Genetic Diversity of Bovine Hemoprotozoa in South Korea


Abstract
:1. Introduction
2. Results
2.1. PCR and Molecular Identification
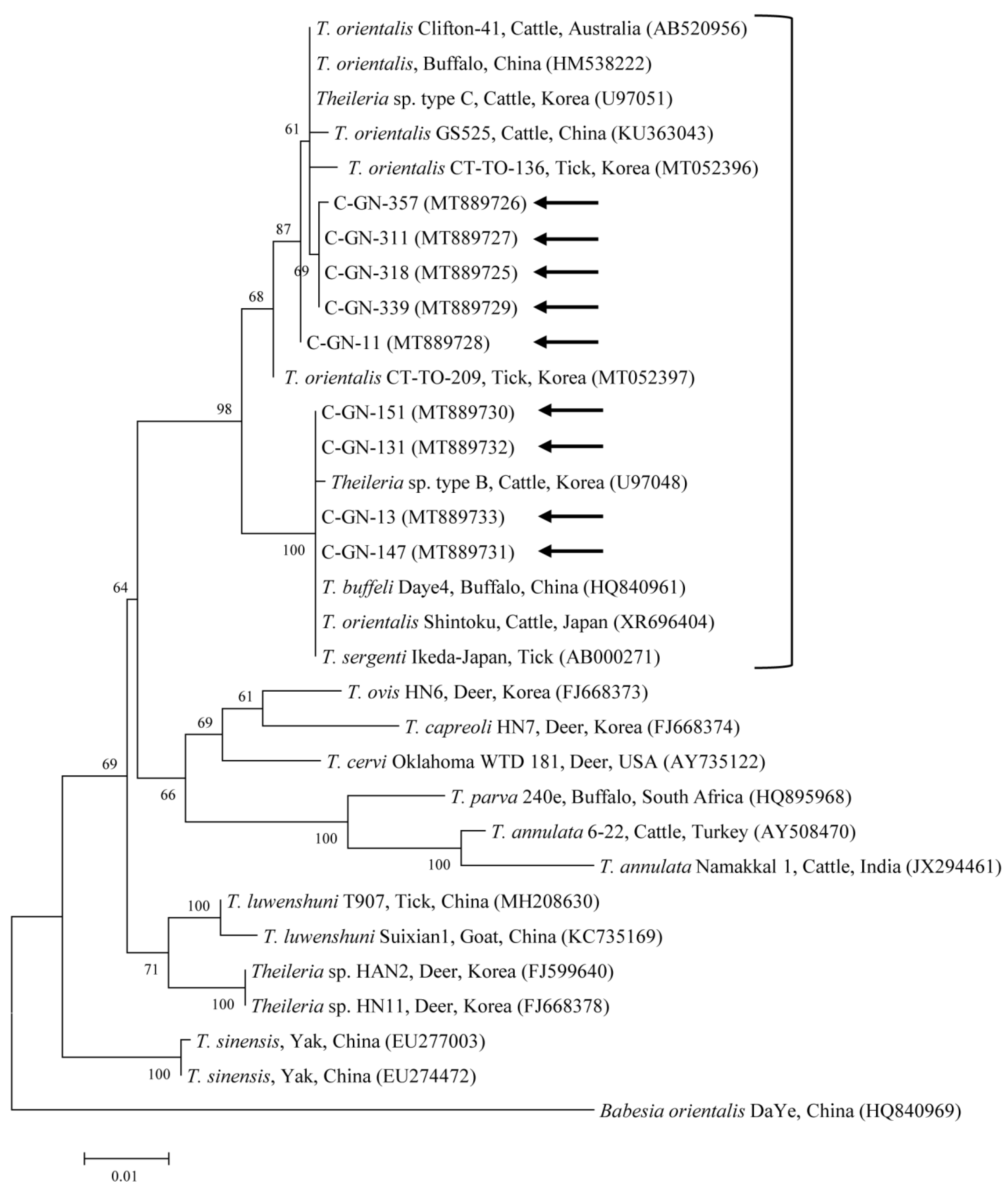
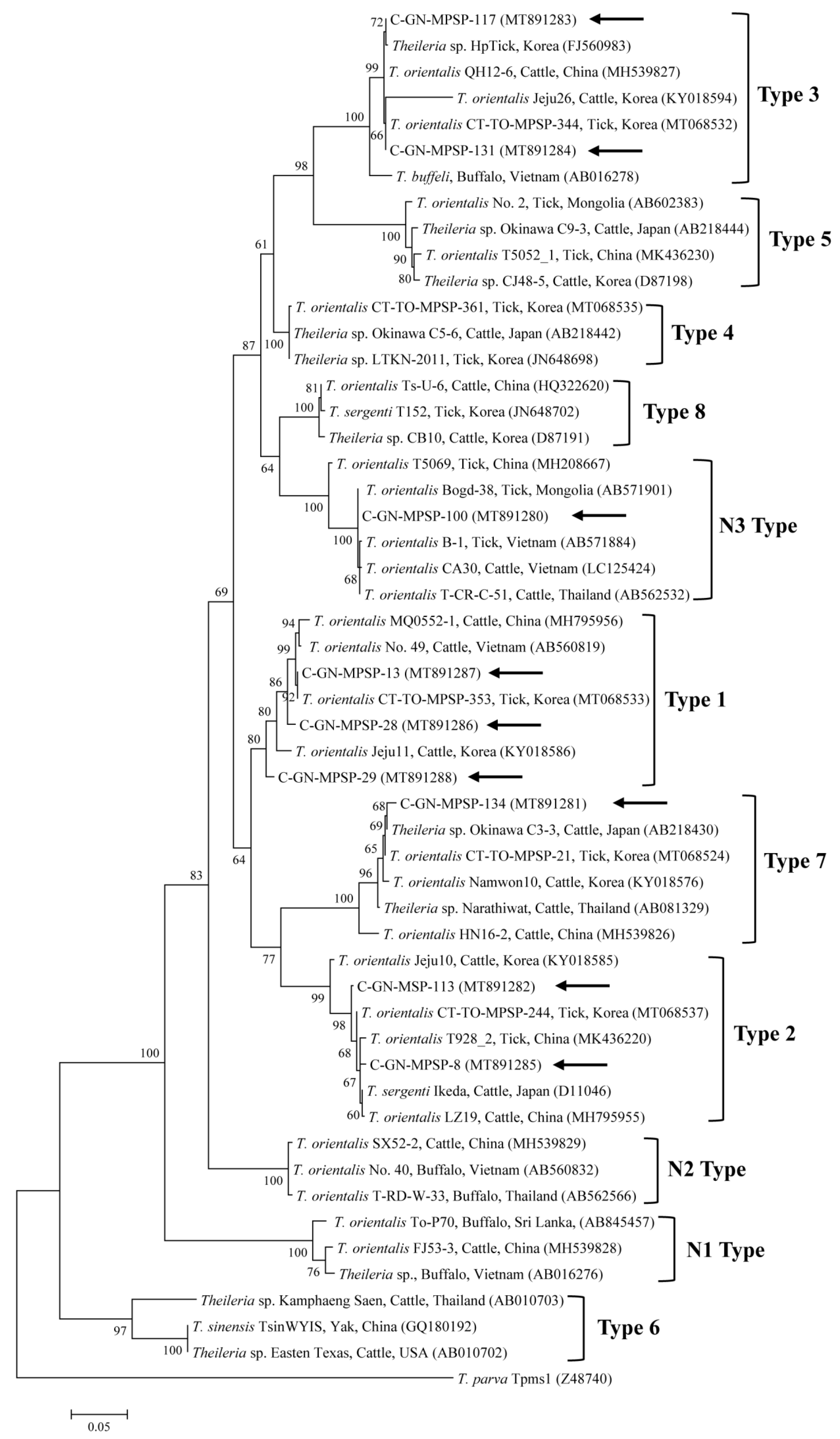
2.2. Molecular Analyses
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Samples
4.3. DNA Extraction and PCR
4.4. DNA Cloning
4.5. DNA Sequencing and Phylogenetic Analysis
4.6. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vayssier-Taussat, M.; Cosson, J.F.; Degeilh, B.; Eloit, M.; Fontanet, A.; Moutailler, S.; Raoult, D.; Sellal, E.; Ungeheuer, M.N.; Zylbermann, P. How a multidisciplinary ’One Health’ approach can combat the tick-borne pathogen threat in Europe. Future Microbiol. 2015, 10, 809–818. [Google Scholar] [CrossRef]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. International collaborative research: Significance of tick-borne hemoparasitic diseases to world animal health. Vet. Parasitol. 1995, 57, 19–41. [Google Scholar] [CrossRef]
- Sivakumar, T.; Hayashida, K.; Sugimoto, C.; Yokoyama, N. Evolution and genetic diversity of Theileria. Infect. Genet. Evol. 2014, 27, 250–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, M.; Altay, K.; Dumanli, N. A molecular survey of bovine Theileria parasites among apparently healthy cattle and with a note on the distribution of ticks in eastern Turkey. Vet. Parasitol. 2006, 138, 179–185. [Google Scholar] [CrossRef]
- Kamau, J.; de Vos, A.J.; Playford, M.; Salim, B.; Kinyanjui, P.; Sugimoto, C. Emergence of new types of Theileria orientalis in Australian cattle and possible cause of theileriosis outbreaks. Parasit. Vectors 2011, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Zhao, S.; Xie, S.; Li, H.; Wang, H.; Zhang, S. Molecular prevalence of Theileria infections in cattle in Yanbian, north-eastern China. Parasite 2020, 27, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Yu, D.H.; Kang, S.W.; Chae, J.B.; Choi, K.S.; Kim, H.C.; Park, B.K.; Chae, J.S.; Park, J. Hematological changes associated with Theileria orientalis infection in Korean indigenous cattle. Korean J. Parasitol. 