Abstract
We investigated the combined effects of biopreservation and high-pressure treatment on bacterial communities of diced cooked ham prepared with diminished nitrite salt. First, bacterial communities of four commercial brands of diced cooked ham from local supermarkets were characterized and stored frozen. Second, sterile diced cooked ham, prepared with reduced levels of nitrite, was inoculated with two different microbiota collected from the aforementioned commercial samples together with a nisin-producing Lactococcus lactis protective strain able to recover from a 500 MPa high-pressure treatment. Samples were then treated at 500 MPa for 5 min, and bacterial dynamics were monitored during storage at 8 °C. Depending on samples, the ham microbiota was dominated by different Proteobacteria (Pseudomonas, Serratia, Psychrobacter, or Vibrio) or by Firmicutes (Latilactobacillus and Leuconostoc). Applied alone, none of the treatments stabilized during the growth of the ham microbiota. Nevertheless, the combination of biopreservation and high-pressure treatment was efficient in reducing the growth of Proteobacteria spoilage species. However, this effect was dependent on the nature of the initial microbiota, showing that the use of biopreservation and high-pressure treatment, as an alternative to nitrite reduction for ensuring cooked ham microbial safety, merits attention but still requires improvement.
1. Introduction
Nitrite salts have been used since ancient times as curing agents for the production of cured meat products. Nitrite (and eventually nitrate) salts are commonly added to the brine for the manufacturing of cooked ham. Their role is important for the typical pinky/reddish color development of cured meats [1]. Nitrite salts also participate in the hurdle technology for ensuring microbial safety as bactericidal and bacteriostatic agents against several pathogenic bacteria occurring in meat products, in particular Clostridium botulinum [2]. However, nitrite can potentially lead to carcinogenic nitrosamine production in these products, which raises the concern of a potential public health risk [3]. In the European Union (EU), all additives authorized before the 20 January 2009 had to be re-evaluated by 2020. In that context, the European Food Safety Authority (EFSA) re-assessed the safety of nitrite and nitrate salts in 2017. The current acceptable daily intakes for nitrite and nitrate were assessed, and experts stated that exposure to nitrites used as food additives was within safe levels for most of the population, except for children [4]. Nevertheless, although experts evaluated the need for further scientific information, a positive link could be established between the dietary intake of nitrite (or of processed meat containing both nitrite and nitrate) and some cancers [5,6]. Therefore, reducing nitrite levels in processed meats appears necessary in order to limit this sanitary issue.
Several hurdle technologies can be proposed to compensate for a higher risk of microbial safety in cooked ham with reduced nitrite salts. Among those, high-pressure processing (HPP) has been proposed in the context of food additive reduction (see [7] for a review) and was shown to efficiently reduce bacterial contaminants in cooked ham after applying 400–600 MPa treatment for 3–20 min depending on the study [8,9,10,11,12,13,14,15,16,17,18], with many reporting an effect at 400 or 500 MPa [9,10,11,12,14,15,16].
Biopreservation, a method using protective cultures, often lactic acid bacteria (LAB) or their metabolites, was described more than 20 years ago for fighting undesired bacterial contaminants [19]. The addition of such LAB protective cultures to cooked meat products, and in particular, cooked ham was indeed reported [20,21]. HPP combined with biopreservation has also been studied [22]. HPP treatment of cooked ham treated with bacteriocins has been shown to reduce the population of several bacterial pathogens or to prolong cold storage [12,13,16,23,24].
In the present study, we investigated the combined effect of HPP and biopreservation on the dynamics of bacterial communities of cooked ham with a reduced level of nitrite salts. We had previously selected a nisin-producing Lactococcus lactis strain, which, although sensitive to HPP treatment, was able to recover and regrow after a 500 MPa HPP treatment [25]. Here, we first determined and collected bacterial communities present on commercial diced cooked ham. Then, the dynamics of these bacterial communities were monitored, following their inoculation in sterile diced cooked ham with a reduced nitrite level, in the presence of the bioprotective strain, and after the application of the HPP treatment. Our study provides data in line with the following questions: does HPP display a generic effect on microbial inactivation, or is there a microbiota signature in pressurized ham? Is the use of nisin-producing L. lactis mediated biopreservation a worthwhile hurdle strategy when combined with HPP, and is this biopreservation effect also microbiota dependent?
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
Lactococcus lactis CH-HP15 [25] was first cultivated on M17 agar plates (Biokar diagnostic, Beauvais, France) at 30 °C for 72 h. One colony was inoculated for preculture in Medium Modelling Ham (MMH) [26] at 30 °C for 24 h, and then cultured in MMH at 30 °C under agitation at 65 rpm for 13 h until the early stationary phase. Bacterial enumerations were performed by plating serial dilutions of bacterial cultures, microbiota, or ham stomached samples. The total viable counts were determined on Plate Count Agar (PCA) (Biokar diagnostic, Beauvais, France). Psychrophilic and mesophilic counts were determined after incubation for 4 days at 15 °C or 24 h at 30 °C, respectively. LAB were enumerated after 4 days incubation at 25 °C on de Man Rogosa and Sharpe (MRS) agar medium, pH 5.2 (AES, Rennes, France), containing bromocresol green (25 mg·L−1) to estimate LAB diversity, as previously described [27]. In ham samples inoculated with L. lactis CH-HP15, M17 agar plates were used for enumeration after 48 h incubation at 30 °C. For anaerobic conditions, plates were incubated in jars with anaerocult sachets (Anaerocult A, Merck, Darmstatd, Germany).
