Salmonella Typhimurium Triggered Unilateral Epididymo-Orchitis and Splenomegaly in a Holstein Bull in Assiut, Egypt: A Case Report


Abstract
:1. Introduction
2. Results
2.1. Microbiological and Sequencing Results:
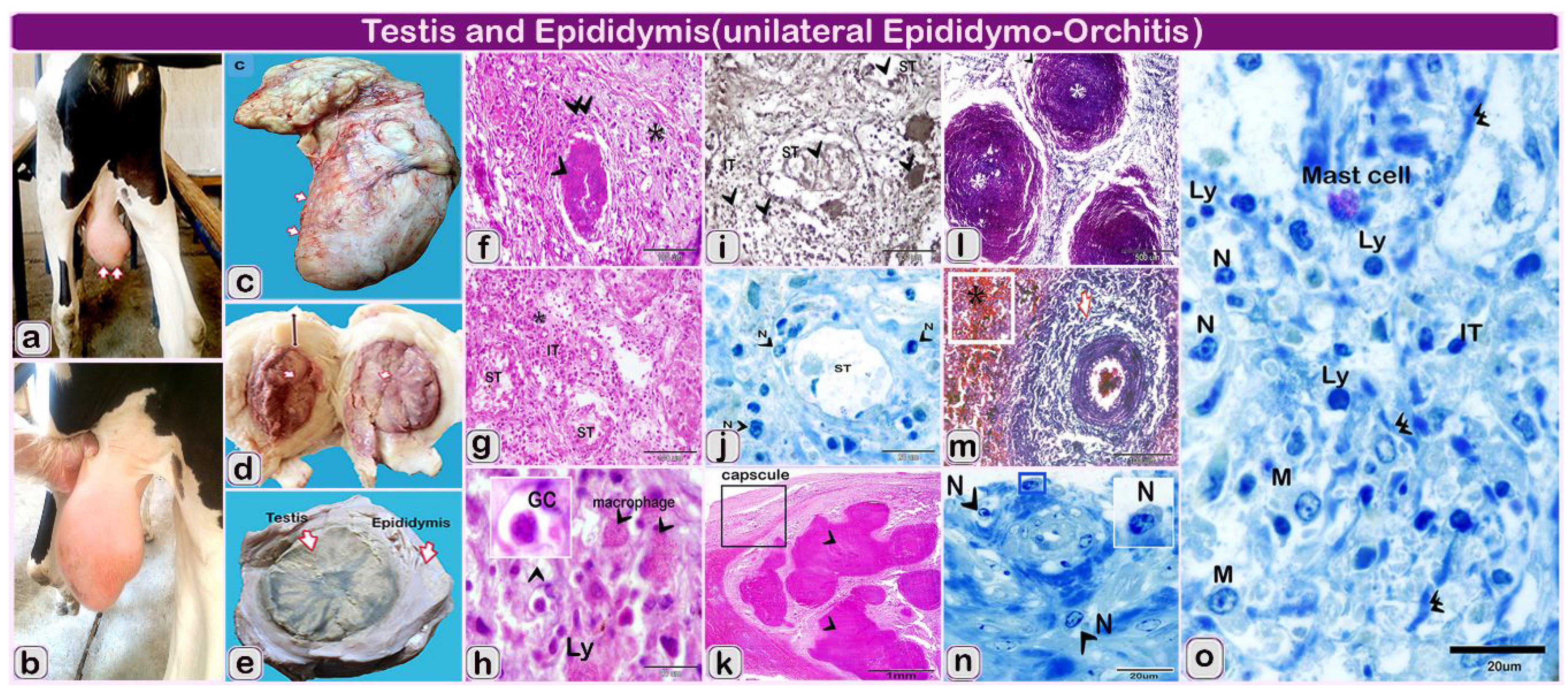
2.2. Histopathological Results
2.2.1. Gross Findings
2.2.2. Microscopically
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Sampling
4.3. Microbiological Examination
4.3.1. VITEK Identification
4.3.2. PCR and Virulence Genes Identification
4.3.3. Gene Sequencing
4.4. Histopathological Examination
4.4.1. Gross Examination
4.4.2. Light Microscopy
Histochemical Investigation
Enzyme Histochemistry
Semi Thin Section Preparations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bansal, N.; Kaistha, N.; Chander, J. Epididymo-orchitis: An unusual manifestation of salmonellosis. J. Microbiol. Immunol. Infect. 2012, 45, 318–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saphra, I.; Winter, J.W. Clinical manifestations of salmonellosis in man: An evaluation of 7779 human infections identified at the New York Salmonella Center. N. Engl. J. Med. 1957, 256, 1128–1134. [Google Scholar] [CrossRef] [PubMed]
- Herenda, D.C.; Chambers, P.; Ettriqui, A. Manual on Meat Inspection for Developing Countries; Food & Agriculture Org.: Rome, Italy, 1994. [Google Scholar]
- Davies, R.; Dalziel, R.; Gibbens, J.; Wilesmith, J.; Ryan, J.; Evans, S.; Byrne, C.; Paiba, G.; Pascoe, S.; Teale, C. National survey for Salmonella in pigs, cattle and sheep at slaughter in Great Britain (1999–2000). J. Appl. Microbiol. 2004, 96, 750–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. The Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Jajere, S.M. A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Vet. World 2019, 12, 504. [Google Scholar] [CrossRef] [Green Version]
- Demirbilek, S.K. Salmonellosis in Animals. In Salmonella-A Re-emerging Pathogen; IntechOpen: Budapest, Hungary, 2017. [Google Scholar]
- Nakano, M.; Yamasaki, E.; Ichinose, A.; Shimohata, T.; Takahashi, A.; Akada, J.K.; Nakamura, K.; Moss, J.; Hirayama, T.; Kurazono, H. Salmonella enterotoxin (Stn) regulates membrane composition and integrity. Dis. Models Mech. 2012, 5, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Bertelloni, F.; Tosi, G.; Massi, P.; Fiorentini, L.; Parigi, M.; Cerri, D.; Ebani, V.V. Some pathogenic characters of paratyphoid Salmonella enterica strains isolated from poultry. Asian Pac. J. Trop. Med. 2017, 10, 1161–1166. [Google Scholar] [CrossRef]