2017, 55, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Han, Y.J.; Han, D.G.; Chae, J.B.; Chae, J.S.; Yu, D.H.; Lee, Y.S.; Park, B.K.; Kim, H.C.; Choi, K.S. Genetic characterization of Theileria orientalis from cattle in the Republic of Korea. Parasitol. Res. 2017, 116, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Chae, J.B.; Kim, S.; Yu, D.H.; Kim, H.C.; Park, B.K.; Chae, J.S.; Choi, K.S. Evaluation of the efficacy of ivermectin against Theileria orientalis infection in grazing cattle. BMC. Vet. Res. 2019, 15, 297. [Google Scholar]
- Seo, M.G.; Do, J.C.; Cho, M.H.; Seo, H.J.; Kim, J.K.; Kim, Y.H.; Park, N.C.; Kwak, D.M. Prevalence of Theileria sergenti infection in cattle of eastern areas in Gyeongbuk province by PCR. Korean J. Vet. Serv. 2011, 34, 251–258. [Google Scholar] [CrossRef]
- Ota, N.; Mizuno, D.; Kuboki, N.; Igarashi, I.; Nakamura, Y.; Yamashina, H.; Hanzaike, T.; Fujii, K.; Onoe, S.; Hata, H.; et al. Epidemiological survey of Theileria orientalis infection in grazing cattle in the eastern part of Hokkaido, Japan. J. Vet. Med. Sci. 2009, 71, 937–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khukhuu, A.; Lan, D.T.; Long, P.T.; Ueno, A.; Li, Y.; Luo, Y.; Macedo, A.C.; Matsumoto, K.; Inokuma, H.; Kawazu, S.; et al. Molecular epidemiological survey of Theileria orientalis in Thua Thien Hue Province, Vietnam. J. Vet. Med. Sci. 2011, 73, 701–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.W.; Doan, H.T.; Choe, S.E.; Noh, J.H.; Yoo, M.S.; Reddy, K.E.; Kim, Y.H.; Kweon, C.H.; Jung, S.C.; Chang, K.Y. Molecular investigation of tick-borne pathogens in ticks from grazing cattle in Korea. Parasitol. Int. 2013, 62, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Kwon, O.D.; Kwak, D. Genotypic Analysis of Piroplasms and Associated Pathogens from Ticks Infesting Cattle in Korea. Microorganisms 2020, 8, 728. