
Exploration of Bacterial Re-Growth as In Vitro Phenomenon Affecting Methods for Analysis of the Antimicrobial Activity of Chimeric Bacteriophage Endolysins

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Determination
2.2. Time–Kill Assays
- (i)
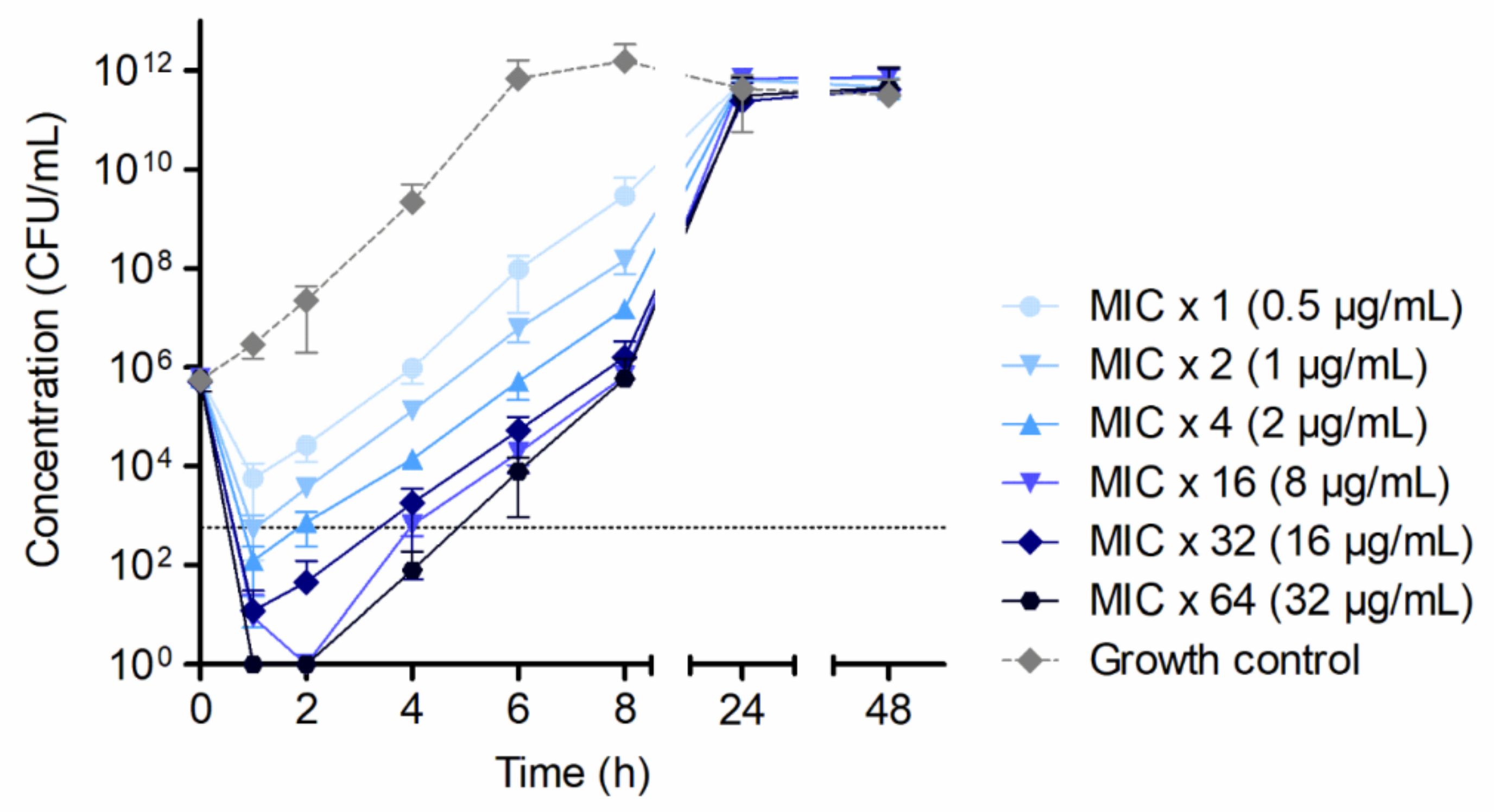
- Evaluation of bactericidal concentrations of HY-133 against S. aureus ATCC 29213 over time was based on MIC values determined via broth microdilution methodology. Time–kill curves were carried out in glass baffled flasks (GBF; Schott AG, Mainz, Germany). The 1-, 2-, 4-, 16-, 32-, and 64-fold MIC of HY-133 (HYPharm GmbH, Bernried, Germany) was analyzed.
