Tripartite Interactions among Ixodiphagus hookeri, Ixodes ricinus and Deer: Differential Interference with Transmission Cycles of Tick-Borne Pathogens

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Prevalence of I. hookeri and Tick-Borne Pathogens in Different Life Stages of I. ricinus
2.2. Infestations with I. hookeri and Infection with Horizontally Transmitted Tick-Borne Pathogens
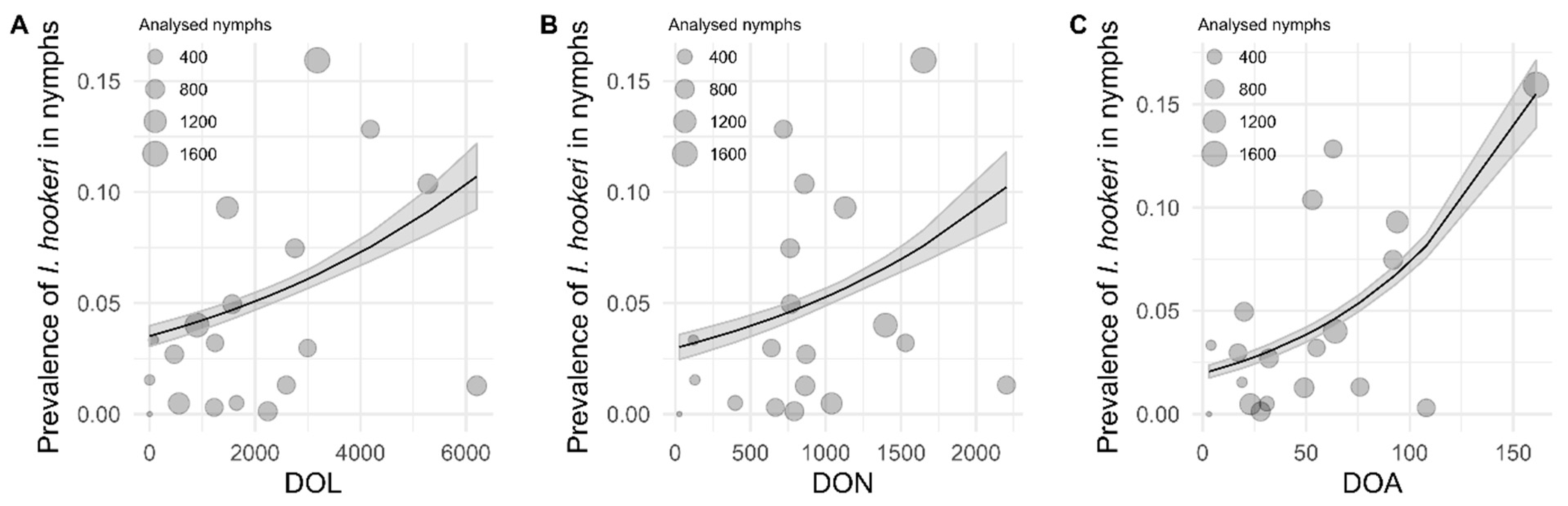
2.3. Association of I. hookeri Prevalence in Questing Nymphs with Density of I. ricinus
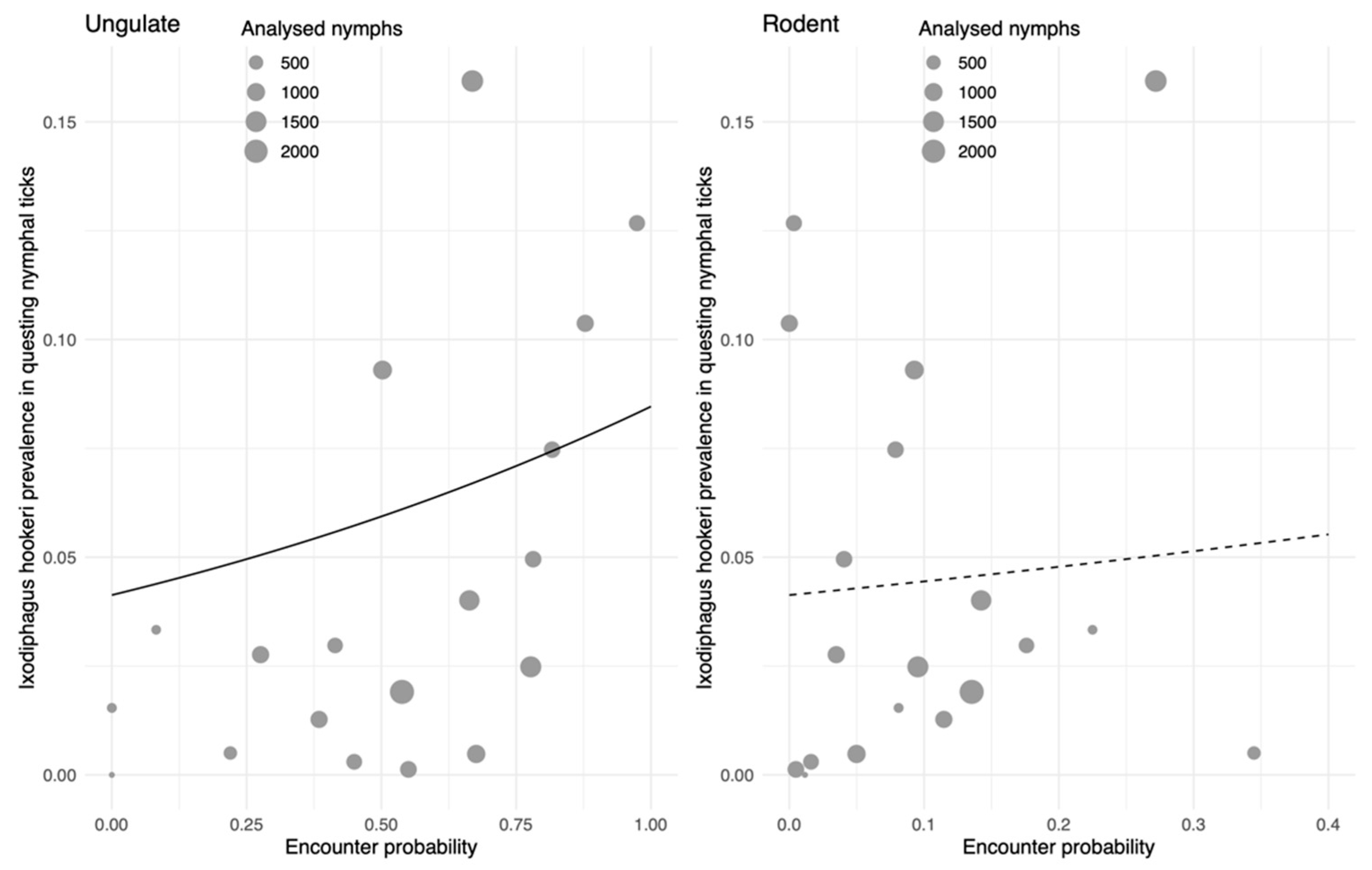
2.4. Association of I. hookeri with Densities of Ungulates and Rodents
2.5. Artificial Blood-Feeding of I. ricinus Nymphs
3. Discussion
4. Materials and Methods
4.1. Tick Collection
4.2. Cross-Sectional Study
4.3. Detection of I. hookeri and Tick-Borne Pathogens
4.4. Co-Infection Analysis
4.5. Association of I. hookeri Prevalence in Questing Nymphs with the Density of I. ricinus
4.6. Association of I. hookeri Prevalence with Vertebrate Encounter Probability
4.7. Artificial Blood-Feeding of I. ricinus Nymphs and Examination of Wasp Emergence
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Sprong, H.; Azagi, T.; Hoornstra, D.; Nijhof, A.M.; Knorr, S.; Baarsma, M.E.; Hovius, J.W. Control of Lyme borreliosis and other Ixodes ricinus-borne diseases. Parasites Vectors 2018, 11, 145. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L. The blacklegged tick, Ixodes scapularis: An increasing public health concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, E.; Andersson, Y.; Suk, J.E.; Sudre, B.; Semenza, J.C. Public health. Monitoring EU emerging infectious disease risk due to climate change. Science 2012, 336, 418–419. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Pena, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Matei, I.A.; Estrada-Pena, A.; Cutler, S.J.; Vayssier-Taussat, M.; Varela-Castro, L.; Potkonjak, A.; Zeller, H.; Mihalca, A.D. A review on the eco-epidemiology and clinical management of human granulocytic anaplasmosis and its agent in Europe. Parasites Vectors 2019, 12, 599. [Google Scholar] [CrossRef]
- Azagi, T.; Hoornstra, D.; Kremer, K.; Hovius, J.W.R.; Sprong, H. Evaluation of disease causality of rare Ixodes ricinus-borne infections in Europe. Pathogens 2020, 9, 150. [Google Scholar] [CrossRef]
- Hofhuis, A.; van de Kassteele, J.; Sprong, H.; van den Wijngaard, C.C.; Harms, M.G.; Fonville, M.; van Leeuwen, A.D.; Simoes, M.; van Pelt, W. Predicting the risk of Lyme borreliosis after a tick bite, using a structural equation model. PLoS ONE 2017, 12, e0181807. [Google Scholar] [CrossRef]