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Yun, S.H.; Choi, S.K.; Cho, G.J.; Park, Y.S.; Cho, K.H.; Kwon, O.D.; Kwak, D. Molecular and phylogenetic analysis of equine piroplasms in the Republic of Korea. Res. Vet. Sci. 2013, 94, 579–583. [Google Scholar] [CrossRef]
- Choi, K.S.; Yu, D.H.; Chae, J.S.; Park, B.K.; Yoo, J.G.; Park, J. Seasonal changes in hemograms and Theileria orientalis infection rates among Holstein cattle pastured in the mountains in the Republic of Korea. Prev. Vet. Med. 2016, 127, 77–83. [Google Scholar] [CrossRef]
- Liu, A.; Guan, G.; Liu, Z.; Liu, J.; Leblanc, N.; Li, Y.; Gao, J.; Ma, M.; Niu, Q.; Ren, Q.; et al. Detecting and differentiating Theileria sergenti and Theileria sinensis in cattle and yaks by PCR based on major piroplasm surface protein (MPSP). Exp. Parasitol. 2010, 126, 476–481. [Google Scholar] [CrossRef]
- Kang, S.W.; Nguyen, L.T.; Noh, J.H.; Reddy, K.E.; Kweon, C.H.; Choe, S.E. Phylogenetic analysis of benign Theileria species based on major piroplasm surface protein (MPSP) genes from ticks of grazing cattle in Korea. Vet. Parasitol. 2012, 189, 145–152. [Google Scholar] [CrossRef]
- Li, L.H.; Wang, J.Z.; Zhu, D.; Li, X.S.; Lu, Y.; Yin, S.Q.; Li, S.G.; Zhang, Y.; Zhou, X.N. Detection of novel piroplasmid species and Babesia microti and Theileria orientalis genotypes in hard ticks from Tengchong County, Southwest China. Parasitol. Res. 2020, 119, 1259–1269. [Google Scholar] [CrossRef]
- Wang, J.; Yang, J.; Liu, J.; Wang, X.; Xu, J.; Liu, A.; Li, Y.; Liu, Z.; Ren, Q.; Luo, J.; et al. Molecular detection and genetic diversity of Theileria orientalis in cattle in China. Parasitol. Res. 2018, 117, 3689–3694. [Google Scholar] [CrossRef] [PubMed]
- Korean Statistical Information Service. Number of Farms in Households and Animals in Heads by Type. Available online: http://kosis.kr/statHtml/statHtml.do?orgId=101&tblId=DT_1EO099&conn_path=I2&language=en/ (accessed on 1 January 2020).