- (ii)
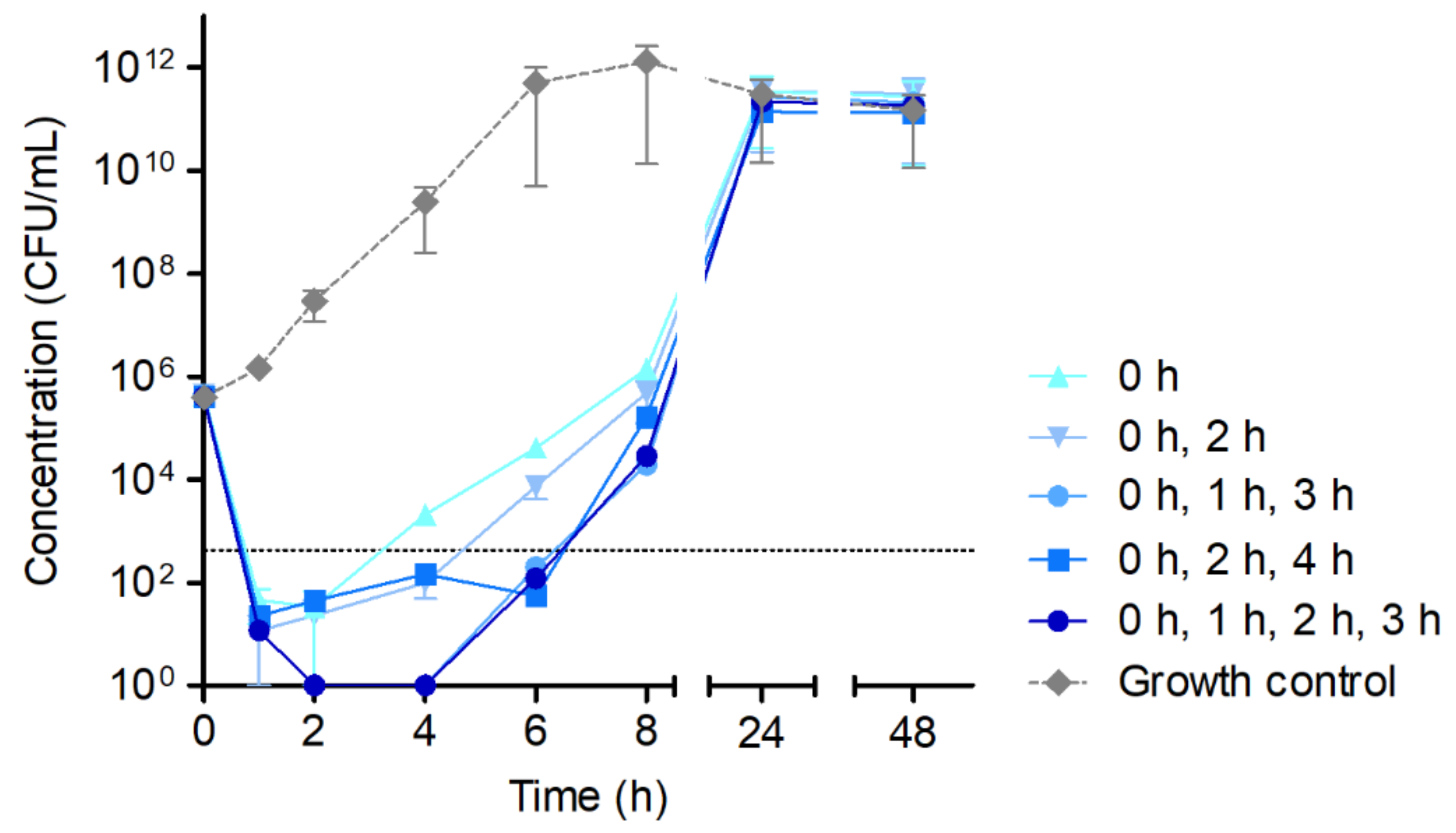
- Interval dosage experiments were carried out adding 16-fold MIC of HY-133 to the ATCC 29213 culture at time point 0 h and after (i) 2 h, (ii) 1 h and 3 h, (iii) 2 h and 4 h, or (iv) 1 h, 2 h, and 3 h, respectively. Experiments were performed in GBFs (Schott AG).
