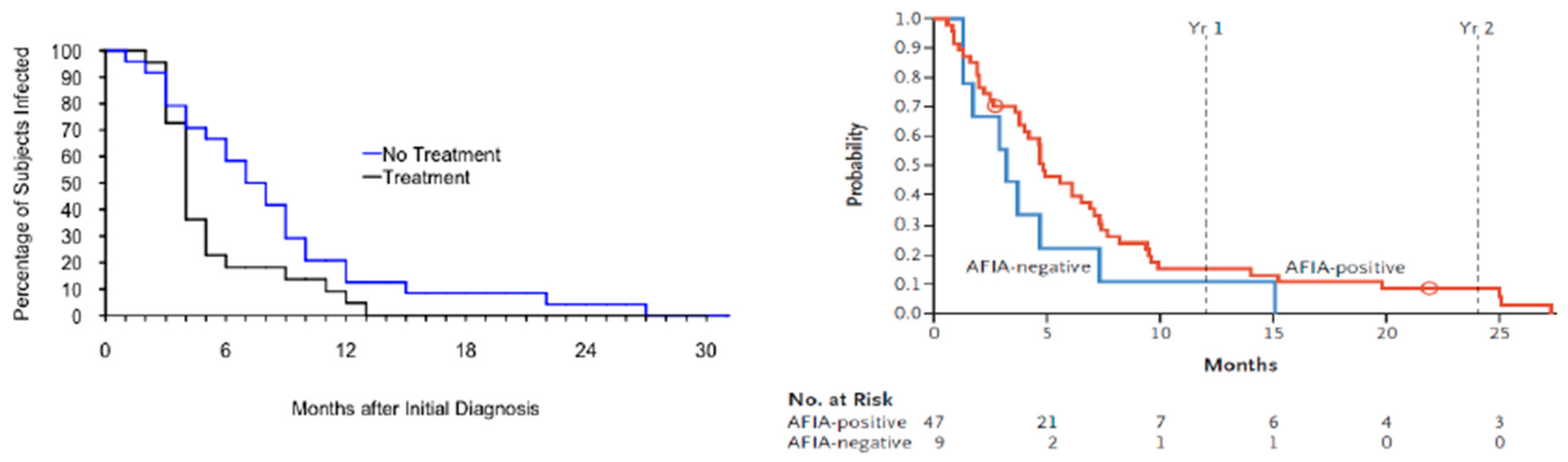
3. Persistent Babesia microti Infection in Immunocompetent and Immunocompromised Hosts
4. Persistent Babesia microti infection in Asymptomatic Immune-Intact Blood Donors
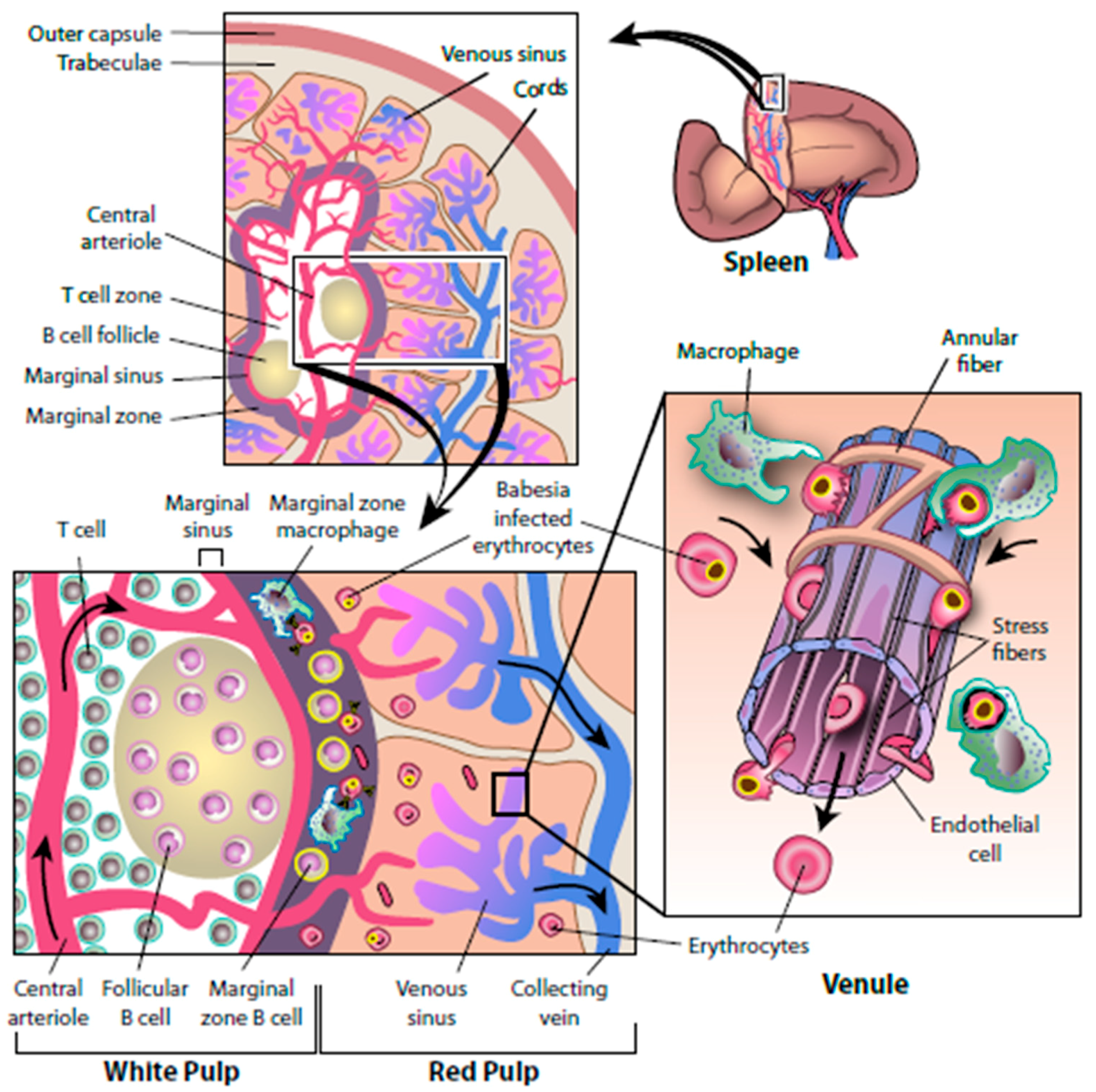
5. Mechanisms of Babesia Persistence
6. Conclusions
- Persistence of Babesia infection in mammalian hosts is an important characteristic of these parasites that maximizes the probability of transmission between vector and host.
- Asymptomatic persistence of Babesia infection in blood donors increases the probability of transmission to blood transfusion recipients.
- Persistent asymptomatic Babesia infection in blood donors is being addressed through implementation of blood donor screening.
- Symptomatic persistence of Babesia infection in immunocompromised hosts increases the probability of disease complications and death.
