
Molecular Detection of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays

 ,
,
Abstract
:1. Introduction
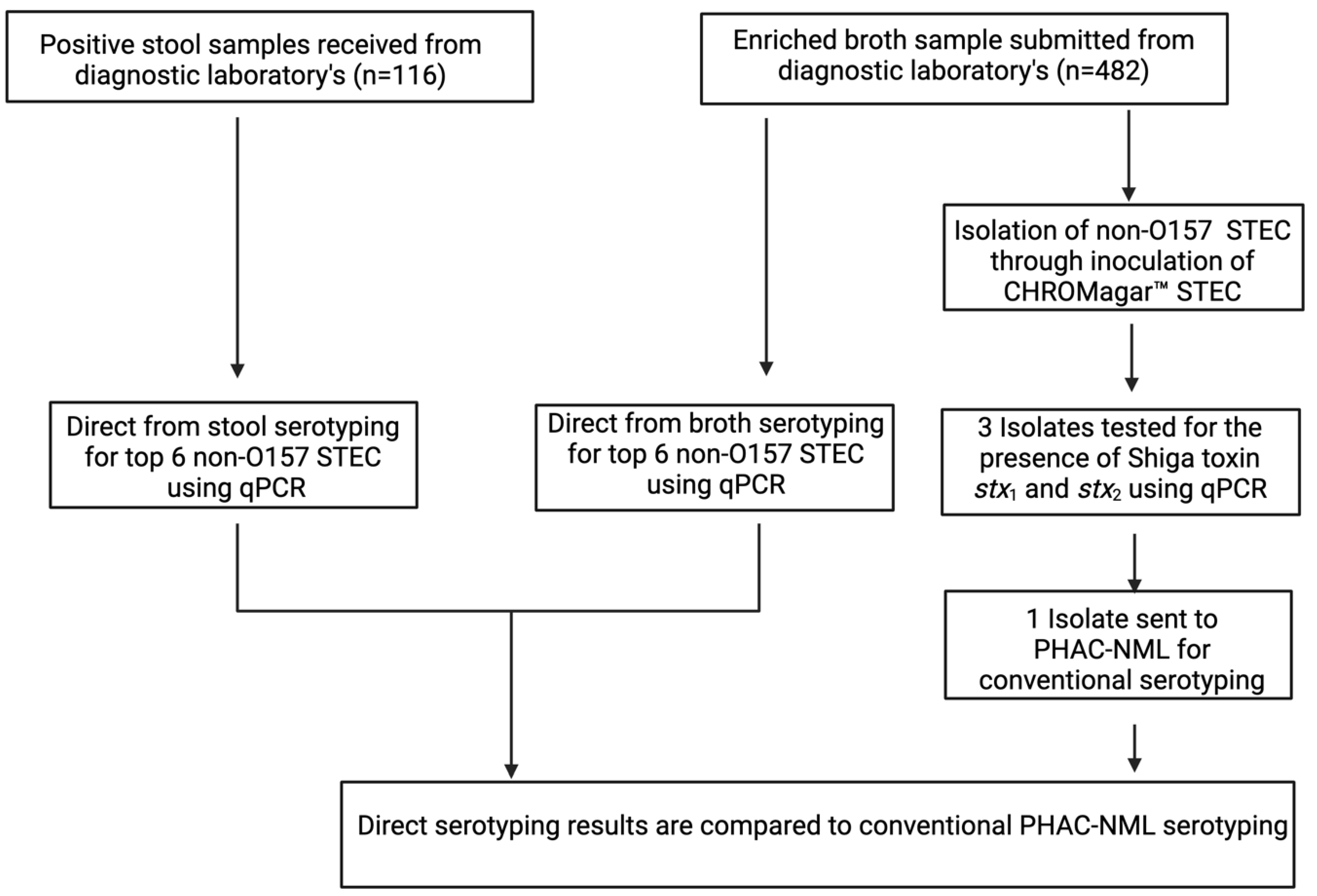
2. Materials and Methods
2.1. Sensitivity Assay
2.2. Specificity Assay
2.3. Patient Samples
2.4. Direct Molecular Serotyping from Clinical Samples
2.5. Isolation of Non-O157 STEC
2.6. Statistics
3. Results
3.1. Sensitivity and Specificity
3.2. Clinical Samples Testing
3.3. Mixed Infections between Broth and Stool Samples
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, M.K.; Murray, R.; Flockhart, L.; Pintar, K.; Fazil, A.; Nesbitt, A.; Marshall, B.; Tataryn, J.; Pollari, F. Estimates of foodborne illness-related hospitalizations and deaths in Canada for 30 specified pathogens and unspecified agents. Foodborne Pathog. Dis 2015, 12, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Bryan, A.; Youngster, I.; McAdam, A.J. Shiga Toxin Producing Escherichia coli. Clin. Lab. Med. 2015, 35, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Fuller, C.A.; Pellino, C.A.; Flagler, M.J.; Strasser, J.E.; Weiss, A.A. Shiga toxin subtypes display dramatic differences in potency. Infect. Immun. 2011, 79, 1329–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crockett, C.S.; Haas, C.N.; Fazil, A.; Rose, J.B.; Gerba, C.P. Prevalence of shigellosis in the U.S.: Consistency with dose-response information. Int. J. Food Microbiol. 1996, 30, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Menge, C. The Role of Escherichia coli Shiga Toxins in STEC Colonization of Cattle. Toxins 2020, 12, 607. [Google Scholar] [CrossRef] [PubMed]
- Hrudey, S.E.; Payment, P.; Huck, P.M.; Gillham, R.W.; Hrudey, E.J. A fatal waterborne disease epidemic in Walkerton, Ontario: Comparison with other waterborne outbreaks in the developed world. Water Sci. Technol. 2003, 47, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Karama, M.; Cenci-Goga, B.T.; Malahlela, M.; Smith, A.M.; Keddy, K.H.; El-Ashram, S.; Kabiru, L.M.; Kalake, A. Virulence Characteristics and Antimicrobial Resistance Profiles of Shiga Toxin-Producing Escherichia coli Isolates from Humans in South Africa: 2006–2013. Toxins 2019, 11, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Z.; Liang, W.; Hu, Z.; Li, X.; Guo, R.; Hua, L.; Tang, X.; Tan, C.; Chen, H.; Wang, X.; et al. O-serogroups, virulence genes, antimicrobial susceptibility, and MLST genotypes of Shiga toxin-producing Escherichia coli from swine and cattle in Central China. BMC Vet. Res. 2019, 15, 427. [Google Scholar] [CrossRef]
- De Rauw, K.; Jacobs, S.; Pierard, D. Twenty-seven years of screening for Shiga toxin-producing Escherichia coli in a university hospital. Brussels, Belgium, 1987–2014. PLoS ONE 2018, 13, e0199968. [Google Scholar] [CrossRef]
