Development of an In Vitro Membrane Model to Study the Function of EsxAB Heterodimer and Establish the Role of EsxB in Membrane Permeabilizing Activity of Mycobacterium tuberculosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Procedures
2.1. Lipids
2.2. EsxA and EsxAB Heterodimer Expression and Purification
2.3. EsxAB Heterodimer Separation
2.4. Liposome Preparation
2.5. Time Lapse Intensity Measurement of EsxA Membrane Permeabilization by ANTS/DPX Fluorescence Dequenching
2.6. Time-Lapse Intensity Measurement of NBD Fluorescence for EsxA Membrane Insertion
2.7. EsxB Stabilization Effect on EsxA at 37 °C
3. Results
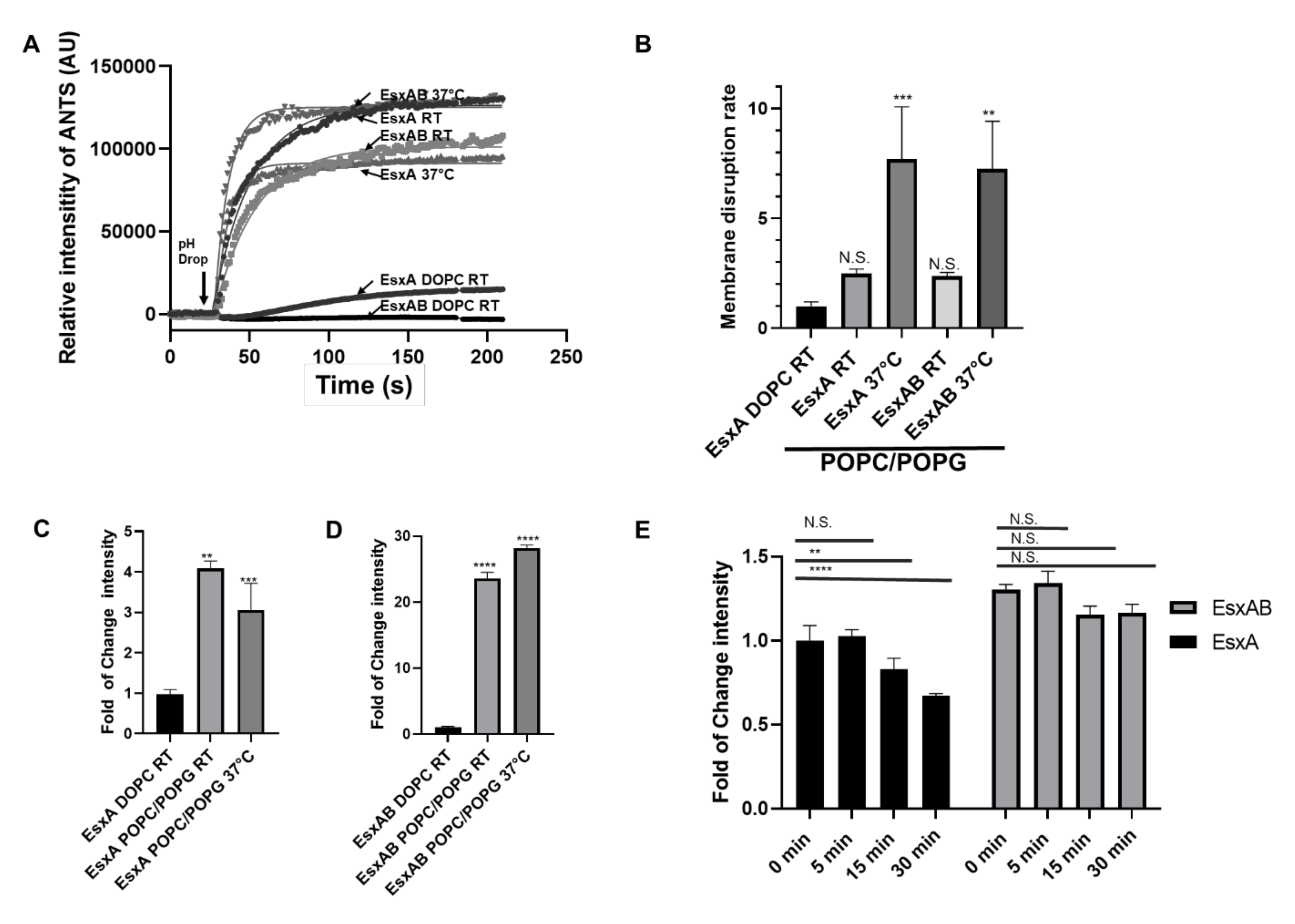
3.1. Incorporation of Negatively Charged Lipids Increased Nα-Acetylated EsxA (N-EsxA) Membrane Permeabilization
3.2. The New Liposome System Was Validated by NBD Fluorescence
3.3. Characterization of the Heterodimer’s MPA in the PC/PG Liposomes
3.4. EsxAB is More Stable at 37 °C Than N-EsxA
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- W.H.O. Global Tuberculosis Report 2020; World Health Organization: Geneva, Switzerland, 2020; Licence: CC BY-NC-SA 3.0 IGO. [Google Scholar]
- Tascon, R.E.; Soares, C.S.; Ragno, S.; Stavropoulos, E.; Hirst, E.M.A.; Colston, M.J. Mycobacterium tuberculosis-activated dendritic cells induce protective immunity in mice. Immunology 2000, 99, 473–480. [Google Scholar] [CrossRef]
- Schlesinger, L.S. Macrophage phagocytosis of virulent but not attenuated strains of Mycobacterium tuberculosis is mediated by mannose receptors in addition to complement receptors. J. Immunol. 1993, 150, 2920–2930. [Google Scholar]
- Wong, D.; Bach, H.; Sun, J.; Hmama, Z.; Av-Gay, Y. Mycobacterium tuberculosis protein tyrosine phosphatase (PtpA) excludes host vacuolar-H +-ATPase to inhibit phagosome acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 19371–19376. [Google Scholar] [CrossRef] [Green Version]
- Brodin, P.; Poquet, Y.; Levillain, F.; Peguillet, I.; Larrouy-Maumus, G.; Gilleron, M.; Ewann, F.; Christophe, T.; Fenistein, D.; Jang, J.; et al. High content phenotypic cell-based visual screen identifies Mycobacterium tuberculosis acyltrehalose-containing glycolipids involved in phagosome remodeling. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pethe, K.; Swenson, D.L.; Alonso, S.; Anderson, J.; Wang, C.; Russell, D.G. Isolation of Mycobacterium tuberculosis mutants defective in the arrest of phagosome maturation. Proc. Natl. Acad. Sci. USA 2004, 101, 13642–13647. [Google Scholar] [CrossRef] [Green Version]
- Philips, J.A.; Ernst, J.D. Tuberculosis Pathogenesis and Immunity. Annu. Rev. Pathol. Mech. Dis. 2012, 7, 353–380. [Google Scholar] [CrossRef]
- Simeone, R.; Bobard, A.; Lippmann, J.; Bitter, W.; Majlessi, L.; Brosch, R.; Enninga, J. Phagosomal rupture by Mycobacterium tuberculosis results in toxicity and host cell death. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef]
- Smith, J.; Manoranjan, J.; Pan, M.; Bohsali, A.; Xu, J.; Liu, J.; McDonald, K.L.; Szyk, A.; LaRonde-LeBlanc, N.; Gao, L.Y. Evidence for pore formation in host cell membranes by ESX-1-secreted ESAT-6 and its role in Mycobacterium marinum escape from the vacuole. Infect. Immun. 2008, 76, 5478–5487. [Google Scholar] [CrossRef] [Green Version]