- (iii)
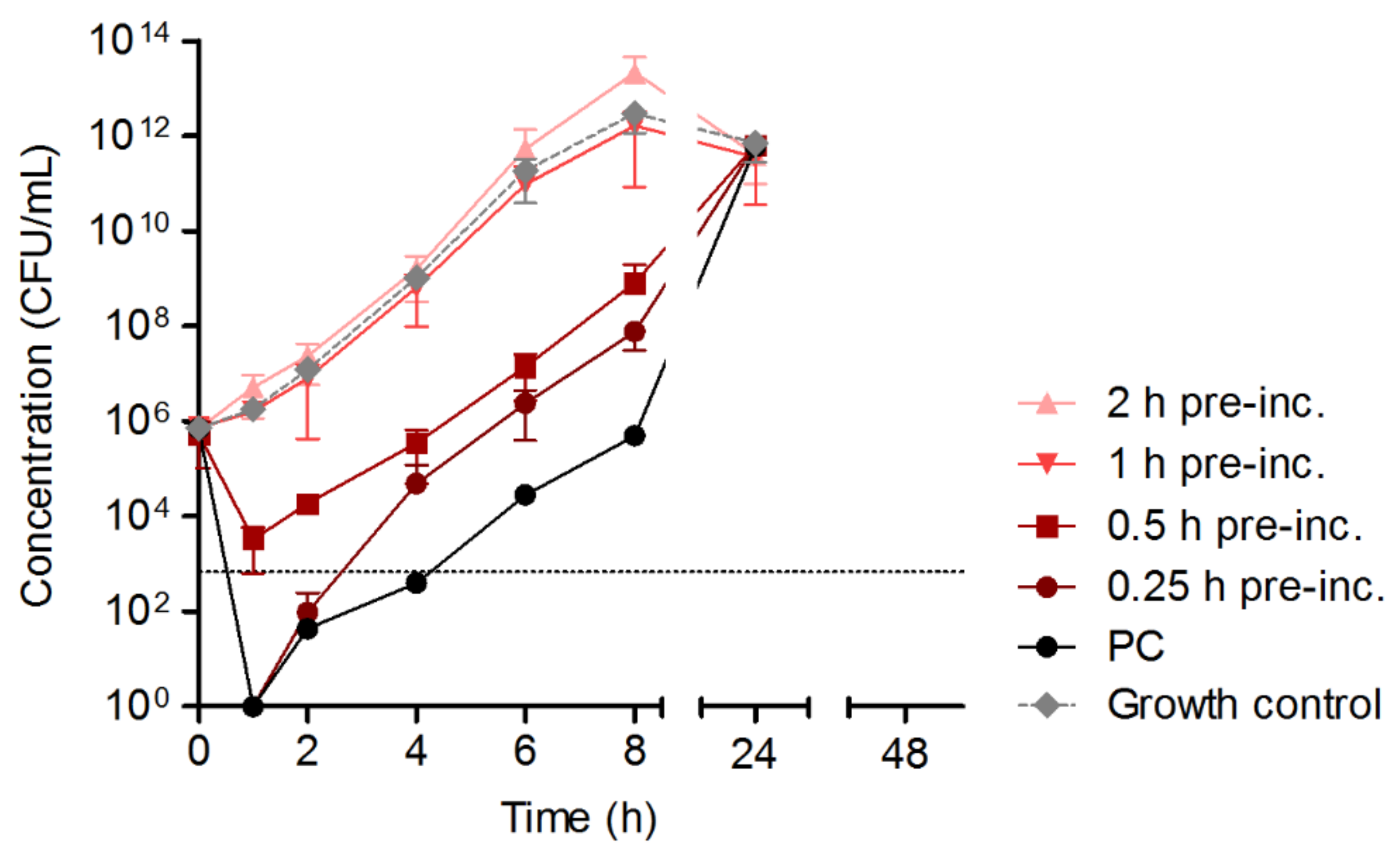
- The maintenance of HY-133 activity against S. aureus ATCC 29213 in time–kill curve experiments (CLSI guideline M26-A) [19] was determined by pre-incubation of the substance (16-fold MIC dissolved in CAMHB (Becton Dickinson)) for different time spans (0.25 h, 0.5 h, 1 h, and 2 h) under given conditions (GBFs, 37 °C, 160 rpm) before adding the starting inoculum of S. aureus ATCC 29213. GBFs (Schott AG) were further used for time–kill curves.