- Mannelli, A.; Boggiatto, G.; Grego, E.; Cinco, M.; Murgia, R.; Stefanelli, S.; De Meneghi, D.; Rosati, S. Acarological risk of exposure to agents of tick-borne zoonoses in the first recognized Italian focus of Lyme borreliosis. Epidemiol. Infect. 2003, 131, 1139–1147. [Google Scholar] [CrossRef]
- Coipan, E.C.; Jahfari, S.; Fonville, M.; Maassen, C.B.; van der Giessen, J.; Takken, W.; Takumi, K.; Sprong, H. Spatiotemporal dynamics of emerging pathogens in questing Ixodes ricinus. Front. Cell. Infect. Microbiol. 2013, 3, 36. [Google Scholar] [CrossRef]
- Randolph, S.E. Tick ecology: Processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors. Parasitology 2004, 129 (Suppl. 1), S37–S65. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Talleklint, L.; Lundqvist, L.; Olsen, B.; Chirico, J.; Mejlon, H. Geographical distribution, host associations, and vector roles of ticks (Acari: Ixodidae, Argasidae) in Sweden. J. Med. Entomol. 1994, 31, 240–256. [Google Scholar] [CrossRef]
- Takumi, K.; Sprong, H.; Hofmeester, T.R. Impact of vertebrate communities on Ixodes ricinus-borne disease risk in forest areas. Parasites Vectors 2019, 12, 434. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; van der Lei, P.B.; van Leeuwen, A.D.; Sprong, H.; van Wieren, S.E. New foci of Haemaphysalis punctata and Dermacentor reticulatus in the Netherlands. Ticks Tick Borne Dis. 2016, 7, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Sprong, H.; Jansen, P.A.; Prins, H.H.T.; van Wieren, S.E. Deer presence rather than abundance determines the population density of the sheep tick, Ixodes ricinus, in Dutch forests. Parasites Vectors 2017, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Rand, P.W.; Lubelczyk, C.; Lavigne, G.R.; Elias, S.; Holman, M.S.; Lacombe, E.H.; Smith, R.P., Jr. Deer density and the abundance of Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2003, 40, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.; Maffey, G.L.; Ramsay, S.L.; Hester, A.J. The effect of deer management on the abundance of Ixodes ricinus in Scotland. Ecol. Appl. 2012, 22, 658–667. [Google Scholar] [CrossRef]
- Ostfeld, R.S.; Schauber, E.M.; Canham, C.D.; Keesing, F.; Jones, C.G.; Wolff, J.O. Effects of acorn production and mouse abundance on abundance and Borrelia burgdorferi infection prevalence of nymphal Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001, 1, 55–63. [Google Scholar] [CrossRef]
- Krawczyk, A.I.; van Duijvendijk, G.L.A.; Swart, A.; Heylen, D.; Jaarsma, R.I.; Jacobs, F.H.H.; Fonville, M.; Sprong, H.; Takken, W. Effect of rodent density on tick and tick-borne pathogen populations: Consequences for infectious disease risk. Parasites Vectors 2020, 13, 34. [Google Scholar] [CrossRef]