- Beutin, L.; Martin, A. Outbreak of Shiga toxin-producing Escherichia coli (STEC) O104:H4 infection in Germany causes a paradigm shift with regard to human pathogenicity of STEC strains. J. Food Prot. 2012, 75, 408–418. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Bagi, L.K.; Abdul-Wakeel, A. Detection and Isolation of the "Top Seven" Shiga Toxin-Producing Escherichia coli in Ground Beef: Comparison of RapidFinder Kits to the U.S. Department of Agriculture Microbiology Laboratory Guidebook Method. J. Food Prot. 2017, 80, 829–836. [Google Scholar] [CrossRef]
- Zelyas, N.; Poon, A.; Patterson-Fortin, L.; Johnson, R.P.; Lee, W.; Chui, L. Assessment of commercial chromogenic solid media for the detection of non-O157 Shiga toxin-producing Escherichia coli (STEC). Diagn. Microbiol. Infect. Dis. 2016, 85, 302–308. [Google Scholar] [CrossRef] [PubMed]
- Bording-Jorgensen, M.; Tyrrell, H.; Lloyd, C.; Chui, L. Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli. Microorganisms 2020, 9, 503. [Google Scholar] [CrossRef]
- Chui, L.; Christianson, S.; Alexander, D.C.; Arseneau, V.; Bekal, S.; Berenger, B.; Chen, Y.; Davidson, R.; Farrell, D.J.; German, G.J.; et al. CPHLN recommendations for the laboratory detection of Shiga toxin-producing Escherichia coli (O157 and non-O157). Can. Commun. Dis. Rep. 2018, 44, 304–307. [Google Scholar] [CrossRef]
- Marder, E.P.; Cieslak, P.R.; Cronquist, A.B.; Dunn, J.; Lathrop, S.; Rabatsky-Ehr, T.; Ryan, P.; Smith, K.; Tobin-D’Angelo, M.; Vugia, D.J.; et al. Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food and the Effect of Increasing Use of Culture-Independent Diagnostic Tests on Surveillance-Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2013–2016. MMWR. Morb. Mortal. Wkly. Rep. 2017, 66, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Jaros, P.; Dufour, M.; Gilpin, B.; Freeman, M.M.; Ribot, E.M. PFGE for Shiga toxin-producing Escherichia coli O157:H7 (STEC O157) and non-O157 STEC. Methods Mol. Biol. 2015, 1301, 171–189. [Google Scholar] [CrossRef]
- Bogaerts, B.; Nouws, S.; Verhaegen, B.; Denayer, S.; Van Braekel, J.; Winand, R.; Fu, Q.; Crombe, F.; Pierard, D.; Marchal, K.; et al. Validation strategy of a bioinformatics whole genome sequencing workflow for Shiga toxin-producing Escherichia coli using a reference collection extensively characterized with conventional methods. Microb. Genom. 2021, 7, 531. [Google Scholar] [CrossRef]