- (iv)
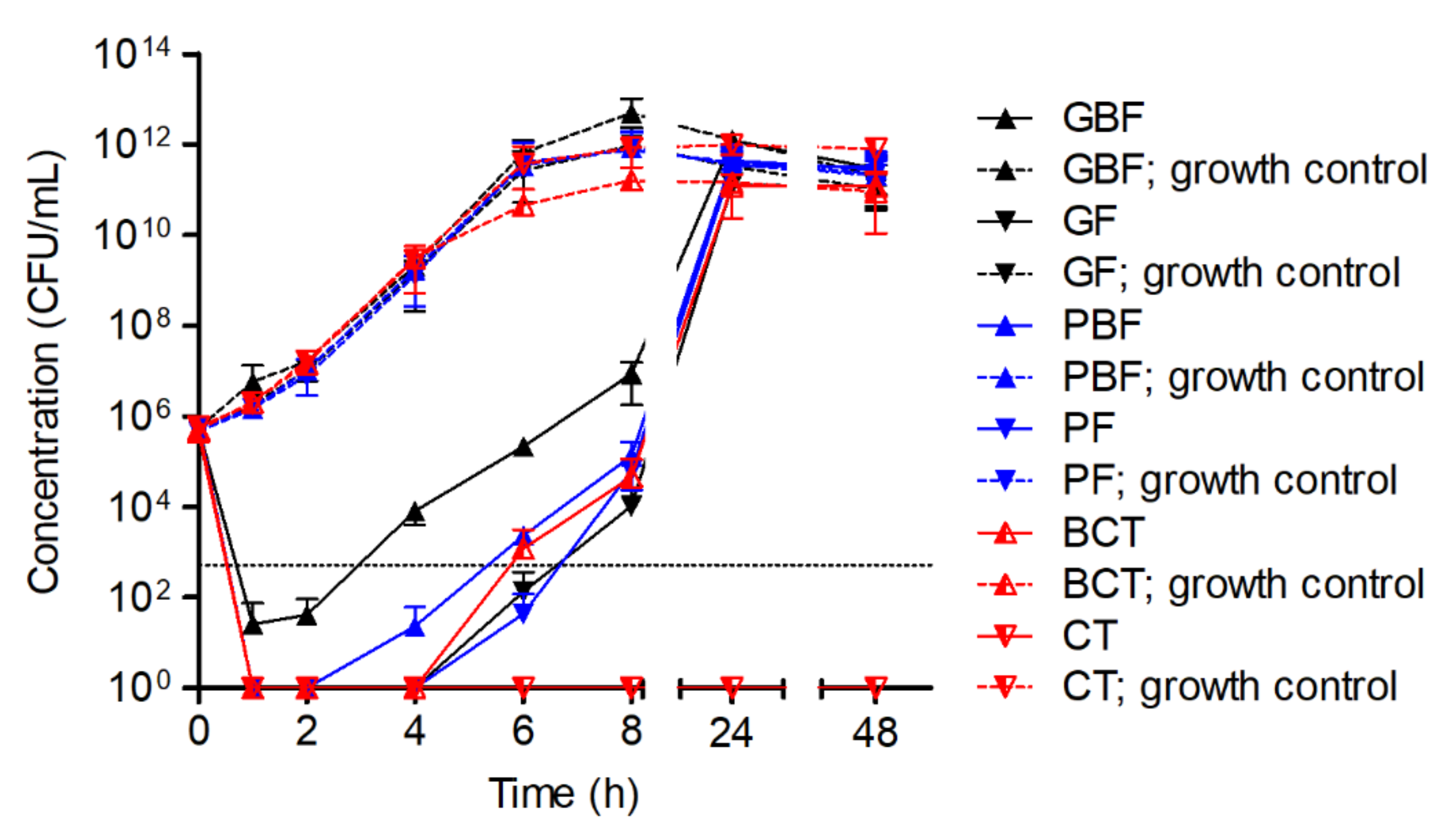
- The influence of different shearing forces, culture flask materials, and their forms was evaluated by applying the 16-fold MIC of HY-133 against S. aureus strain ATCC 29213 in different culture vessels (Table 1). Incubation was carried out shaking at 160 rpm.