- Vazquez, M.; Muehlenbein, C.; Cartter, M.; Hayes, E.B.; Ertel, S.; Shapiro, E.D. Effectiveness of personal protective measures to prevent Lyme disease. Emerg. Infect. Dis. 2008, 14, 210–216. [Google Scholar] [CrossRef]
- Eisen, L.; Dolan, M.C. Evidence for personal protective measures to reduce human contact with blacklegged ticks and for environmentally based control methods to suppress host-seeking blacklegged ticks and reduce infection with Lyme disease spirochetes in tick vectors and rodent reservoirs. J. Med. Entomol. 2016, 53, 1063–1092. [Google Scholar] [CrossRef] [PubMed]
- Foster, E.; Fleshman, A.C.; Ford, S.L.; Levin, M.L.; Delorey, M.J.; Eisen, R.J.; Eisen, L. Preliminary evaluation of human personal protective measures against the nymphal stage of the Asian longhorned tick (Acari: Ixodidae). J. Med. Entomol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Hyland, K.E.; Oliver, J.H. A review on the use of Ixodiphagus wasps (Hymenoptera: Encyrtidae) as natural enemies for the control of ticks (Acari:Ixodidae). Syst. Appl. Acarol. 1988, 3, 19–28. [Google Scholar] [CrossRef]
- Smith, C.N.; Cole, M.M. Studies of parasites of the American dog tick. J. Econ. Entomol. 1943, 36, 569–572. [Google Scholar] [CrossRef]
- Mwangi, E.N.; Hassan, S.M.; Kaaya, G.P.; Essuman, S. The impact of Ixodiphagus hookeri, a tick parasitoid, on Amblyomma variegatum (Acari: Ixodidae) in a field trial in Kenya. Exp. Appl. Acarol. 1997, 21, 117–126. [Google Scholar] [CrossRef]
- Hu, R.; Hyland, K.E. Effects of the feeding process of Ixodes scapularis (Acari: Ixodidae) on embryonic development of its parasitoid, Ixodiphagus hookeri (Hymenoptera: Encyrtidae). J. Med. Entomol. 1998, 35, 1050–1053. [Google Scholar] [CrossRef]
- Collatz, J.; Selzer, P.; Fuhrmann, A.; Oehme, R.M.; Mackenstedt, U.; Kahl, O.; Steidle, J.L.M. A hidden beneficial: Biology of the tick-wasp Ixodiphagus hookeri in Germany. J. Appl. Entomol. 2011, 135, 351–358. [Google Scholar] [CrossRef]
- Larrousse, F.; King, A.G.; Wolbach, S.B. The over-wintering in Massachusetts of Ixodiphagous caucurteri. Science 1928, 67, 351–353. [Google Scholar] [CrossRef]
- Plantard, O.; Bouju-Albert, A.; Malard, M.A.; Hermouet, A.; Capron, G.; Verheyden, H. Detection of Wolbachia in the tick Ixodes ricinus is due to the presence of the hymenoptera endoparasitoid Ixodiphagus hookeri. PLoS ONE 2012, 7, e30692. [Google Scholar] [CrossRef]
- Wood, H.P. Notes on the life history of the tick parasite Hunterellus hookeri Howard. J. Econ. Entomol. 1911, 4, 425–431. [Google Scholar] [CrossRef]
- Tijsse-Klasen, E.; Braks, M.; Scholte, E.J.; Sprong, H. Parasites of vectors--Ixodiphagus hookeri and its Wolbachia symbionts in ticks in The Netherlands. Parasites Vectors 2011, 4, 228. [Google Scholar] [CrossRef] [PubMed]