- Lewis, K.N.; Liao, R.; Guinn, K.M.; Hickey, M.J.; Smith, S.; Behr, M.A.; Sherman, D.R. Deletion of RD1 from Mycobacterium tuberculosis Mimics Bacille Calmette-Guérin Attenuation. J. Infect. Dis. 2003, 187, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Pym, A.S.; Brodin, P.; Majlessi, L.; Brosch, R.; Demangel, C.; Williams, A.; Griffiths, K.E.; Marchal, G.; Leclerc, C.; Cole, S.T. Recombinant BCG exporting ESAT-6 confers enhanced protection against tuberculosis. Nat. Med. 2003, 9, 533–539. [Google Scholar] [CrossRef]
- Guinn, K.M.; Hickey, M.J.; Mathur, S.K.; Zakel, K.L.; Grotzke, J.E.; Lewinsohn, D.M.; Smith, S.; Sherman, D.R. Individual RD1-region genes are required for export of ESAT-6/CFP-10 and for virulence of Mycobacterium tuberculosis. Mol. Microbiol. 2004, 51, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.Y.; Guo, S.; McLaughlin, B.; Morisaki, H.; Engel, J.N.; Brown, E.J. A mycobacterial virulence gene cluster extending RD1 is required for cytolysis, bacterial spreading and ESAT-6 secretion. Mol. Microbiol. 2004, 51, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Berthet, F.X.; Rasmussen, P.B.; Rosenkrands, I.; Andersen, P.; Gicquel, B. A Mycobacterium tuberculosis operon encoding ESAT-6 and a novel low-molecular-mass culture filtrate protein (CFP-10). Microbiology 1998, 51, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Renshaw, P.S.; Panagiotidou, P.; Whelan, A.; Gordon, S.V.; Glyn Hewinson, R.; Williamson, R.A.; Carr, M.D. Conclusive evidence that the major T-cell antigens of the Mycobacterium tuberculosis complex ESAT-6 and CFP-10 form a tight, 1:1 complex and characterization of the structural properties of ESAT-6, CFP-10, and the ESAT-6·CFP-10 complex. Implications for patogenesis and virulence. J. Biol. Chem. 2002, 227, 21598–21603. [Google Scholar] [CrossRef] [Green Version]
- Sherman, D.R.; Guinn, K.M.; Hickey, M.J.; Mathur, S.K.; Zakel, K.L.; Smith, S. Mycobacterium tuberculosis H37Rv:ΔRD1 Is More Virulent than M. bovis Bacille Calmette-Guérin in Long-Term Murine Infection. J. Infect. Dis. 2004, 190, 123–126. [Google Scholar] [CrossRef] [Green Version]
- DiGiuseppe Champion, P.A.; Stanley, S.A.; Champion, M.M.; Brown, E.J.; Cox, J.S. C-terminal signal sequence promotes virulence factor secretion in Mycobacterium tuberculosis. Science 2006, 313, 1632–1636. [Google Scholar] [CrossRef]
- De Jonge, M.I.; Pehau-Arnaudet, G.; Fretz, M.M.; Romain, F.; Bottai, D.; Brodin, P.; Honoré, N.; Marchal, G.; Jiskoot, W.; England, P.; et al. ESAT-6 from Mycobacterium tuberculosis dissociates from its putative chaperone CFP-10 under acidic conditions and exhibits membrane-lysing activity. J. Bacteriol. 2007, 189, 6028–6034. [Google Scholar] [CrossRef] [Green Version]