- Thrusfield, M. Veterinary Epidemiology, 3rd ed.; Blackwell Publishing: Oxford, UK, 2005. [Google Scholar]
- Kakuda, T.; Shiki, M.; Kubota, S.; Sugimoto, C.; Brown, W.C.; Kosum, C.; Nopporn, S.; Onuma, M. Phylogeny of benign Theileria species from cattle in Thailand, China and the U.S.A. based on the major piroplasm surface protein and small subunit ribosomal RNA genes. Int. J. Parasitol. 1998, 28, 1261–1267. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Category | No. Tested | No. Positive (%) | RR 1 | 95% CI 2 | p-Value | |
|---|---|---|---|---|---|---|
| Sex | Female | 327 | 8 (2.4) | 10.0 | 1.3–79.8 | 0.0293 * |
| Male | 410 | 1 (0.2) | 0.1 | 0.01–0.8 | 0.0293 * | |
| Age | <2 | 122 | 0 | 0.3 | 0.02–4.5 | 0.357 |
| 2–4 | 413 | 3 (0.7) | 0.4 | 0.1–1.6 | 0.1832 | |
| >4 | 202 | 6 (3.0) | 5.3 | 1.3–21.0 | 0.0176 * | |
| Breed | Hanwoo | 525 | 0 | 0.02 | 0–0.9 | 0.0079 * |
| Holstein | 212 | 9 (4.2) | 46.9 | 2.7–802.6 | 0.0079 * | |
| Region | Gyeongbuk | 361 | 0 | 0.1 | 0–0.9 | 0.0451 * |
| Gyeongnam | 376 | 9 (2.4) | 18.3 | 1.1–312.3 | 0.0451 * | |
| Season | Spring | 282 | 0 | 0.1 | 0–1.5 | 0.0886 |
| Summer | 178 | 9 (5.1) | 3.0 | 1.2–7.5 | 0.0166 * | |
| Autumn | 277 | 0 | 0.1 | 0–1.5 | 0.0924 | |
| Total | 737 | 9 (1.2) | 0.4–2.0 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwak, D.; Seo, M.-G. Genetic Diversity of Bovine Hemoprotozoa in South Korea. Pathogens 2020, 9, 768. https://doi.org/10.3390/pathogens9090768
Kwak D, Seo M-G. Genetic Diversity of Bovine Hemoprotozoa in South Korea. Pathogens. 2020; 9(9):768. https://doi.org/10.3390/pathogens9090768
Chicago/Turabian StyleKwak, Dongmi, and Min-Goo Seo. 2020. "Genetic Diversity of Bovine Hemoprotozoa in South Korea" Pathogens 9, no. 9: 768. https://doi.org/10.3390/pathogens9090768
APA StyleKwak, D., & Seo, M.-G. (2020). Genetic Diversity of Bovine Hemoprotozoa in South Korea. Pathogens, 9(9), 768. https://doi.org/10.3390/pathogens9090768