- Collatz, J.; Fuhrmann, A.; Selzer, P.; Oehme, R.M.; Hartelt, K.; Kimmig, P.; Meiners, T.; Mackenstedt, U.; Steidle, J.L.M. Being a parasitoid of parasites: Host finding in the tick wasp Ixodiphagus hookeri by odours from mammals. Entomol. Exp. Appl. 2010, 134, 131–137. [Google Scholar] [CrossRef]
- Ramos, R.A.; Campbell, B.E.; Whittle, A.; Lia, R.P.; Montarsi, F.; Parisi, A.; Dantas-Torres, F.; Wall, R.; Otranto, D. Occurrence of Ixodiphagus hookeri (Hymenoptera: Encyrtidae) in Ixodes ricinus (Acari: Ixodidae) in southern Italy. Ticks Tick Borne Dis. 2015, 6, 234–236. [Google Scholar] [CrossRef] [PubMed]
- Bohacsova, M.; Mediannikov, O.; Kazimirova, M.; Raoult, D.; Sekeyova, Z. Arsenophonus nasoniae and Rickettsiae infection of Ixodes ricinus due to parasitic wasp Ixodiphagus hookeri. PLoS ONE 2016, 11, e0149950. [Google Scholar] [CrossRef] [PubMed]
- Sormunen, J.J.; Sippola, E.; Kaunisto, K.M.; Vesterinen, E.J.; Saaksjarvi, I.E. First evidence of Ixodiphagus hookeri (Hymenoptera: Encyrtidae) parasitization in Finnish castor bean ticks (Ixodes ricinus). Exp. Appl. Acarol. 2019, 79, 395–404. [Google Scholar] [CrossRef]
- Hofmeester, T.R.; Jansen, P.A.; Wijnen, H.J.; Coipan, E.C.; Fonville, M.; Prins, H.H.T.; Sprong, H.; van Wieren, S.E. Cascading effects of predator activity on tick-borne disease risk. Proc. Biol. Sci. 2017, 284, 20170453. [Google Scholar] [CrossRef]
- Armitage, P.; Berry, G.; Matthews, J.N.S. Statistical Methods in Medical Research, 4th ed.; Blackwell Publishers: Malden, MA, USA, 2001; p. 817. [Google Scholar]
- Demas, F.A.; Hassanali, A.; Mwangi, E.N.; Kunjeku, E.C.; Mabveni, A.R. Cattle and Amblyomma variegatum odors used in host habitat and host finding by the tick parasitoid, Ixodiphagus hookeri. J. Chem. Ecol. 2000, 26, 1079–1093. [Google Scholar] [CrossRef]
- Hu, R.; Hyland, K.E. Prevalence and seasonal activity of the wasp parasitoid, Ixodiphagus hookeri (Hymenoptera: Encyrtidae) in its tick host, Ixodes scapularis (Acari: Ixodidae). Syst. Appl. Acarol. 1997, 2, 95–100. [Google Scholar] [CrossRef]
- Stafford, K.C., 3rd; Denicola, A.J.; Kilpatrick, H.J. Reduced abundance of Ixodes scapularis (Acari: Ixodidae) and the tick parasitoid Ixodiphagus hookeri (Hymenoptera: Encyrtidae) with reduction of white-tailed deer. J. Med. Entomol. 2003, 40, 642–652. [Google Scholar] [CrossRef]
- Hu, R.; Hyland, K.E.; Mather, T.N. Occurrence and distribution in Rhode Island of Hunterellus hookeri (Hymenoptera: Encyrtidae), a wasp parasitoid of Ixodes dammini. J. Med. Entomol. 1993, 30, 277–280. [Google Scholar] [CrossRef]