- The health burden of persistent and recrudescent babesiosis can be minimized by development of novel therapeutic measures, such as new antiparasitic drugs or drug combinations, improved antiparasitic drug duration strategies, or immunoglobulin preparations; and novel preventive approaches, including early detection methods, tick-avoidance, and blood donor screening.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vannier, E.; Krause, P.J. Human Babesiosis. N. Engl. J. Med. 2012, 66, 2397–2407. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; McKay, K.; Gadbaw, J.; Christianson, D.; Closter, L.; Lepore, T.; Telford, S.R., 3rd; Sikand, V.; Ryan, R.; Persing, D.; et al. Increasing health burden of human babesiosis in endemic sites. Am. J. Trop. Med. Hyg. 2003, 68, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Leiby, D.A.; Chung, A.P.; Gill, J.E.; Houghton, R.L.; Persing, D.H.; Badon, S.; Cable, R.G. Demonstrable parasitemia among Connecticut blood donors with antibodies to Babesia microti. Transfusion 2005, 45, 1804–1810. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Linden, J.V.; Bosserman, E.; Young, C.; Olkowska, D.; Wilson, M. Transfusion-associated babesiosis in the United States: A description of cases. Ann Intern Med. 2011, 155, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Daily, J.; Telford, S.R.; Vannier, E.; Lantos, P.; Spielman, A. Shared features in the pathobiology of babesiosis and malaria. Trends Parasitol. 2007, 23, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.R. Babesiosis: Persistence in the face of adversity. Trends Parasitol. 2003, 19, 51–55. [Google Scholar] [CrossRef]
- White, N.J. Determinants of relapse periodicity in Plasmodium vivax malaria. Malar. J. 2011, 10, 297. [Google Scholar] [CrossRef]
- Trager, W.; Most, H. A long-delayed primary attack of Ovale malaria. Am. J. Trop. Med. Hyg. 1963, 12, 837–839. [Google Scholar] [CrossRef]
- Suarez, C.E.; Alzan, H.F.; Silva, M.G.; Rathinasamy, V.; Poole, W.A.; Cooke, B.M. Unravelling the cellular and molecular pathogenesis of bovine babesiosis: Is the sky the limit? Int. J. Parasitol. 2019, 49, 183–197. [Google Scholar] [CrossRef]
- Krause, P.J.; Spielman, A.; Telford, S.; Sikand, V.J.; McKay, K.; Christianson, D.; Pollack, R.; Brassard, P.; Magera, J.; Ryan, R.; et al. Persistent parasitemia following acute babesiosis. N. Engl. J. Med. 1998, 339, 160–165. [Google Scholar] [CrossRef]
- Moritz, E.; Winton, C.S.; Tonnetti, L.; Townsend, R.L.; Berardi, V.P.; Hewins, M.E.; Weeks, K.E.; Dodd, R.Y.; Stramer, S.L. Screening for Babesia microti in the US blood supply. N. Engl. J. Med. 2016, 375, 2236–2245. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Gewurz, B.E.; Hill, D.; Marty, F.M.; Vannier, E.; Foppa, I.M.; Furman, R.R.; Neuhaus, E.; Skowron, G.; Gupta, S.; et al. Persistent and relapsing babesiosis in immunocompromised patients. Clin. Infect. Dis. 2008, 46, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Raffalli, J.; Wormser, G.P. Persistence of babesiosis for >2 years in a patient on Rituximab for rheumatoid arthritis. Diag. Microbiol. Infect. Dis. 2016, 85, 231–232. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention, DPDx- Laboratory Identification of Parasites of Public Health Concern. Available online: https://www.cdc.gov/dpdx/babesiosis/index.html (accessed on 12 July 2019).