- Holland, J.L.; Louie, L.; Simor, A.E.; Louie, M. PCR detection of Escherichia coli O157:H7 directly from stools: Evaluation of commercial extraction methods for purifying fecal DNA. J. Clin. Microbiol. 2000, 38, 4108–4113. [Google Scholar] [CrossRef] [Green Version]
- Perelle, S.; Dilasser, F.; Grout, J.; Fach, P. Detection by 5’-nuclease PCR of Shiga-toxin producing Escherichia coli O26, O55, O91, O103, O111, O113, O145 and O157:H7, associated with the world’s most frequent clinical cases. Mol. Cell. Probes 2004, 18, 185–192. [Google Scholar] [CrossRef]
- Reynolds, C.; Checkley, S.; Chui, L.; Otto, S.; Neumann, N.F. Evaluating the risks associated with Shiga-toxin-producing Escherichia coli (STEC) in private well waters in Canada. Can. J. Microbiol. 2020, 66, 337–350. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, 324. [CrossRef]
- Gill, A.; Carrillo, C.; Hadley, M.; Kenwell, R.; Chui, L. Bacteriological analysis of wheat flour associated with an outbreak of Shiga toxin-producing Escherichia coli O121. Food Microbiol. 2019, 82, 474–481. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Reference Gene, Primer/Probe | Sequence 5′–3′ [19] |
|---|---|
| stx1-F | TTT GTY ACT GTS ACA GCW GAA GCY TTA CG |
| stx1-R | CCC CAG TTC ARW GTR AGR TCM ACR TC |
| stx1-P | CTG GAT GAT CTC AGT GGG CGT TCT TAT GTA A |
| stx2-F | TTT GTY ACT GTS ACA GCW GAA GCY TTA CG |
| stx2-R | CCC CAG TTC ARW GTR AGR TCM ACR TC |
| stx2-P | TCG TCA GGC ACT GTC TGA AAC TGC TCC |
| wzx O26-F | CGC GAC GGC AGA GAA AAT T |
| wzx O26-R | AGC AGG CTT TTA TAT TCT CCA ACT TT |
| wzx O26-P | CCC CGT TAA ATC AAT ACT ATT TCA CGA GGT TGA |
| wzx O45-F | TAC GTC TGG CTG CAG GG |
| wzx O45-R | ACT TGC AGC AAA AAA TCC CC |
| wzx O45-P | TTC GTT GCG TTG TGC ATG GTG GC |
| wzx O103-F | CAA GGT GAT TAC GAA AAT GCA TGT |
| wzx O103-R | GAA AAA AGC ACC CCC GTA CTT AT |
| wzx O103-P | CAT AGC CTG TTG TTT TAT |
| wbdl O111-F | CGA GGC AAC ACA TTA TAT AGT GCT TT |
| wbdl O111-R | TTT TTG AAT AGT TAT GAA CAT CTT GTT TAG C |
| wbdl O111-P | TTG AAT CTC CCA GAT GAT CAA CAT CGT GAA |
| wzx O121-F | TGG TCT CTT AGA CTT AGG GC |
| wzx O121-R | TTA GCA ATT TTC TGT AGT CCA GC |
| wzx O121-P | TCC AAC AAT TGG TCG TGA AAC AGC TCG |
| ihpl O145-F | CGA TAA TAT TTA CCC CAC CAG TAC AG |
| ihpl O145-R | CCG CCA TTC AGA ATG CAC ACA ATA TCG |
| ihpl O145-P | ACA GTG CCA GCA TTC GCT TGC GA |
| Singleplex (Ct Values) | ||||||
| Primer/Probe Set | O26 | O111 | O45 | O121 | O103 | O145 |
| Target Organism | E. coli O26 | E. coli O111 | E. coli O45 | E. coli O121 | E. coli O103 | E. coli O145 |
| 104 (CFU/mL) | 25.6 | 27.5 | 26.3 | 25.2 | 29.9 | 25.5 |
| 103 (CFU/mL) | 29.3 | 30.8 | 30.6 | 28.8 | 33.5 | 28.7 |
| 102 (CFU/mL) | 35 | 34.3 | 33 | 33.8 | 35 | 31.8 |
| Multiplex (Ct Values) | ||||||