- Tagliapietra, V.; Rosa, R.; Arnoldi, D.; Cagnacci, F.; Capelli, G.; Montarsi, F.; Hauffe, H.C.; Rizzoli, A. Saturation deficit and deer density affect questing activity and local abundance of Ixodes ricinus (Acari, Ixodidae) in Italy. Vet. Parasitol. 2011, 183, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Takasu, K.; Takano, S.; Sasaki, M.; Yagi, S.; Nakamura, S. Host recognition by the tick parasitoid Ixodiphagus hookeri (Hymenoptera: Encyrtidae). Environ. Entomol. 2003, 32, 614–617. [Google Scholar] [CrossRef][Green Version]
- Hofmeester, T.R.; Coipan, C.; van Wieren, S.; Prins, H.H.T.; Takken, W.; Sprong, H. Few vertebrate species dominate the Borrelia burgdorferi s.l. life cycle. Environ. Res. Lett. 2016, 11, 043001. [Google Scholar] [CrossRef]
- Hartemink, N.A.; Randolph, S.E.; Davis, S.A.; Heesterbeek, J.A. The basic reproduction number for complex disease systems: Defining R(0) for tick-borne infections. Am. Nat. 2008, 171, 743–754. [Google Scholar] [CrossRef] [PubMed]
- van Duijvendijk, G.; Coipan, C.; Wagemakers, A.; Fonville, M.; Ersoz, J.; Oei, A.; Foldvari, G.; Hovius, J.; Takken, W.; Sprong, H. Larvae of Ixodes ricinus transmit Borrelia afzelii and B. miyamotoi to vertebrate hosts. Parasites Vectors 2016, 9, 97. [Google Scholar] [CrossRef]
- Jahfari, S.; Coipan, E.C.; Fonville, M.; van Leeuwen, A.D.; Hengeveld, P.; Heylen, D.; Heyman, P.; van Maanen, C.; Butler, C.M.; Foldvari, G.; et al. Circulation of four Anaplasma phagocytophilum ecotypes in Europe. Parasites Vectors 2014, 7, 365. [Google Scholar] [CrossRef] [PubMed]
- Stuen, S.; Granquist, E.G.; Silaghi, C. Anaplasma phagocytophilum—A widespread multi-host pathogen with highly adaptive strategies. Front. Cell. Infect. Microbiol. 2013, 3, 31. [Google Scholar] [CrossRef]
- Hamsikova, Z.; Silaghi, C.; Takumi, K.; Rudolf, I.; Gunar, K.; Sprong, H.; Kazimirova, M. Presence of roe deer affects the occurrence of Anaplasma phagocytophilum ecotypes in questing Ixodes ricinus in different habitat types of Central Europe. Int. J. Environ. Res. Public Health 2019, 16, 4725. [Google Scholar] [CrossRef]
- Mather, T.N.; Piesman, J.; Spielman, A. Absence of spirochaetes (Borrelia burgdorferi) and piroplasms (Babesia microti) in deer ticks (Ixodes dammini) parasitized by chalcid wasps (Hunterellus hookeri). Med. Vet. Entomol. 1987, 1, 3–8. [Google Scholar] [CrossRef]
- Hanincova, K.; Schafer, S.M.; Etti, S.; Sewell, H.S.; Taragelova, V.; Ziak, D.; Labuda, M.; Kurtenbach, K. Association of Borrelia afzelii with rodents in Europe. Parasitology 2003, 126, 11–20. [Google Scholar] [CrossRef]