- Krause, P.J.; Telford, S.; Spielman, A.; Ryan, R.; Magera, J.; Rajan, T.V.; Christianson, D.; Alberghini, T.V.; Bow, L.; Persing, D. Comparison of PCR with blood smear and small animal inoculation for diagnosis of Babesia microti parasitemia. J. Clin. Microbiol. 1996, 34, 2791–2794. [Google Scholar] [PubMed]
- Wang, G.; Villafuerte, P.; Zhuge, J.; Visintainer, P.; Wormser, G.P. Comparison of a quantitative PCR assay with peripheral blood smear examination for detection and quantitation of Babesia microti infection in humans. Diagn. Microbiol. Infect. Dis. 2015, 82, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.M.; Lee, T.H.; Krause, P.J.; Telford, S.R., 3rd; Montalvo, L.; Chafets, D.; Usmani-Brown, S.; Lepore, T.J.; Busch, M.P. Development of a real-time polymerase chain reaction assay for sensitive detection and quantitation of Babesia microti infection. Transfusion 2013, 53, 2299–2306. [Google Scholar] [PubMed]
- Grabias, B.; Clement, J.; Krause, P.J.; Lepore, T.; Kumar, S. Superior real-time polymerase chain reaction detection of Babesia microti parasites in whole blood utilizing high-copy BMN antigens as amplification targets. Transfusion 2018, 58, 1924–1932. [Google Scholar] [CrossRef]
- Johnson, S.T.; Van Tassell, E.R.; Tonnetti, L.; Cable, R.G.; Berardi, V.P.; Leiby, D.A. Babesia microti real-time polymerase chain reaction testing of Connecticut blood donors: Potential implications for screening algorithms. Transfusion 2013. [Google Scholar] [CrossRef]
- Cornillot, E.; Hadj-Kaddour, K.; Dassouli, A.; Noel, B.; Ranwez, V.; Vacherie, B.; Augagneur, Y.; Brès, V.; Duclos, A.; Randazzo, S.; et al. Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti. Nucleic Acids Res. 2012, 40, 9102–9114. [Google Scholar] [CrossRef]
- Thekkiniath, J.; Mootien, S.; Lawres, L.; Perrin, B.A.; Gewirtz, M.; Krause, P.J.; Williams, S.; Doggett, J.S.; Ledizet, M.; Ben Mamoun, C. BmGPAC, an antigen capture assay for detection of active Babesia microti infection. J Clin Microbiol. 2018, 25, 56. [Google Scholar] [CrossRef]
- Elton, C.M.; Rodriguez, M.; Ben Mamoun, C.; Lobo, C.A.; Wright, G.J. A library of recombinant Babesia microti cell surface and secreted proteins for diagnostics discovery and reverse vaccinology. Int. J. Parasitol. 2019, 49, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Telford, S.; Ryan, R.; Conrad, P.A.; Conrad, M.; Wilson, M.; Thomford, J.W.; Spielman, A. Diagnosis of babesiosis: Evaluation of a serologic test for the detection of Babesia microti antibody. J. Inf. Dis. 1994, 169, 923–926. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.B.; Herwaldt, B.L. Babesiosis Surveillance—United States, 2011–2015. MMWR Surveill Summ 2019, 68, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Priest, J.W.; Moss, D.M.; Won, K.; Todd, C.W.; Henderson, L.; Jones, C.C.; Wilson, M. Multiplex assay detection of immunoglobulin G antibodies that recognize Babesia microti antigens. Clin. Vacc. Immunol. 2012, 19, 1539–1548. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.E.; Williamson, P.C.; Erwin, J.L.; Cyrus, S.; Bloch, E.M.; Shaz, B.H.; Kessler, D.; Telford, S.R., 3rd; Krause, P.J.; Wormser, G.P.; et al. Determination of Babesia microti seroprevalence in blood donor populations using an investigational enzyme immunoassay. Transfusion 2014, 54, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.E.; Williamson, P.C.; Bloch, E.M.; Clifford, J.; Cyrus, S.; Shaz, B.H.; Kessler, D.; Gorlin, J.; Erwin, J.L.; Krueger, N.X.; et al. Serologic screening of United States blood donors for Babesia microti using an investigational enzyme immunoassay. Transfusion 2016, 56, 1866–1874. [Google Scholar] [CrossRef] [PubMed]