- Zhang, S.; Kingsley, R.A.; Santos, R.L.; Andrews-Polymenis, H.; Raffatellu, M.; Figueiredo, J.; Nunes, J.; Tsolis, R.M.; Adams, L.G.; Bäumler, A.J. Molecular pathogenesis of Salmonella enterica serotype Typhimurium-induced diarrhea. Infect. Immun. 2003, 71, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hapfelmeier, S.; Ehrbar, K.; Stecher, B.; Barthel, M.; Kremer, M.; Hardt, W.-D. Role of the Salmonella pathogenicity island 1 effector proteins SipA, SopB, SopE, and SopE2 in Salmonella enterica subspecies 1 serovar Typhimurium colitis in streptomycin-pretreated mice. Infect. Immun. 2004, 72, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Singh, Y.; Tiwari, A.; Kumar, R.; Saxena, M. Cloning, Sequencing and Phylogenetic Analysis of stn gene of Salmonella Typhimurium. Biosci. Biotechnol. Res. Asia 2017, 14, 1387–1393. [Google Scholar] [CrossRef]
- del Carmen Ferreras, M.; Muñoz, M.; Pérez, V.; Benavides, J.; García-Pariente, C.; Fuertes, M.; Adúriz, G.; García-Marín, J.F. Unilateral orchitis and epididymitis caused by Salmonella enterica subspecies diarizonae infection in a ram. J. Vet. Diagn. Investig. 2007, 19, 194–197. [Google Scholar] [CrossRef] [Green Version]
- Svenungsson, B. Suppurative epididymo-orchitis due to Salmonella Enteritidis. J. Infect. 1984, 8, 64–66. [Google Scholar] [CrossRef]
- Berner, R.; Schumacher, R.; Zimmerhackl, L.; Frankenschmidt, A.; Brandis, M. Salmonella Enteritidis orchitis in a 10–week-old boy. Acta Paediatr. 1994, 83, 992–993. [Google Scholar] [CrossRef]
- Ejlertsen, T.; Jensen, H. Orchitis and testicular abscess formation caused by non-typhoid Salmonellosis: A case report. APMIS 1990, 98, 294–298. [Google Scholar] [CrossRef]
- Bell, G. Diseases of the bovine scrotum. UK Vet. Livest. 2006, 11, 19–23. [Google Scholar] [CrossRef]
- Youngquist, R.S.; Threlfall, W.R. Current Therapy in Large Animal Theriogenology-E-Book; Elsevier Health Sciences: London, UK, 2006. [Google Scholar]
- Jones, T.F.; Ingram, L.A.; Cieslak, P.R.; Vugia, D.J.; Tobin-D’Angelo, M.; Hurd, S.; Medus, C.; Cronquist, A.; Angulo, F.J. Salmonellosis outcomes differ substantially by serotype. J. Infect. Dis. 2008, 198, 109–114. [Google Scholar] [CrossRef]
- Lim, E.; Koh, W.H.; Loh, S.F.; Lam, M.S.; Howe, H.S. Non-thyphoidal salmonellosis in patients with systemic lupus erythematosus. A study of fifty patients and a review of the literature. Lupus 2001, 10, 87–92. [Google Scholar] [CrossRef]
- Boucif, A.; Boulkaboul, A.; Niar, A.; Azzi, N.; Tainturier, D. The testicular pathologies in rams of the Algerian local breed “Rembi” clinical and histopathological classification. Asian J. Anim. Vet. Adv. 2011, 6, 831–837. [Google Scholar] [CrossRef]
- González, F.S.; Encinas, M.G.; Napal, S.L.; Rajab, R. Brucellar orchiepididymitis with abscess. Arch. Esp. Urol. 1997, 50, 289–292. [Google Scholar]
- Alizadeh, M.; Shahnavaz, H.; Hojjati, H.N. A Case of Granulomatous Orchitis Due to Brucellosis with Unusual Clinical and Paraclinical Findings. Arch. Clin. Infect. Dis. 2013, 8, e14508. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, P.C.; Palmer, N. Pathology of Domestic Animals; Academic Press: Cambridge, MA, USA, 2007. [Google Scholar]
