1. Introduction
Typical steppe is mainly distributed in arid and semi-arid areas, especially in Inner Mongolia, China, which serves as an essential component of the terrestrial ecosystem [
1], and plays a crucial role in promoting human well-being and providing sufficient nutrition to meet the demands of animal productions [
2]. Hay is a traditional management practice in the utilization of natural grassland [
3]. Generally, native grass hay is sufficient for animal feed during summer, but the supply can be limited in the autumn and winter when the forage grows slowly; therefore, it is hard to shake off an imbalance between the supply and demand of the ruminants [
4]. In addition, the quality and yield of forage were mainly affected by the dominant species in natural grassland [
5].
Stipa grandis was the dominant species of the typical steppe, and the application of which was limited due to the hard and sharp awn needles on the caryopses after maturity [
6]. Therefore, it is necessary to look for a suitable technique for
Stipa grandis to cover these shortages and promote its application in animal production.
Ensiling is a more practical way for forage preservation because of its fewer nutrient losses and good palatability [
7]. However, the absence of lactic acid bacteria (LAB) counts, water-soluble carbohydrate (WSC) content, and moisture, associated with the high buffering capacity, are factors that often hamper natural grasses ensilage [
8]. The inoculation of LAB has been proven to be a promising strategy to overcome this problem of poor fermentation [
9], which can rapidly reduce the pH value and inhibit the activity of undesirable microorganisms for improving the ensiling performance of silage [
10]. Zi et al. [
11] reported that the addition of LAB can increase the abundance of desirable members of the bacterial community and eliminate harmful bacteria. Nevertheless, the reconstitution mechanism of LAB during ensiling is complex [
12]. Fijałkowska et al. [
13] proved that only suitable LAB strains can be used to guarantee successful silage. A previous study has reported that the best LAB strains for promoting fermentation quality always source from the forage itself [
14]. LAB treatment has been isolated from multiple materials, including pineapple residue [
15], defective banana [
16], elephant silage [
17], and alfalfa [
18]. However, little knowledge is available on the LAB isolation and application from the dominant species of native grass, and to the best of our knowledge, there is no report on the
Stipa grandis in LAB isolation and application.
Therefore, the purpose of this study was to characterize the isolated LAB from the dominant species of native grass using phenotypic, chemotaxonomic characteristics, and molecular methods and evaluate their effects on the silage quality and bacterial community of Stipa grandis silage.
4. Discussion
The strategy of isolating novel LAB candidates in fermented forage has been widely used [
38]. However, it is challenging to differentiate the species according to the phenotyping procedures to assign isolates [
39]. The 16S rRNA sequence analysis method has proven to be highly effective in identifying the genus and species [
26]. In the present study, strains ZX301 and YX34 were identified by 16SrRNA gene sequencing, which was determined to be homofermentative, and belong to
Pediococcus pentosaceus and
Lactiplantibacillus plantarum, respectively. The result was consistent with the findings of Yang et al. [
40], who reported that the LAB genera, including
Lactobacillus and
Pediococcus, were the most frequently isolated microorganisms and exhibited a phylogenetically coherent treatment that clearly distinguishes them from other bacteria. Notably, the strains ZX301 and YX34 could grow normally at pH 3.5, which demonstrates an ability to grow in low pH environments of the strains and agree with the findings that homofermentative strains expressed high tolerance to a low-pH environment [
41]. Generally, the optimum growth and reproduction temperatures of LAB should be below 45 °C [
42]. In the present study, all strains could grow normally at temperatures of 10 °C or 30 °C, whereas strain ZX301 could weakly grow at 45 °C, which might be justified by the long-term evolution and natural selection on the unique environment of typical steppe [
14]. In addition, all strains could ferment arabinose, glucose, fructose, and cellobiose, which suggests that the isolated strains in this study could widely use various fermentation substrates. Therefore, the unique traits of the two selected strains offer a warrant for using these LAB in silage making.
The material characteristics of the forage could directly affect the silage quality. In the present study, the DM content of
Stipa grandis was 50.71%, consistent with the previously published [
43]. Generally, WSC content is critical for successful silage. The WSC content found in this study was slightly lower than the level of 5% suggested by Amer et al. [
44]. This study had the opposite result of Chiy et al. [
45], who reported that Graminaceous species generally contain higher WSC content, likely as a result of prolonged drought stress in typical grassland, which could increase fructan hydrolysis and reduce the WSC content. However, the result of this study agrees with Hou et al. [
8], who found the native grass in typical steppe had a low WSC content. In addition, the result of this study showed that the CP ingredient was lower, while NDF and ADF ingredients were higher in
Stipa grandis, which agrees with Bu et al. [
46].