| Primer/Probe Set | O26/O45 | O26/O45 | O121/O145 | O121/O145 | O103/O111 | O111/O103 |
| Target Organism | E. coli O26 | E. coli O45 | E. coli O145 | E. coli O121 | E. coli O103 | E. coli O111 |
| 104 (CFU/mL) | 25.6 | 26.3 | 25.4 | 25.3 | 29.4 | 27.4 |
| 103 (CFU/mL) | 29.4 | 30.5 | 28.9 | 28.9 | 33.4 | 31.3 |
| 102 (CFU/mL) | 35 | 32 | 31.5 | 33 | 35 | 34.6 |
| Serotype | Stools/Broths/Isolate | Stools/Broths | Stools/Isolate | Broths/Isolate | Broth-Only | Isolate-Only | Stool-Only |
|---|---|---|---|---|---|---|---|
| E. coli O26 | 37 | 1 | 0 | 1 | 0 | 0 | 2 # |
| E. coli O111 | 17 | 0 | 0 | 0 | 0 | 0 | 1 # |
| E. coli O121 | 7 | 0 | 0 | 1 | 0 | 0 | 3 |
| E. coli O103 | 10 | 0 | 0 | 8 | 0 | 0 | 1 |
| E. coli O145 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| E. coli non-top six | 25 | 0 | 0 | 3 | 0 | 1 | 7 |
| Serotype | Broths/Isolate | Broth-Only | Isolate Only |
|---|---|---|---|
| E. coli O26 | 106 | 1 | 0 |
| E. coli O111 | 63 | 1 | 1 |
| E. coli O121 | 25 | 0 | 1 |
| E. coli O103 | 50 | 4 | 0 |
| E. coli O145 | 11 | 1 | 0 |
| E. coli non-top six | 102 | 2 | 7 |
| Serotype | stx1 | stx2 | stx1 and stx2 |
|---|---|---|---|
| E. coli O26 | 117 | 13 | 16 |
| E. coli O111 | 64 | 10 | 7 |
| E. coli O121 | 31 | 4 | 0 |
| E. coli O103 | 64 | 2 | 4 |
| E. coli O145 | 12 | 4 | 2 |
| E. coli non-top six | 105 | 26 | 7 |
| In-House Direct Serotype | Conventional Serotype Based on Picked Colonies from CHROMagar™ STEC Plate |
|---|---|
| O26 | O69 |
| O145 | O27 |
| O111 | O: undetermined |
| O26 | O: undetermined |
| O103 | O187 |
| O103 | O69 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bording-Jorgensen, M.; Parsons, B.; Szelewicki, J.; Lloyd, C.; Chui, L. Molecular Detection of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays. Microorganisms 2022, 10, 329. https://doi.org/10.3390/microorganisms10020329
Bording-Jorgensen M, Parsons B, Szelewicki J, Lloyd C, Chui L. Molecular Detection of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays. Microorganisms. 2022; 10(2):329. https://doi.org/10.3390/microorganisms10020329
Chicago/Turabian StyleBording-Jorgensen, Michael, Brendon Parsons, Jonas Szelewicki, Colin Lloyd, and Linda Chui. 2022. "Molecular Detection of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays" Microorganisms 10, no. 2: 329. https://doi.org/10.3390/microorganisms10020329
APA StyleBording-Jorgensen, M., Parsons, B., Szelewicki, J., Lloyd, C., & Chui, L. (2022). Molecular Detection of Non-O157 Shiga Toxin-Producing Escherichia coli (STEC) Directly from Stool Using Multiplex qPCR Assays. Microorganisms, 10(2), 329. https://doi.org/10.3390/microorganisms10020329