- Clark, J.; Shotts Jr, E.; Hill, J.; McCall, J. Salmonellosis in gerbils induced by nonrelated experimental procedure. Lab. Anim. Sci. 1992, 42, 161–163. [Google Scholar] [PubMed]
- Constable, P.D.; Hinchcliff, K.W.; Done, S.H.; Grünberg, W. Veterinary Medicine-E-Book: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats; Elsevier Health Sciences: New York, NY, USA, 2017. [Google Scholar]
- Davies, R.; Evans, S.; Chappell, S.; Kidd, S.; Jones, Y.; Preece, B. Increase in Salmonella enterica Subspecies Diarizonae Serovar 61: K: L, 5,(7) in Sheep. Veterinary Record 2001, 149, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Ladds, P. The male genital system. In Pathology of Domestic Animals, 4th ed.; Jubb, K.V.F., Kennedy, P.C., Palmer, N., Eds.; Elsevier Academic Press: San Diego, CA, USA, 1993; pp. 471–529. [Google Scholar]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vazquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.I.; Bartlett, J.A.; Corey, G.R. Extra-intestinal manifestations of salmonella infections. Medicine 1987, 66, 349–388. [Google Scholar] [CrossRef]
- Greenfield, J.; Greenway, J.; Bigland, C. Arizona infections in sheep associated with gastroenteritis and abortion. Vet. Record 1973, 92, 400. [Google Scholar] [CrossRef]
- Kostalas, G.; Thomas, E. Orchitis and testicular abscess formation as complications of Salmonella virchow gastroenteritis. Med. J. Aust. 1973, 2, 769–770. [Google Scholar] [CrossRef]
- Handrick, W.; Bergmann, L.; Spencker, F.; Bennek, J. Orchitis and epididymitis caused by Salmonella Cholerae suis in a premature infant. Z. Urol. Nephrol. 1987, 80, 89–91. [Google Scholar]
- Rabsch, W.; Andrews, H.L.; Kingsley, R.A.; Prager, R.; Tschäpe, H.; Adams, L.G.; Bäumler, A.J. Salmonella enterica serotype Typhimurium and its host-adapted variants. Infect. Immun. 2002, 70, 2249–2255. [Google Scholar] [CrossRef] [Green Version]
- Rosche, K.L.; Aljasham, A.T.; Kipfer, J.N.; Piatkowski, B.T.; Konjufca, V. Infection with Salmonella enterica serovar Typhimurium leads to increased proportions of F4/80+ red pulp macrophages and decreased proportions of B and T lymphocytes in the spleen. PLoS ONE 2015, 10, e0130092. [Google Scholar] [CrossRef]
- Borges, K.A.; Furian, T.Q.; Borsoi, A.; Moraes, H.L.; Salle, C.T.; Nascimento, V.P. Detection of virulence-associated genes in Salmonella Enteritidis isolates from chicken in South of Brazil. Pesqui. Vet. Bras. 2013, 33, 1416–1422. [Google Scholar] [CrossRef]
- Pullinger, G.D.; Paulin, S.M.; Charleston, B.; Watson, P.R.; Bowen, A.J.; Dziva, F.; Morgan, E.; Villarreal-Ramos, B.; Wallis, T.S.; Stevens, M.P. Systemic translocation of Salmonella enterica serovar Dublin in cattle occurs predominantly via efferent lymphatics in a cell-free niche and requires type III secretion system 1 (T3SS-1) but not T3SS-2. Infect. Immun. 2007, 75, 5191–5199. [Google Scholar] [CrossRef] [Green Version]