Generally, successful silage depends mainly on the amount and type of microorganisms attached to the FM [
47]. The previous study suggested that the optimum lactic acid bacteria content in raw materials should exceed 10
5 cfu/g [
48]. However, a lower content of LAB and higher numbers of undesirable microorganisms were found in FM in this study, indicating that direct ensilement would not be likely to succeed. Consequently, it is necessary to inhibit the growth of harmful bacteria during the early stages by using LAB additives for
Stipa grandis silage.
As expected, the WSC, ADF, and NDF content was reduced compared to the fresh
Stipa grandis after ensiling, mainly due to the conversion of available substrates to lactic acid by microbes related to the utilization of carbohydrates under anaerobic conditions [
49]. Additionally, the acid hydrolysis of the digestible cell wall fraction might decrease the ADF and NDF content [
50]. Notably, the inoculated silage treatments provided a benefit in preserving the WSC content relative to the CK treatment, which was in agreement with the research published by Sifeeldein et al. [
17], who found that the LAB inoculants could inhibit the loss of WSC fermented by speeding up the process of LA fermentation to reduce pH value rapidly. However, a stable DM content was observed between CK and inoculated silage treatments, which agrees with the report that a combination of
L. acidophilus and
L. plantarum leads to a comparable DM content with controls [
51]. The result might be because lactic acid fermentation predominantly occurred in all treatments, as is shown in
Table 5, which will result in minimal DM losses [
52]. Compared to the fresh material of
Stipa grandis, a reduction in the CP content was detected after ensiling, which might be due to the action of plant and microbial enzymes contributing to the conversion of plant protein to non-protein nitrogen (NPN). In addition, the absence of WSC in
Stipa grandis limited the production of lactic acid and the decrease in silage pH, and then failed to inhibit the activity of proteinase [
53]. Whereas, the CP content between CK and inoculated silage groups was stable, which is consistent with a previous study published by Muck et al. [
54], who observed that bacterial inoculants provided only a small benefit in inhibiting proteolysis. According to a previous report, protease exhibits optimal activity generally at the condition of a silage pH > 4.5 [
55]. This result might be due to the lower terminal pH (4.00–4.15) in silage of all treatments, which could limit the activity of proteolytic enzymes [
39].
The LA content, pH value, and NH
3-N level of silages could directly reflect the silage quality [
56]. As expected, the inoculants of LAB in
Stipa grandis silage increased the LA content and led to a lower pH value and NH
3-N content compared to the CK treatment in this study, all of which were within the recommended range provided by [
57]. This result might be due to the LAB stimulating the LA fermentation by utilizing soluble carbohydrates and then producing a large amount of lactic acid, which could continuously reduce the pH value, and then inhibit the activity of proteases from plants and microbes to reduce proteolysis [
58,
59,
60]. Similarly, Sucu et al. [
61] proved that the addition of lactic acid bacteria could rapidly reduce the pH value and NH
3-N content by increasing the LA content.
The Goods’ coverage of all samples exceeded 99%, indicating that the depth of sequencing was adequately captured for bacterial community analysis [
62]. Generally, forage after ensiling has low alpha diversity caused by the dominant lactobacillus, which can inhibit the growth of other microbes and reduce the diversity of the bacterial community [
63]. However, in the present study, the Shannon and Simpson indices were higher in the ZX301 treatment than that in FM and other inoculated silage treatments, which might be explained by the antagonistic activity between the dominant genera sourced from exogenous LAB and other bacteria, which weaken the advantage of the dominant genera (
Figure 2C), and then lead to higher α-diversity indices in ZX301 treatment [
64,
65].
The PCoA analysis based on the Weighted UniFrac distances was performed to visualize the similarity and distinction of bacterial communities among treatments. FM treatment was clearly separated from silages, indicating that ensiling influenced the bacterial community. This finding was in agreement with Zhao et al. [
66], who reported that significant differences in the bacterial community composition were detected before and after ensiling. In addition, the treatments of YX34 and ZX301 were also well separated from the CK treatment, which suggests that the addition of LAB reconstructs the microbial community.