- Hockman, D.; Dong, M.; Zheng, H.; Kumar, S.; Huff, M.D.; Grigorenko, E.; Beanan, M.; Duncan, R. Comparison of multiplex PCR hybridization-based and singleplex real-time PCR-based assays for detection of low prevalence pathogens in spiked samples. J. Microbiol. Methods 2017, 132, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Magni, R.; Luchini, A.; Liotta, L.; Molestina, R.E. Analysis of the Babesia microti proteome in infected red blood cells by a combination of nanotechnology and mass spectrometry. Int. J. Parasitol. 2019, 49, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Ruebush, T.K.; Chisholm, E.S.; Sulzer, A.J.; Healy, G.R. Development and persistence of antibody in persons infected with Babesia microti. Am. J. Trop. Med. Hyg. 1981, 30, 291–292. [Google Scholar] [CrossRef]
- Krause, P.J.; Ryan, R.; Telford, S.; Persing, D.; Spielman, A. Efficacy of an IgM serodiagnostic test for the rapid diagnosis of acute babesiosis. J. Clin. Microbiol. 1996, 34, 2014–2016. [Google Scholar]
- Western, K.A.; Benson, G.D.; Gleason, N.N.; Healy, G.R.; Schultz, M.G. Babesiosis in a Massachusetts resident. N. Engl. J. Med. 1970, 283, 854–856. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Klempner, M.S. Babesiosis in patients with AIDS: A chronic infection presenting as fever of unknown origin. Clin. Infect. Dis. 1996, 22, 809–812. [Google Scholar] [CrossRef] [PubMed]
- Haselbarth, K.; Tenter, A.M.; Brade, V.; Krieger, G.; Hunfeld, K.P. First case of human babesiosis in Germany—Clinical presentation and molecular characterization of the pathogen. Int. J. Med. Microbiol. 2007, 297, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Stowell, C.P.; Gelfand, J.A.; Shepard, J.O.; Kratz, A. Case 17–2007—A 25- year-old woman with relapsing fevers and recent onset of dyspnea. N. Engl. J. Med. 2007, 356, 2313–2319. [Google Scholar] [CrossRef] [PubMed]
- Vannier, E.; Borggraefe, I.; Telford, S.R.; Menon, S.; Brauns, T.; Spielman, A.; Gelfand, J.A.; Wortis, H.H. Age-associated decline in resistance to Babesia microti is genetically determined. J. Infect. Dis. 2004, 189, 1721–1728. [Google Scholar] [CrossRef]
- Mareedu, N.; Schotthoefer, A.M.; Tomkins, J.; Hall, M.C.; Fritsche, T.R.; Frost, H.M. Risk factors for severe infection, hospitalization, and prolonged antimicrobial therapy in patients with babesiosis. Am. J. Trop. Med. Hyg. 2017, 97, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Leiby, D.A. Transfusion-transmitted Babesia spp.: bull’s-eye on Babesia microti. Clin. Microbiol. Rev. 2011, 24, 14–28. [Google Scholar] [CrossRef]
- Tonnetti, L.; Townsend, R.L.; Deisting, B.M.; Haynes, J.M.; Dodd, R.Y.; Stramer, S.L. The impact of Babesia microti blood donation screening. Transfusion 2019, 59, 593–600. [Google Scholar] [CrossRef]
- Bloch, E.M.; Herwaldt, B.L.; Leiby, D.A.; Shaieb, A.; Herron, R.M.; Chervenak, M.; Reed, W.; Hunter, R.; Ryals, R.; Hagar, W.; et al. The third described case of transfusion-transmitted Babesia duncani. Transfusion 2012, 52, 1517–1522. [Google Scholar] [CrossRef]