- De Leon, J.; Jiang, G.; Ma, Y.; Rubin, E.; Fortune, S.; Sun, J. Mycobacterium tuberculosis ESAT-6 Exhibits a Unique Membrane-interacting Activity That Is Not Found in Its Ortholog from Non-pathogenic Mycobacterium smegmatis. J. Biol. Chem. 2012, 287, 44184–44191. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Keil, V.; Sun, J. Characterization of Mycobacterium tuberculosis EsxA membrane insertion: Roles of N- and C-terminal flexible arms and central helix-turn-helix motif. J. Biol. Chem. 2015, 290, 7314–7322. [Google Scholar] [CrossRef] [Green Version]
- Conrad, W.H.; Osman, M.M.; Shanahan, J.K.; Chu, F.; Takaki, K.K.; Cameron, J.; Hopkinson-Woolley, D.; Brosch, R.; Ramakrishnan, L. Mycobacterial ESX-1 secretion system mediates host cell lysis through bacterium contact-dependent gross membrane disruptions. Proc. Natl. Acad. Sci. USA 2017, 114, 1371–1376. [Google Scholar] [CrossRef] [Green Version]
- Meher, A.K.; Bal, N.C.; Chary, K.V.R.; Arora, A. Mycobacterium tuberculosis H37Rv ESAT-6-CFP-10 complex formation confers thermodynamic and biochemical stability. FEBS J. 2006, 273, 1445–1462. [Google Scholar] [CrossRef] [PubMed]
- Lightbody, K.L.; Ilghari, D.; Waters, L.C.; Carey, G.; Bailey, M.A.; Williamson, R.A.; Renshaw, P.S.; Carr, M.D. Molecular features governing the stability and specificity of functional complex formation by Mycobacterium tuberculosis CFP-10/ESAT-6 family proteins. J. Biol. Chem. 2008, 283, 17681–17690. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, J.; Karki, C.B.; Li, L.; Reyes, S.V.; Estevao, I.; Grajeda, B.I.; Zhang, Q.; Arico, C.D.; Ouellet, H.; Sun, J. Nα-Acetylation of the virulence factor EsxA is required for mycobacterial cytosolic translocation and virulence. J. Biol. Chem. 2020, 293, 5785–5794. [Google Scholar] [CrossRef] [Green Version]
- Ray, S.; Vazquez Reyes, S.; Xiao, C.; Sun, J. Effects of membrane lipid composition on Mycobacterium tuberculosis EsxA membrane insertion: A dual play of fluidity and charge. Tuberculosis 2019, 118. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, D.; Jiang, G.; Liu, W.; Deng, Q.; Li, X.; Qian, W.; Ouellet, H.; Sun, J. EsxA membrane-permeabilizing activity plays a key role in mycobacterial cytosolic translocation and virulence: Effects of single-residue mutations at glutamine 5. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Ladokhin, A.S.; Wimley, W.C.; White, S.H. Leakage of membrane vesicle contents: determination of mechanism using fluorescence requenching. Biophys. J. 1995, 69, 1964–1971. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Rutz, J.M.; Feix, J.B.; Klebba, P.E. Permeability properties of a large gated channel within the ferric enterobactin receptor, FepA. Proc. Natl. Acad. Sci. USA 1993, 93, 10653–10657. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Lang, A.E.; Aktories, K.; Collier, R.J. Phenylalanine-427 of anthrax protective antigen functions in both pore formation and protein translocation. Proc. Natl. Acad. Sci. USA 2008, 105, 4346–4351. [Google Scholar] [CrossRef] [Green Version]
- Vanni, S.; Hirose, H.; Barelli, H.; Antonny, B.; Gautier, R. A sub-nanometre view of how membrane curvature and composition modulate lipid packing and protein recruitment. Nat. Commun. 2014, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bissig, C.; Gruenberg, J. Lipid sorting and multivesicular endosome biogenesis. Cold Spring Harb. Perspect. Biol. 2013, 5, a016816. [Google Scholar] [CrossRef] [Green Version]