- Salcedo, S.P.; Noursadeghi, M.; Cohen, J.; Holden, D.W. Intracellular replication of Salmonella Typhimurium strains in specific subsets of splenic macrophages in vivo. Cell. Microbiol. 2001, 3, 587–597. [Google Scholar] [CrossRef]
- Elkenany, R.; Elsayed, M.M.; Zakaria, A.I.; El-sayed, S.A.-E.-S.; Rizk, M.A. Antimicrobial resistance profiles and virulence genotyping of Salmonella enterica serovars recovered from broiler chickens and chicken carcasses in Egypt. BMC Vet. Res. 2019, 15, 124. [Google Scholar] [CrossRef] [Green Version]
- Prager, R.; Rabsch, W.; Streckel, W.; Voigt, W.; Tietze, E.; Tschäpe, H. Molecular properties of Salmonella enterica serotype Paratyphi B distinguish between its systemic and its enteric pathovars. J. Clin. Microbiol. 2003, 41, 4270–4278. [Google Scholar] [CrossRef] [Green Version]
- Ammar, A.M.; Mohamed, A.A.; Abd El-Hamid, M.I.; El-Azzouny, M.M. Virulence genotypes of clinical Salmonella Serovars from broilers in Egypt. J. Infect. Dev. Ctries 2016, 10, 337–346. [Google Scholar] [CrossRef] [Green Version]
- Muthu, G.; Suresh, A.; VishnuPrabu, D.; Munirajan, A.; ESTHER, M.S.; Sathishkumar, E.; Gopinath, P.; Srivani, S. Detection of virulence genes from Salmonella species in Chennai, India. CIB Technol. J. Microbiol. 2014, 3, 11–14. [Google Scholar]
- Barilli, E.; Bacci, C.; StellaVilla, Z.; Merialdi, G.; D’Incau, M.; Brindani, F.; Vismarra, A. Antimicrobial resistance, biofilm synthesis and virulence genes in Salmonella isolated from pigs bred on intensive farms. Ital. J. Food Saf. 2018, 7, 7223. [Google Scholar] [CrossRef]
- Poester, F.; Samartino, L.; Santos, R. Pathogenesis and pathobiology of brucellosis in livestock. Rev. Sci. Technol. 2013, 32, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Rankin, J. Brucella abortus in bulls: A study of twelve naturally infected cases. Vet. Record 1965, 77, 132–135. [Google Scholar]
- Campero, C.M.; Ladds, P.; Hoffmann, D.; Duffield, B.; Watson, D.; Fordyce, G. Immunopathology of experimental Brucella abortus strain 19 infection of the genitalia of bulls. Vet. Immunol. Immunopathol. 1990, 24, 235–246. [Google Scholar] [CrossRef]
- Jansen, B. The Epidemiology of Bacterial Infection of the Genitalia in Rams. Onderstepoort J. Vet. Res. 1983, 50, 275–282. [Google Scholar]
- Foster, R.; Weber, T.R.; Kleiman, M.; Grosfeld, J.L. Salmonella Enteritidis: Testicular abscess in a newborn. J. Urol. 1983, 130, 790–791. [Google Scholar] [CrossRef]
- Adem, J.; Bushra, E. Bovine salmonellosis and its public health importance: A review. Adv. Life Sci. Technol. 2016, 44, 62–71. [Google Scholar]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef]