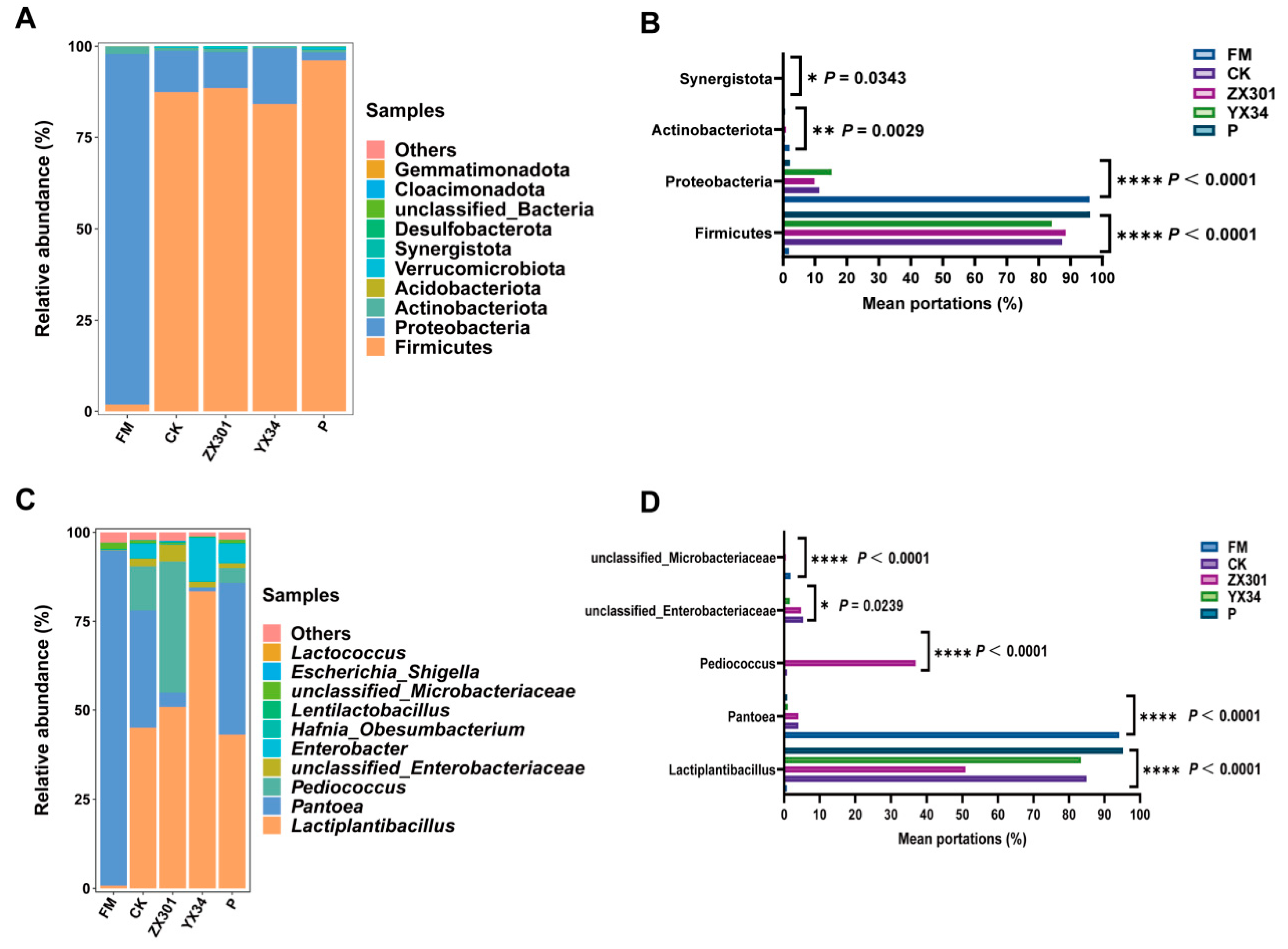
At the phylum level, the bacterial community in
Stipa grandis silage involves a shift from
Proteobacteria to
Firmicutes. As is shown in
Figure 2A.
Proteobacteria was the most abundant phyla in the fresh
Stipa grandis. After 60 days of fermentation, the abundance of the
Proteobacteria phylum was obviously decreased, whereas the
Firmicutes phylum increased to become the new dominant phylum, which can be commonly found in previous studies [
67,
68].
Proteobacteria can result in proteolysis by competing with LAB to utilize WSC and can be animal pathogens [
69].
Firmicutes plays a pivotal role in producing LA and AA after the later stage in ensiling [
70]. This study showed a similarity in the richness and composition of bacteria in the CK, ZX301, YX 34, and P treatments at the phylum level. The relative stability of the main bacteria might reflect the existence of the core microbiome [
71].