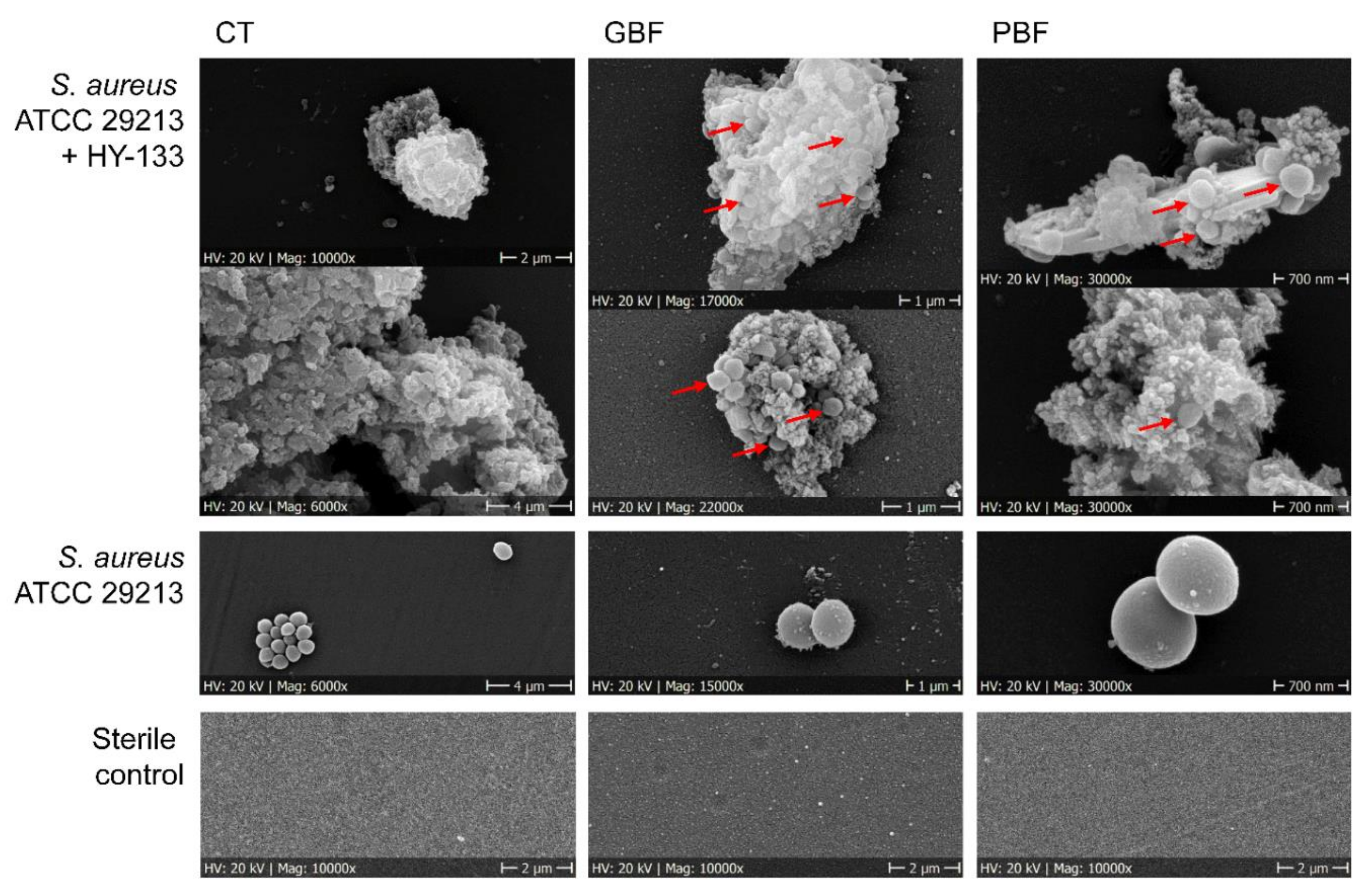
2.3. Scanning Electron Microscopy
2.4. Fluorescence Assays
2.5. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE)
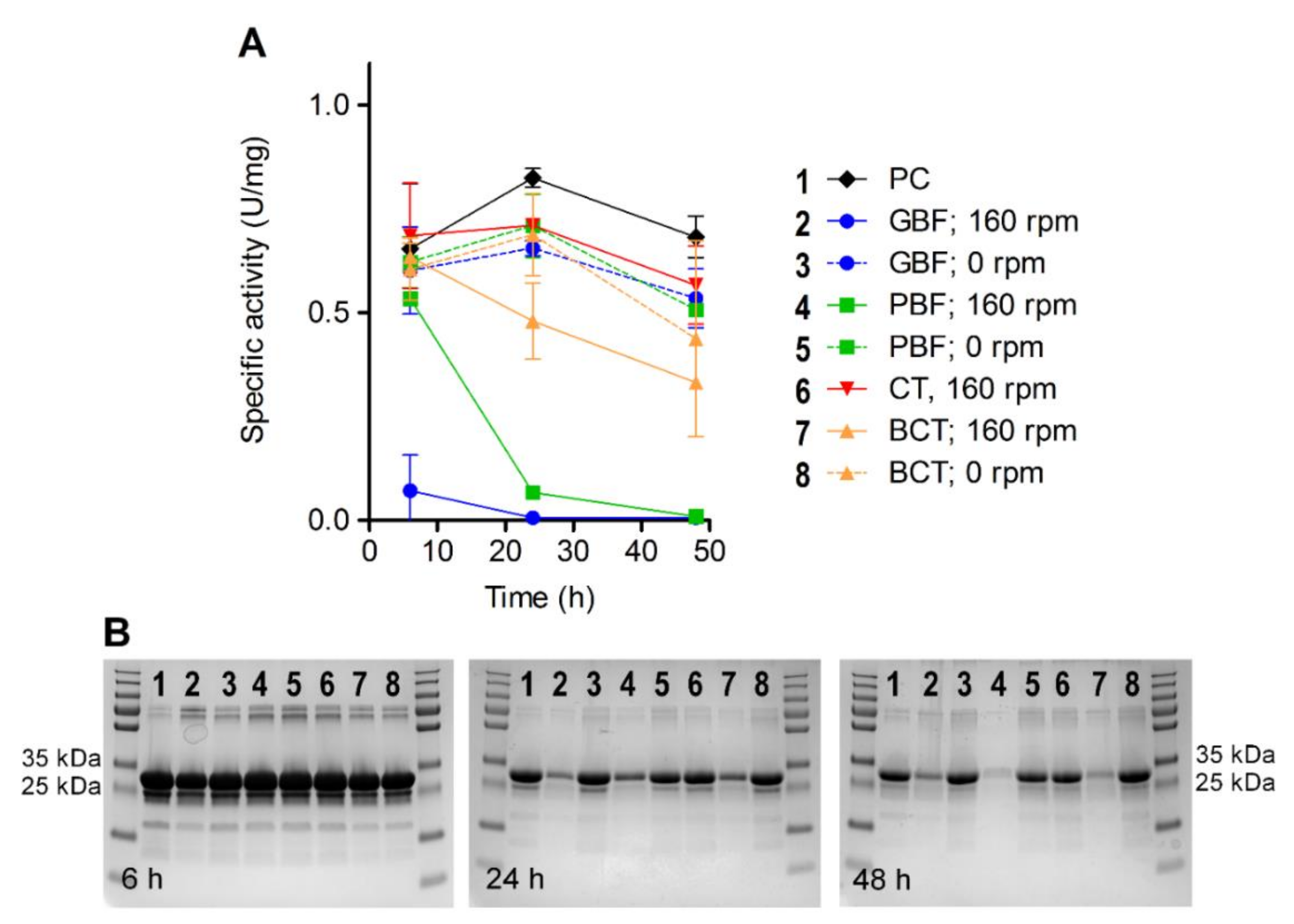
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, A.S.; De Lencastre, H.; Garau, J.; Kluytmans, J.; Malhotra-Kumar, S.; Peschel, A.; Harbarth, S. Methicillin-resistant Staphylococcus aureus. Nat. Rev. Dis. Primers 2018, 4, 18033. [Google Scholar] [CrossRef] [PubMed]
- Köck, R.; Becker, K.; Cookson, B.; van Gemert-Pijnen, J.E.; Harbarth, S.; Kluytmans, J.; Mielke, M.; Peters, G.; Skov, R.L.; Struelens, M.J.; et al. Systematic literature analysis and review of targeted preventive measures to limit healthcare-associated infections by meticillin-resistant Staphylococcus aureus. Eurosurveillance 2014, 19, 20860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, H.; Becker, K.; Dohmen, P.M.; Petrosillo, N.; Spencer, M.; Van Rijen, M.; Wechsler-Fördös, A.; Pujol, M.; Dubouix, A.; Garau, J. Staphylococcus aureus and surgical site infections: Benefits of screening and decolonization before surgery. J. Hosp. Infect. 2016, 94, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Saraswat, M.K.; Magruder, J.T.; Crawford, T.C.; Gardner, J.M.; Duquaine, D.; Sussman, M.S.; Maragakis, L.L.; Whitman, G.J. Preoperative Staphylococcus aureus screening and targeted decolonization in cardiac surgery. Ann. Thorac. Surg. 2017, 104, 1349–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurke, A.; Daniels-Haardt, I.; Silvis, W.; Berends, M.S.; Glasner, C.; Becker, K.; Köck, R.; Friedrich, A.W. Changing epidemiology of meticillin-resistant Staphylococcus aureus in 42 hospitals in the Dutch-German border region, 2012 to 2016: Results of the search-and-follow-policy. Eurosurveillance 2019, 24, 1800244. [Google Scholar] [CrossRef]