- Coipan, C.E.; van Duijvendijk, G.L.A.; Hofmeester, T.R.; Takumi, K.; Sprong, H. The genetic diversity of Borrelia afzelii is not maintained by the diversity of the rodent hosts. Parasites Vectors 2018, 11, 454. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Vilchez, I.; Mateos, M. Spiroplasma bacteria enhance survival of Drosophila hydei attacked by the parasitic wasp Leptopilina heterotoma. PLoS ONE 2010, 5, e12149. [Google Scholar] [CrossRef] [PubMed]
- Bhide, M.R.; Travnicek, M.; Levkutova, M.; Curlik, J.; Revajova, V.; Levkut, M. Sensitivity of Borrelia genospecies to serum complement from different animals and human: A host-pathogen relationship. FEMS Immunol. Med. Microbiol. 2005, 43, 165–172. [Google Scholar] [CrossRef]
- Chen, G.; Severo, M.S.; Sakhon, O.S.; Choy, A.; Herron, M.J.; Felsheim, R.F.; Wiryawan, H.; Liao, J.; Johns, J.L.; Munderloh, U.G.; et al. Anaplasma phagocytophilum dihydrolipoamide dehydrogenase 1 affects host-derived immunopathology during microbial colonization. Infect. Immun. 2012, 80, 3194–3205. [Google Scholar] [CrossRef]
- Hillyard, P.D. Ticks of North-West Europe: Keys and Notes for Identification of the Species; Field Studies Council: Shrewsbury, UK, 1996. [Google Scholar]
- Wielinga, P.R.; Gaasenbeek, C.; Fonville, M.; de Boer, A.; de Vries, A.; Dimmers, W.; Akkerhuis Op Jagers, G.; Schouls, L.M.; Borgsteede, F.; van der Giessen, J.W. Longitudinal analysis of tick densities and Borrelia, Anaplasma, and Ehrlichia infections of Ixodes ricinus ticks in different habitat areas in The Netherlands. Appl. Environ. Microbiol. 2006, 72, 7594–7601. [Google Scholar] [CrossRef]
- Heylen, D.; Tijsse, E.; Fonville, M.; Matthysen, E.; Sprong, H. Transmission dynamics of Borrelia burgdorferi s.l. in a bird tick community. Environ. Microbiol. 2013, 15, 663–673. [Google Scholar] [CrossRef]
- Jahfari, S.; Fonville, M.; Hengeveld, P.; Reusken, C.; Scholte, E.J.; Takken, W.; Heyman, P.; Medlock, J.M.; Heylen, D.; Kleve, J.; et al. Prevalence of Neoehrlichia mikurensis in ticks and rodents from North-west Europe. Parasites Vectors 2012, 5, 74. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Version 3.6.1 “Action of the Toes”; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- R Core Team. R Studio: Integrated Development for R; 0.99.903; R Studio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Oliver, J.D.; Lynn, G.E.; Burkhardt, N.Y.; Price, L.D.; Nelson, C.M.; Kurtti, T.J.; Munderloh, U.G. Infection of immature Ixodes scapularis (Acari: Ixodidae) by membrane feeding. J. Med. Entomol. 2016, 53, 409–415. [Google Scholar] [CrossRef]
- Krull, C.; Bohme, B.; Clausen, P.H.; Nijhof, A.M. Optimization of an artificial tick feeding assay for Dermacentor reticulatus. Parasites Vectors 2017, 10, 60. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Symbiont | Larvae (n = 367) | Nymphs (n = 684) | Adults (n = 78) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| n | % | (Range) | n | % | (Range) | n | % | (Range) | |