- Bissig, C.; Johnson, S.; Gruenberg, J. Studying Lipids Involved in the Endosomal Pathway. In Methods in Cell Biology; Academic Press: Pittsburgh, PA, USA, 2012; Volume 108, pp. 19–46. [Google Scholar] [CrossRef]
- Nüsse, O. Biochemistry of the phagosome: The challenge to study a transient organelle. Sci. World J. 2011, 11, 2364–2381. [Google Scholar] [CrossRef] [Green Version]
- Chroneos, Z.C.; Midde, K.; Sever-Chroneos, Z.; Jagannath, C. Pulmonary surfactant and tuberculosis. Tuberculosis 2009, 89. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Guha, S.; Rajani, M.; Padh, H. Identification and characterization of lipids from endosomes purified by electromagnetic chromatography. Indian J. Biochem. Biophys. 2007, 44, 443–449. [Google Scholar]
- Van der Goot, F.G.; Gruenberg, J. Intra-endosomal membrane traffic. Trends Cell Biol. 2006, 16, 514–521. [Google Scholar] [CrossRef]
- Gruenberg, J. Lipids in endocytic membrane transport and sorting. Curr. Opin. Cell Biol. 2003, 15, 382–388. [Google Scholar] [CrossRef]
- Kobayashi, T.; Beuchat, M.H.; Chevallier, J.; Makino, A.; Mayran, N.; Escola, J.M.; Lebrand, C.; Cosson, P.; Kobayashi, T.; Gruenberg, J. Separation and characterization of late endosomal membrane domains. J. Biol. Chem. 2002, 277, 32157–32164. [Google Scholar] [CrossRef] [Green Version]
- Sprong, H.; Van Der Sluijs, P.; Van Meer, G. How proteins move lipids and lipids move proteins. Nat. Rev. Mol. Cell Biol. 2001, 2, 504–513. [Google Scholar] [CrossRef]
- Schmitz, G.; Muller, G. Structure and function of lamellar bodies, lipid-protein complexes involved in storage and secretion of cellular lipids. J. Lipid Res. 1991, 10, 1539–1570. [Google Scholar]
- Sun, J.; Vernier, G.; Wigelsworth, D.J.; Collier, R.J. Insertion of anthrax protective antigen into liposomal membranes: Effects of a receptor. J. Biol. Chem. 2007, 282, 1059–1065. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, B.E.; Grinstein, S. Pathogen destruction versus intracellular survival: The role of lipids as phagosomal fate determinants. J. Clin. Investig. 2008, 118, 2002–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vazquez Reyes, S.; Ray, S.; Aguilera, J.; Sun, J. Development of an In Vitro Membrane Model to Study the Function of EsxAB Heterodimer and Establish the Role of EsxB in Membrane Permeabilizing Activity of Mycobacterium tuberculosis. Pathogens 2020, 9, 1015. https://doi.org/10.3390/pathogens9121015
Vazquez Reyes S, Ray S, Aguilera J, Sun J. Development of an In Vitro Membrane Model to Study the Function of EsxAB Heterodimer and Establish the Role of EsxB in Membrane Permeabilizing Activity of Mycobacterium tuberculosis. Pathogens. 2020; 9(12):1015. https://doi.org/10.3390/pathogens9121015
Chicago/Turabian StyleVazquez Reyes, Salvador, Supriyo Ray, Javier Aguilera, and Jianjun Sun. 2020. "Development of an In Vitro Membrane Model to Study the Function of EsxAB Heterodimer and Establish the Role of EsxB in Membrane Permeabilizing Activity of Mycobacterium tuberculosis" Pathogens 9, no. 12: 1015. https://doi.org/10.3390/pathogens9121015