- Poovelikunnel, T.; Gethin, G.; Humphreys, H. Mupirocin resistance: Clinical implications and potential alternatives for the eradication of MRSA. J. Antimicrob. Chemother. 2015, 70, 2681–2692. [Google Scholar] [CrossRef] [Green Version]
- McNeil, J.C.; Kok, E.Y.; Vallejo, J.G.; Campbell, J.R.; Hulten, K.G.; Mason, E.O.; Kaplan, S.L. Clinical and Molecular Features of Decreased Chlorhexidine Susceptibility among Nosocomial Staphylococcus aureus Isolates at Texas Children’s Hospital. Antimicrob. Agents Chemother. 2016, 60, 1121–1128. [Google Scholar] [CrossRef] [Green Version]
- Dadashi, M.; Hajikhani, B.; Darban-Sarokhalil, D.; van Belkum, A.; Goudarzi, M. Mupirocin resistance in Staphylococcus aureus: A systematic review and meta-analysis. J. Glob. Antimicrob. Resist. 2020, 20, 238–247. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, H.; Han, L.; Shu, W.; Wu, Q.; Ni, Y. Frequency of biocide-resistant genes and susceptibility to chlorhexidine in high-level mupirocin-resistant, methicillin-resistant Staphylococcus aureus (MuH MRSA). Diagn. Microbiol. Infect. Dis. 2015, 82, 278–283. [Google Scholar] [CrossRef]
- Fischetti, V.A. Development of Phage Lysins as Novel Therapeutics: A Historical Perspective. Viruses 2018, 10, 310. [Google Scholar] [CrossRef] [Green Version]
- Idelevich, E.A.; von Eiff, C.; Friedrich, A.W.; Iannelli, D.; Xia, G.; Peters, G.; Peschel, A.; Wanninger, I.; Becker, K. In vitro activity against Staphylococcus aureus of a novel antimicrobial agent, PRF-119, a recombinant chimeric bacteriophage endolysin. Antimicrob. Agents Chemother. 2011, 55, 4416–4419. [Google Scholar] [CrossRef] [Green Version]