| I. hookeri % | 2 | 0.5% | (0.1–2.5) | 30 | 4.4% | (3.0–6.2) | 0 | 0% | (0.0–4.6) |
| B. burgdorferi s.l. % | 1 | 0.3% | (0.0–1.5) | 82 | 12% | (9.6–14.7) | 13 | 16.7% | (9.2–26.8) |
| A. phagocytophilum % | 0 | 0% | (0.0–1.0) | 19 | 2.8% | (1.7–4.3) | 7 | 9% | (3.7–17.6) |
| N. mikurensis % | 0 | 0% | (0.0–1.0) | 30 | 4.4% | (3.0–6.2) | 4 | 5.1% | (1.4–12.6) |
| A. phagocytophilum | B. afzelii | N. mikurensis | |
|---|---|---|---|
| Observed co-occurrence | 72 | 4 | 9 |
| Expected co-occurrence | 26 | 17 | 46 |
| Odds ratio | 3.3 | 0.2 | 0.2 |
| p-value | <0.001 | <0.001 | <0.001 |
| Ticks | I. hookeri % | A. phagocytophilum % | B. burgdorferi s.l. % | N. mikurensis % | |||||
|---|---|---|---|---|---|---|---|---|---|
| Female | (n = 64) | 7.8 | (2.6–17.3) | 15.6 | (7.8–26.9) | 0.0 | (0.0–5.6) | 0.0 | (0.0–5.6) |
| Male | (n = 87) | 5.7 | (1.9–12.9) | 19.5 | (11.8–29.4) | 0.0 | (0.0–4.2) | 0.0 | (0.0–4.2) |
| Not molted | (n = 14) | 64.3 | (35.1–87.2) | 14.3 | (1.8–42.8) | 0.0 | (0.0–23.2) | 0.0 | (0.0–23.2) |
| With wasps | (n = 7) | 100 | (59.0–100.0) | 28.6 | (3.7–71.0) | 0.0 | (0.0–41.0) | 0.0 | (0.0–41.0) |
| Total | (n = 172) | 15.1 | (10.1–21.4) | 18.0 | (12.6–24.6) | 0.0 | (0.0–2.1) | 0.0 | (0.0–2.1) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyk, A.I.; Bakker, J.W.; Koenraadt, C.J.M.; Fonville, M.; Takumi, K.; Sprong, H.; Demir, S. Tripartite Interactions among Ixodiphagus hookeri, Ixodes ricinus and Deer: Differential Interference with Transmission Cycles of Tick-Borne Pathogens. Pathogens 2020, 9, 339. https://doi.org/10.3390/pathogens9050339
Krawczyk AI, Bakker JW, Koenraadt CJM, Fonville M, Takumi K, Sprong H, Demir S. Tripartite Interactions among Ixodiphagus hookeri, Ixodes ricinus and Deer: Differential Interference with Transmission Cycles of Tick-Borne Pathogens. Pathogens. 2020; 9(5):339. https://doi.org/10.3390/pathogens9050339
Chicago/Turabian StyleKrawczyk, Aleksandra I., Julian W. Bakker, Constantianus J. M. Koenraadt, Manoj Fonville, Katsuhisa Takumi, Hein Sprong, and Samiye Demir. 2020. "Tripartite Interactions among Ixodiphagus hookeri, Ixodes ricinus and Deer: Differential Interference with Transmission Cycles of Tick-Borne Pathogens" Pathogens 9, no. 5: 339. https://doi.org/10.3390/pathogens9050339
APA StyleKrawczyk, A. I., Bakker, J. W., Koenraadt, C. J. M., Fonville, M., Takumi, K., Sprong, H., & Demir, S. (2020). Tripartite Interactions among Ixodiphagus hookeri, Ixodes ricinus and Deer: Differential Interference with Transmission Cycles of Tick-Borne Pathogens. Pathogens, 9(5), 339. https://doi.org/10.3390/pathogens9050339