- Cesta, M.F. Normal structure, function, and histology of the spleen. Toxicol. Pathol. 2006, 34, 455–465. [Google Scholar] [CrossRef]
- Den Haan, J.M.; Kraal, G. Innate immune functions of macrophage subpopulations in the spleen. J. Innate Immun. 2012, 4, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Koury, M.J.; Sawyer, S.T.; Brandt, S.J. New insights into erythropoiesis. Curr. Opin. Hematol. 2002, 9, 93–100. [Google Scholar] [CrossRef]
- Mastroeni, P.; Skepper, J.; Hormaeche, C. Effect of anti-tumor necrosis factor alpha antibodies on histopathology of primary Salmonella infections. Infect. Immun. 1995, 63, 3674–3682. [Google Scholar] [CrossRef] [Green Version]
- Shilangale, R.P.; Kaaya, G.P.; Chimwamurombe, P.M. Prevalence and characterization of Salmonella isolated from beef in Namibia. Eur. J. Nutr. Food Saf. 2015, 5, 267–274. [Google Scholar] [CrossRef]
- Pincus, D.H. Microbial identification using the bioMérieux Vitek® 2 system. In Encyclopedia of Rapid Microbiological Methods; Parenteral Drug Association: Bethesda, MD, USA, 2006; pp. 1–32. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Murugkar, H.; Rahman, H.; Dutta, P. Distribution of virulence genes in Salmonella serovars isolated from man & animals. Indian J. Med. Res. 2003, 117, 66–70. [Google Scholar]
- Huehn, S.; La Ragione, R.M.; Anjum, M.; Saunders, M.; Woodward, M.J.; Bunge, C.; Helmuth, R.; Hauser, E.; Guerra, B.; Beutlich, J. Virulotyping and antimicrobial resistance typing of Salmonella enterica serovars relevant to human health in Europe. Foodborne Pathog. Dis. 2010, 7, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, X.; Zhang, L.; Liu, W.; Dan, X.; Shi, C.; Shi, X. Development of a novel multiplex PCR assay for the identification of Salmonella enterica Typhimurium and Enteritidis. Food Control 2012, 27, 87–93. [Google Scholar] [CrossRef]
- Akbarmehr, J.; Salehi, T.Z.; Brujeni, G. Identification of Salmonella isolated from poultry by MPCR technique and evaluation of their hsp groEL gene diversity based on the PCR-RFLP analysis. Afr. J. Microbiol. Res. 2010, 4, 1594–1598. [Google Scholar]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, M.S.; Abd-Elhafeez, H.H.; Talukder, A.K.; Miyamoto, A. Ovulatory follicular fluid induces sperm phagocytosis by neutrophils, but oviductal fluid around oestrus suppresses its inflammatory effect in the buffalo oviduct in vitro. Mol. Reprod. Dev. 2019, 86, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.K. A formaldehyde glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 1A–149A. [Google Scholar]
- Abdel-Hakeem, S.S.; Mahmoud, G.A.-E.; Abdel-Hafeez, H.H. Evaluation and Microanalysis of Parasitic and Bacterial Agents of Egyptian Fresh Sushi, Salmo salar. Microsc. Microanal. 2019, 25, 1498–1508. [Google Scholar] [CrossRef]