- Schleimer, N.; Kaspar, U.; Knaack, D.; von Eiff, C.; Molinaro, S.; Grallert, H.; Idelevich, E.A.; Becker, K. In vitro activity of the bacteriophage endolysin HY-133 against Staphylococcus aureus small-colony variants and their corresponding wild types. Int. J. Mol. Sci. 2019, 20, 716. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, U.; de Haro Sautto, J.A.; Molinaro, S.; Peters, G.; Idelevich, E.A.; Becker, K. The novel phage-derived antimicrobial agent HY-133 Is active against livestock-associated methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2018, 62, e00385-18. [Google Scholar] [CrossRef] [Green Version]
- Idelevich, E.A.; Schaumburg, F.; Knaack, D.; Scherzinger, A.S.; Mutter, W.; Peters, G.; Peschel, A.; Becker, K. The recombinant bacteriophage endolysin HY-133 exhibits in vitro activity against different African clonal lineages of the Staphylococcus aureus complex, including Staphylococcus schweitzeri. Antimicrob. Agents Chemother. 2016, 60, 2551–2553. [Google Scholar] [CrossRef] [Green Version]
- Knaack, D.; Idelevich, E.A.; Schleimer, N.; Molinaro, S.; Kriegeskorte, A.; Peters, G.; Becker, K. Bactericidal activity of bacteriophage endolysin HY-133 against Staphylococcus aureus in comparison to other antibiotics as determined by minimum bactericidal concentrations and time-kill analysis. Diagn. Microbiol. Infect. Dis. 2019, 93, 362–368. [Google Scholar] [CrossRef]
- Idelevich, E.A.; Knaack, D.; Nugroho, N.T.; Peters, G.; Bisdas, T.; Molinaro, S.; Torsello, G.B.; Becker, K.; Herten, M. Comparative in vitro activity of bacteriophage endolysin HY-133 against Staphylococcus aureus attached to vascular graft surface. Med. Microbiol. Immunol. 2020, 209, 51–57. [Google Scholar] [CrossRef]
- M100-S29; Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2019.
- M07-A11; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical Laboratory Standards Institute: Wayne, PA, USA, 2018.
- M26-A; Methods for Determination of Bactericidal Activity of Antimicrobial Agents; CLSI document; National Committee on Clinical Laboratory Standards: Wayne, PA, USA, 1999.