At the genus level, the main microorganism in fresh
Stipa grandis was
Pantoea. Their portions decreased significantly and were converted to
Lactiplantibacillus plantarum after fermentation, which is in agreement with the results of You et al. [
72], who reported that
Lactobacillus can inhibit the undesirable bacteria abundance of fresh material and become the dominant genus in
Caragana silage at the later stage of ensiling.
Pantoea is harmful to fermentation by weakening the reduction in the pH value [
73].
Lactobacillus was the dominant genus in high-quality silage, which can flourish under acid environmental conditions and accumulate LA and reduce the pH value during the initiation of fermentation [
10,
11]. Therefore, the high sensitivity to the changes in pH of
Pantoea and
Lactiplantibacillus plantarum might be the main reason for the conversion of the dominant genus between fresh and silage of
Stipa grandis [
74]. In addition, compared to the CK treatment, the addition of ZX301 and YX34 increased the abundance of
Pediocossus and
Lactiplantibacillus, respectively, indicating that inoculated strains in this study were not outcompeted by the indigenous microbial population, which might be attributed to inoculants of the strains dominating and controlling the microbial events during silage fermentation [
75]. However, compared to the ZX301 treatment, the abundance of
Enterobacter increased in the YX34 treatment, which can compete for available carbohydrates with LAB and, finally, lead to a negative impact on the silage [
76], indicating that the ZX301 was more favorable for inhibiting the harmful bacteria and improving the quality of
Stipa grandis silage.
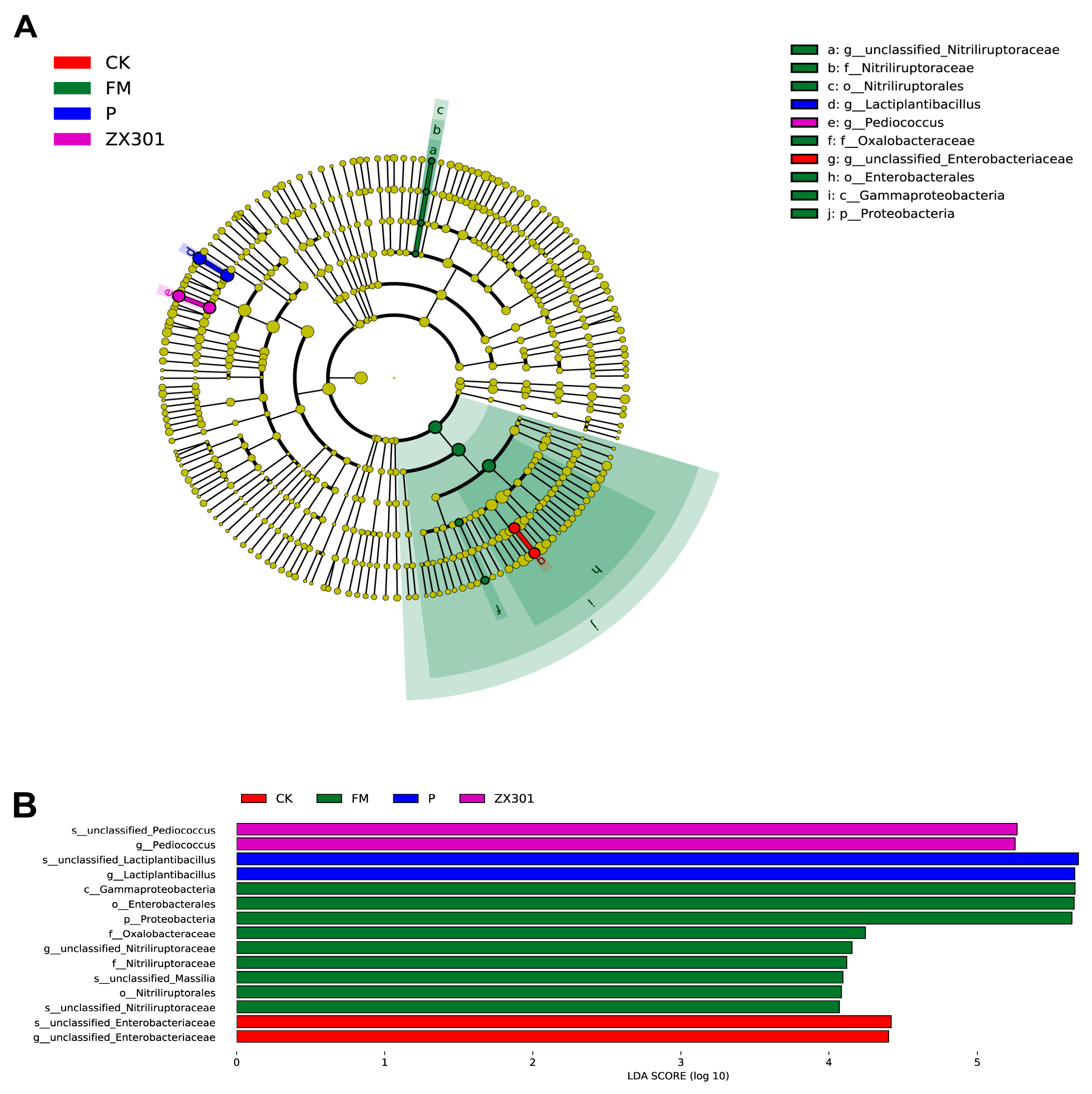
Silage fermentation is actually a process caused by microorganisms. The relationship between the chemical composition, fermentation parameters, and microbial community was elucidated using the correlation analyses in this research. In this study, a significantly higher abundance of
Lactiplantibacillus was observed in the YX34 treatment, which positively correlated with the WSC contents. The result agreed with Xu et al. [
77], who found that
Lactiplantibacillus was positively related to the WSC and NH
3-N contents. These results indicate that
Lactiplantibacillus exhibited a positive effect on nutrient preservation [
77]. In addition, the abundance of
Pediococcus was increased in the ZX301 treatment, and inversely correlated with WSC content, which suggests that this genus was primarily responsible for the LA generation and sugar consumption during ensiling in the ZX301 treatment [
66]. Interestingly, the result showed that
Enterobacter was positively correlated with pH value, and the
unclassified_Enterobacteriaceae was inversely correlated with WSC and LA content, while positively correlated with AA and NH
3-N content. These findings are corroborated by Fang et al. [
78], who reported that the genus of
Enterococcus positively correlated with AA and negatively with LA. The results of this research might be due to the acid sensitivity of