- Burgess, M.J.; Rosenbaum, E.R.; Pritt, B.S.; Haselow, D.T.; Ferren, K.M.; Alzghoul, B.N.; Rico, J.C.; Sloan, L.M.; Ramanan, P.; Purushothaman, R.; et al. Possible transfusion-transmitted Babesia divergens-like/MO-1 infection in an Arkansas patient. Clin. Infect. Dis. 2017, 64, 1622–1625. [Google Scholar] [CrossRef]
- Faddy, H.M.; Rooks, K.M.; Irwin, P.J.; Viennet, E.; Paparini, A.; Seed, C.R.; Stramer, S.L.; Harley, R.J.; Chan, H.T.; Dennington, P.M.; et al. No evidence for widespread Babesia microti transmission in Australia. Transfusion 2019. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.F.; Delage, G.; Scalia, V.; Lindsay, R.; Bernier, F.; Dubuc, S.; Germain, M.; Pilot, G.; Yi, Q.L.; Fearon, M.A. Seroprevalence of Babesia microti infection in Canadian blood donors. Transfusion 2016, 56, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Bloch, E.M.; Yang, Y.; He, M.; Tonnetti, L.; Liu, Y.; Wang, J.; Guo, Y.; Li, H.; Leiby, D.A.; Shan, H.; et al. A pilot serosurvey of Babesia microti in Chinese blood donors. Vox Sang. 2018, 113, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, B.L.; Neitzel, D.F.; Gorlin, J.B.; Jensen, K.A.; Perry, E.H.; Peglow, W.R.; Slemenda, S.B.; Won, K.Y.; Nace, E.K.; Pieniazek, N.J.; et al. Transmission of Babesia microti in Minnesota through four blood donations from the same donor over a 6-month period. Transfusion 2002, 42, 1154–1158. [Google Scholar] [CrossRef] [PubMed]
- Bakkour, S.; Chafets, D.M.; Wen, L.; Muench, M.O.; Telford, S.R., 3rd; Erwin, J.L.; Levin, A.E.; Self, D.; Bres, V.; Linnen, J.M.; et al. Minimal infectious dose and dynamics of Babesia microti parasitemia in a murine model. Transfusion 2018, 58, 2903–2910. [Google Scholar] [CrossRef] [PubMed]
- Meissner, E.G.; McGillicuddy, J.W.; Squires, J.; Skipper, D.; Self, S.; Wray, D.; Moritz, E.D.; Stramer, S.L.; Nadig, S. Across state lines: Fulminant Babesia microti infection in a liver transplant recipient. Transpl. Infect. Dis. 2017, 19, e12741. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.; Civen, R. Babesiosis acquired through blood transfusion, California, USA. Emerg. Infect. Dis. 2009, 15, 785–787. [Google Scholar] [CrossRef] [PubMed]
- Cangelosi, J.J.; Sarvat, B.; Sarria, J.C.; Herwaldt, B.L.; Indrikovs, A.J. Transmission of Babesia microti by blood transfusion in Texas. Vox Sang. 2008, 95, 331–334. [Google Scholar] [CrossRef] [PubMed]
- Leiby, D.A.; Johnson, S.T.; Won, K.Y.; Nace, E.K.; Slemenda, S.B.; Pieniazek, N.J.; Cable, R.G.; Herwaldt, B.L. A longitudinal study of Babesia microti infection in seropositive blood donors. Transfusion 2014, 54, 2217–2225. [Google Scholar] [CrossRef]
- Bloch, E.M.; Levin, A.E.; Williamson, P.C.; Cyrus, S.; Shaz, B.H.; Kessler, D.; Gorlin, J.; Bruhn, R.; Lee, T.H.; Montalvo, L.; et al. A prospective evaluation of chronic Babesia microti infection in seroreactive blood donors. Transfusion 2016, 56, 1875–1882. [Google Scholar] [CrossRef] [PubMed]
- Moritz, E.D.; Winton, C.S.; Johnson, S.T.; Krysztof, D.E.; Townsend, R.L.; Foster, G.A.; Devine, P.; Molloy, P.; Brissette, E.; Berardi, V.P.; et al. Investigational screening for Babesia microti in a large repository of blood donor samples from nonendemic and endemic areas of the United States. Transfusion 2014, 54, 2226–2236. [Google Scholar] [CrossRef]
- FDA. Recommendations for Reducing the Risk of Transfusion-Transmitted Babesiosis; U.S. Department of Health and Human Services: Silver Spring, MD, USA, 2019.