- Herten, M.; Bisdas, T.; Knaack, D.; Becker, K.; Osada, N.; Torsello, G.B.; Idelevich, E.A. Rapid in vitro quantification of S. aureus biofilms on vascular graft surfaces. Front. Microbiol. 2017, 8, 2333. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, H.M.; Silva, R.A.; Cesari, A.C.; Zamit, A.L.; Peressutti, S.R.; Reichelt, R.; Keller, U.; Malkus, U.; Rasch, C.; Maskow, T.; et al. Physiological and morphological responses of the soil bacterium Rhodococcus opacus strain PD630 to water stress. FEMS Microbiol. Ecol. 2004, 50, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Mulisch, M.; Welsch, U. Romeis Mikroskopische Technik, 19th ed.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Vollmer, W.; Blanot, D.; de Pedro, M.A. Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 2008, 32, 149–167. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Dedent, A.C.; Schneewind, O. Sortases and the art of anchoring proteins to the envelopes of gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 192–221. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischetti, V.A. Bacteriophage lytic enzymes: Novel anti-infectives. Trends Microbiol. 2005, 13, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Fischetti, V.A. Bacteriophage lysins as effective antibacterials. Curr. Opin. Microbiol. 2008, 11, 393–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handwerger, S.; Tomasz, A. Antibiotic tolerance among clinical isolates of bacteria. Rev. Infect. Dis. 1985, 7, 368–386. [Google Scholar] [CrossRef]
- Kumar, J.K. Lysostaphin: An antistaphylococcal agent. Appl. Microbiol. Biotechnol. 2008, 80, 555–561. [Google Scholar] [CrossRef]
- Schindler, C.A.; Schuhardt, V.T. Lysostaphin: A New Bacteriolytic Agent for the Staphylococcus. Proc. Natl. Acad. Sci. USA 1964, 51, 414–421. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, S.; Ross, R.P.; Meaney, W.; Fitzgerald, G.F.; Elbreki, M.F.; Coffey, A. Potential of the polyvalent anti-Staphylococcus bacteriophage K for control of antibiotic-resistant staphylococci from hospitals. Appl. Environ. Microbiol. 2005, 71, 1836–1842. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Vessel | Material | Volume (mL) | Manufacturer |
|---|---|---|---|
| Glass baffled flask (GBF) | Glass | 100 | Schott AG, Mainz, Germany |
| Glass flask (GF) | Glass | 100 | Schott AG, Mainz, Germany |
| Plastic baffled flask (PBF) | Non-pyrogenic polycarbonate | 125 | Corning Incorporated, Corning, NY, USA |
| Plastic flask (PF) | Non-pyrogenic polycarbonate | 125 | Corning Incorporated, Corning, NY, USA |
| Baffled centrifuge tube (BCT) 1 | Polypropylene | 50 | Greiner Bio One International, Kremsmünster, Austria |
| Centrifuge tube (CT) | Polypropylene | 50 | Greiner Bio One International, Kremsmünster, Austria |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaspar, U.; Schleimer, N.; Idelevich, E.A.; Molinaro, S.; Becker, K. Exploration of Bacterial Re-Growth as In Vitro Phenomenon Affecting Methods for Analysis of the Antimicrobial Activity of Chimeric Bacteriophage Endolysins. Microorganisms 2022, 10, 445. https://doi.org/10.3390/microorganisms10020445
Kaspar U, Schleimer N, Idelevich EA, Molinaro S, Becker K. Exploration of Bacterial Re-Growth as In Vitro Phenomenon Affecting Methods for Analysis of the Antimicrobial Activity of Chimeric Bacteriophage Endolysins. Microorganisms. 2022; 10(2):445. https://doi.org/10.3390/microorganisms10020445
Chicago/Turabian StyleKaspar, Ursula, Nina Schleimer, Evgeny A. Idelevich, Sonja Molinaro, and Karsten Becker. 2022. "Exploration of Bacterial Re-Growth as In Vitro Phenomenon Affecting Methods for Analysis of the Antimicrobial Activity of Chimeric Bacteriophage Endolysins" Microorganisms 10, no. 2: 445. https://doi.org/10.3390/microorganisms10020445
APA StyleKaspar, U., Schleimer, N., Idelevich, E. A., Molinaro, S., & Becker, K. (2022). Exploration of Bacterial Re-Growth as In Vitro Phenomenon Affecting Methods for Analysis of the Antimicrobial Activity of Chimeric Bacteriophage Endolysins. Microorganisms, 10(2), 445. https://doi.org/10.3390/microorganisms10020445