- Suvarna, S.K. Bancroft’s Theory and Practice of Histological Techniques Consult; Churchill Livingstone, Elsevier: London, UK, 2013. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primers Sequences | Amplified Segment (bp) | Primary Denaturation | Amplification (35 cycles) | Final Extension | Reference | ||
|---|---|---|---|---|---|---|---|---|
| Secondary Denaturation | Annealing | Extension | ||||||
| 16S rRNA | 27F (AGAGTTTGATCCTGGCTCAG) | 1500 | 95 °C 5 min | 95 °C 60 s | 50 °C 60 s | 72 °C 90 s | 72 °C 10 min | Lane 1991 |
| 1492R (GGTTACCTTGTTACGACTT) | ||||||||
| Stn | F(TTG TGT CGC TAT CAC TGG CAA CC) | 617 | 94°C 5 min. | 94°C 30 s | 59°C 40 s | 72°C 45 s | 72 °C 10 min. | Murugkar et al., 2003 |
| R(ATT CGT AAC CCG CTC TCG TCC) | ||||||||
| sopB | F(TCAGAAGRCGTCTAACCACTC) | 517 | 94 °C 5 min. | 94 °C 30 s | 58 °C 40 s | 72 °C 45 s | 72 °C 10 min. | Huehn et al. 2010 |
| R(TAC CGTCCT CAT GCACACTC) | ||||||||
| avrA | F(CCT GTA TTG TTG AGC GTC TGG) | 422 | 94 °C 5 min. | 94 °C 30 s | 58 °C 40 s | 72 °C 50 s | 72 °C 10 min. | |
| R(AGA AGA GCT TCG TTG AAT GTC C) | ||||||||
| S. Typhimurium STM4495 | F(GGT GGC AAG GGA ATG AA) | 915 | 94 °C 5 min. | 94 °C 30 s | 50 °C 40 s | 72 °C 50 s | 72 °C 10 min. | Liu et al., 2012 |
| R(CGC AGC GTA AAG CAA CT) | ||||||||
| S. Enteritidis sefA | F(GCAGCGGTTACTATTGCAGC) | 310 | 94 °C 5 min. | 94 °C 30 s | 52 °C 30 s | 72 °C 30 s | 72 °C 7 min | Akbarmehr et al., 2010 |
| R(TGTGACAGGGACATTTAGCG) | ||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmoud, M.A.M.; Megahed, G.; Yousef, M.S.; Ali, F.A.Z.; Zaki, R.S.; Abdelhafeez, H.H. Salmonella Typhimurium Triggered Unilateral Epididymo-Orchitis and Splenomegaly in a Holstein Bull in Assiut, Egypt: A Case Report. Pathogens 2020, 9, 314. https://doi.org/10.3390/pathogens9040314
Mahmoud MAM, Megahed G, Yousef MS, Ali FAZ, Zaki RS, Abdelhafeez HH. Salmonella Typhimurium Triggered Unilateral Epididymo-Orchitis and Splenomegaly in a Holstein Bull in Assiut, Egypt: A Case Report. Pathogens. 2020; 9(4):314. https://doi.org/10.3390/pathogens9040314
Chicago/Turabian StyleMahmoud, Manal A. M., Gaber Megahed, Mohamed S. Yousef, Fatma Abo Zakaib Ali, Rania S. Zaki, and Hanan H. Abdelhafeez. 2020. "Salmonella Typhimurium Triggered Unilateral Epididymo-Orchitis and Splenomegaly in a Holstein Bull in Assiut, Egypt: A Case Report" Pathogens 9, no. 4: 314. https://doi.org/10.3390/pathogens9040314
APA StyleMahmoud, M. A. M., Megahed, G., Yousef, M. S., Ali, F. A. Z., Zaki, R. S., & Abdelhafeez, H. H. (2020). Salmonella Typhimurium Triggered Unilateral Epididymo-Orchitis and Splenomegaly in a Holstein Bull in Assiut, Egypt: A Case Report. Pathogens, 9(4), 314. https://doi.org/10.3390/pathogens9040314