- Busch, M.P.; Bloch, E.M.; Kleinman, S. Prevention of transfusion-transmitted infections. Blood 2019, 133, 1854–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Coller, K.E.; Marohnic, C.C.; Pfeiffer, Z.A.; Fino, J.R.; Elsing, R.R.; Bergsma, J.; Marcinkus, M.A.; Kar, A.K.; Gumbs, O.H.; et al. Performance evaluation of a prototype architect antibody assay for Babesia microti. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef]
- Rosner, F.; Zarrabi, M.H.; Benach, J.L.; Habicht, G.S. Babesiosis in splenectomized adults: Review of 22 reported cases. Am. J. Med. 1984, 76, 696–701. [Google Scholar] [CrossRef]
- Zintl, A.; Mulcahy, G.; Skerrett, H.E.; Taylor, S.M.; Gray, J.S. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin. Microbiol. Rev. 2003, 16, 622–636. [Google Scholar] [CrossRef]
- Desforges, J.F.; Quimby, F. Babesia abroad. N. Engl. J. Med. 1976, 295, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.; Levine, J.F. Pathology of experimental Babesia microti infection in the Syrian hamster. Lab. Anim. Sci. 1987, 37, 640–643. [Google Scholar] [PubMed]
- Ruebush, T.K., 2nd; Collins, W.E.; Warren, M. Experimental Babesia microti infections in Macaca mulatta: Recurrent parasitemia before and after splenectomy. Am. J. Trop. Med. Hyg. 1981, 30, 304–307. [Google Scholar] [CrossRef]
- Westra, J.; van Assen, S.; Wilting, K.R.; Land, J.; Horst, G.; de Haan, A.; Bijl, M. Rituximab impairs immunoglobulin (Ig)M and IgG (subclass) responses after influenza vaccination in rheumatoid arthritis patients. Clin. Exp. Immunol. 2014, 178, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.R.; Al-Khedery, B. Antigenic variation and cytoadhesion in Babesia bovis and Plasmodium falciparum: Different logics achieve the same goal. Mol. Biochem. Parasitol. 2004, 134, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A.; Budd, A.C.; Hsue, G.; Haymore, B.R.; Joyce, A.J.; Thorner, R.; Krause, P.J. Absence of erythrocyte sequestration in a case of babesiosis in a splenectomized human patient. Malar. J. 2006, 5, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.S.; Westblade, L.F.; Dziedziech, A.; Visone, J.E.; Furman, R.R.; Jenkins, S.G.; Schuetz, A.N.; Kirkman, L.A. Clinical and molecular evidence of atovaquone and azithromycin resistance in relapsed Babesia microti infection associated with Rituximab and chronic lymphocytic leukemia. Clin. Infect. Dis. 2017, 65, 1222–1225. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, J.E.; Tran, A.D.; Freimark, L.; Schaffner, S.F.; Goethert, H.; Andersen, K.G.; Bazner, S.; Li, A.; McGrath, G.; Sloan, L.; et al. A global map of genetic diversity in Babesia microti reveals strong population structure and identifies variants associated with clinical relapse. Nat. Microbiol. 2016, 1, 16079. [Google Scholar] [CrossRef] [PubMed]
- Sacca, R.; Cuff, C.A.; Ruddle, N.H. Mediators of inflammation. Curr. Opinion. Immunol. 1997, 9, 851–857. [Google Scholar] [CrossRef]
- Shaio, M.; Lin, P.R. A case study of cytokine profiles in acute human babesiosis. Am. J. Trop. Med. Hyg. 1998, 58, 335–337. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A.; Alleva, L.M.; Mills, A.C.; Cowden, W.B. Pathogenesis of malaria and clinically similar conditions. Clin. Microbiol. Rev. 2004, 17, 509–539. [Google Scholar] [CrossRef]
- Hemmer, R.M.; Ferrick, D.A.; Conrad, P.A. Up-regulation of tumor necrosis factor alpha and interferon-gamma expression in the spleen and lungs of mice infected with the human Babesia isolate WA1. Parasitol. Res. 2000, 86, 121–128. [Google Scholar] [CrossRef]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bloch, E.M.; Kumar, S.; Krause, P.J. Persistence of Babesia microti Infection in Humans. Pathogens 2019, 8, 102. https://doi.org/10.3390/pathogens8030102
Bloch EM, Kumar S, Krause PJ. Persistence of Babesia microti Infection in Humans. Pathogens. 2019; 8(3):102. https://doi.org/10.3390/pathogens8030102
Chicago/Turabian StyleBloch, Evan M., Sanjai Kumar, and Peter J. Krause. 2019. "Persistence of Babesia microti Infection in Humans" Pathogens 8, no. 3: 102. https://doi.org/10.3390/pathogens8030102
APA StyleBloch, E. M., Kumar, S., & Krause, P. J. (2019). Persistence of Babesia microti Infection in Humans. Pathogens, 8(3), 102. https://doi.org/10.3390/pathogens8030102