Enterococcus [
79]. Furthermore, a previous study has shown that the genera
Enterobacter and
unclassified Enterobacteriaceae belong to the
Enterobacteriaceae family, which has proteolytic activities and might compete for nutrients with LAB and form undesirable silage [
80,
81]. In the present study, the ZX301 and YX34 treatments reduced the abundance of
Enterobacter and
unclassified Enterobacteriaceae, respectively, and enabled them to become new inoculants in
Stipa grandis silage. Unfortunately,
Hafnia_Obesumbacterium genes were detected during the whole process of silage, and were positively correlated with WSC and LA content; although, their abundance was very low. The genes of
Hafnia_Obesumbacterium belong to
Enterobacteriaceae and could survive in a strict anaerobic environment and contribute to proteolysis during ensiling [
82,
83]. Taken together, the presence of
Hafnia_Obesumbacterium at least partially explains the reduction in CP content after ensiling [
84].
The 16S rRNA gene sequences of the
Stipa grandis before and after ensiled were used to infer the KEGG function profiles of microbial communities based on PICRUSt2. The result of the present study showed that the predicted functions from KEGG level 1 to level 3 were generally consistent in all treatments. The KEGG level 2 in silage fermentation mainly involves six metabolism pathways, including global and overview maps, carbohydrate metabolism, amino acid metabolism, cofactors and vitamins metabolism, energy metabolism, and nucleotide metabolism, which could be commonly found in previous studies [
84,
85]. The expression of the carbohydrate metabolism pathway was increased after ensiled, which might be explained by the process of silage fermenting the available carbohydrates to short-chain fatty acids [
66]. Similarly, Su et al. [
86] found that the abundance of carbohydrate metabolism caused an increase in fermentation progress. The amino acid metabolism was decreased more in the ZX301 treatment than that in other treatments, which might be due to the higher abundance of
Pediococcus in the ZX301 treatment, which can rapidly reduce the pH value during the early stage of silage, and then inhibit the amino acid metabolism of silage [
62,
87]. This finding was consistent with the previously published study by Bai et al. [
88], who reported that the addition of LAB can convert amino acid metabolism by modulating the bacterial communities. Additionally, the process of silage increased nucleotide metabolism in the present study. These metabolism functions are associated with the synthesis of DNA and the regulation of cellular processes [
89]. The result was in agreement with Li et al. [
90], who reported that the abundance of nucleotide metabolism increased with fermentation. However, our results could not represent the actual function of silage bacteria only based on the predicted microgenomics. Further detailed microgenomics analyses are needed to explore the potential mechanism and reveal gene functions of bacteria in silage.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}