2.2. Ham Sampling and Microbiota Recovery
Diced cooked ham (except for one sample consisting of sliced cooked ham) was purchased in 2015 and 2016 from local supermarkets in Nantes, France. They were purchased as early as possible considering the use-by-date, transported at cold temperature to the laboratory, and reconditioned immediately. These samples were sold as ready-to-eat packs of 120 to 200 g, conditioned under a modified atmosphere without any indication of the gas mixture used in the packs. At arrival in the laboratory (day 1), bags were opened, and samples were immediately reconditioned as small (25 g) aliquots that were further used for bacterial enumeration. Since we had no indication about the gas mixtures of the commercialized samples, we chose to recondition aliquots under air or vacuum packaging to potentially enrich bacterial diversity and incubate them at 4 °C and 8 °C. Total counts and LAB counts were enumerated on days 1, 7, 14, 21, and 28. When PCA mesophilic counts reached about 7 log10 CFU per gram of ham, microbiotas were collected by mixing 25 g diced cooked ham in 75 mL peptone salt (AES, Rennes, France) for 2 min in a stomacher (Masticator, IUL Instruments, Barcelona, Spain). The homogenate was filtered through the bag filter and centrifuged through a filter from Nucleospin Plant II Midi kit (Macherey Nagel, Hoerdt, France) at 8000× g for 10 min at room temperature. The bacterial pellet was resuspended in 30 mL peptone salt and aliquoted as 1 mL with glycerol 15% and stored at −80 °C. In total, microbiotas from ten different cooked ham samples were recovered from dices conditioned either under air or vacuum.
2.3. Challenge Tests
Low-contaminated diced cooked ham containing 18 g kg−1 sodium chloride, and a reduced level of nitrite (25 mg kg−1; recommended max. level is 120 mg kg−1) were used for challenge tests. Those were produced as previously described [25]. Briefly, pork ham muscles were defatted, trimmed, and minced using a 20 mm grid. One kilogram of this grinding was mixed under vacuum with 100 g of brine containing water 74.1 g, nitrite salt 4.6 g (0.6%), sodium chloride salt 15.2 g, sodium ascorbate 0.6 g, and dextrose 5.5 g. The mix was melded as follows: components were mixed together in the brine water by stirring with a whisk until homogenization. Then, the brine was added to the meat by mixing (not by injection), and then the mix was vacuum-packed. Hams (2.5 kg) were cooked in a 100% humidity atmosphere (90 min at 55 °C, 60 min at 60 °C, and 235 min at 67 °C), cooled at ambient temperature (18 °C) for 20 min, and then stored at 3 °C. Ham cubes of 1 cm × 1 cm were prepared aseptically, aliquoted in 100 g portions, and stored vacuum-packed at −20 °C until use. Cooked ham dices were defrosted at 4 °C for 24 h, first inoculated with microbiota, and then with L. lactis fresh culture. Inoculation was performed by adding the microbial suspensions to ham dices placed in bags and hand-mixed for two minutes. Two different microbiotas were inoculated in low-contaminated diced cooked ham. Each challenge test was performed twice (two independent experiments). Microbiotas previously collected were defrosted rapidly at room temperature, diluted to a final concentration of 6 log10 CFU mL−1, inoculated at 4 log10 CFU·g−1 in diced cooked ham (1% v/w inoculation rate), and stored overnight at 4 °C for allowing bacterial recovery. L. lactis CH-HP15 inoculation was then performed with L. lactis fresh culture as follows. When the early stationary phase was reached, the culture was centrifuged. The bacterial pellet was rinsed in a sterile solution of NaCl 0.9% and resuspended at 9.3 log10 CFU·g−1; 0.5 mL were inoculated in 100 g dice cooked ham for an initial concentration of 7 log10 CFU·g−1. Diced cooked ham inoculated with microbiota but without L. lactis were also used as controls.
2.4. HP Treatments
Cooked ham dices were aliquoted as 20 g portions, vacuum-packed and high-pressure treated at 500 MPa for 5 min. This combination of pressure intensity and duration was chosen according to our previous study [25] for ensuring survival and further regrowth of L. lactis CH-HP15 cells essential for their protective effect. Samples were treated using high pressure in a 3 L vertical high-pressure pilot unit (ACB, Nantes, France) equipped with a temperature regulator device. The pressure transmitting fluid was distilled water. The samples were inserted into the vessel (whose internal temperature was regulated to 20 °C) and processed at a compression rate of 3.4 MPa s−1 until the targeted pressure was reached. The pressure level was held for 5 min, and decompression was nearly instantaneous (less than 2 s). Water temperature reached 24 °C at 500 MPa because of the adiabatic heating. Once treated, packs were then stored at 8 °C, and one portion was used on days 1, 5, 12, and 30 for bacterial enumeration and bacterial pellet collection for further DNA extraction. Unpressurized samples were also included as controls.
2.5. DNA Preparation and Amplicon Sequencing
Total genomic DNA was extracted from the bacterial pellet as described previously [28] using the PowerFood™ Microbial DNA Isolation kit (MoBio Laboratories Inc., Carlsbad, CA, USA) and the High Pure PCR Template Preparation kit (Roche Diagnostics Ltd, Burgess Hill, West Sussex, UK). For each sample, both DNA extracts were pooled. Then, this DNA sample was used as a template for three independent amplifications using either the 16S V3–V4 region of the rRNA encoding gene or an internal 280 bp fragment of the Gyrase B subunit encoding gene gyrB, as described previously [28]. All PCRs were performed in triplicate. Replicates were pooled, and the amplified DNA was purified with a QIAquick kit (Qiagen, Hilden, Germany). Amplicon size, quality, and quantity were checked on a DNA1000 chip (Agilent Technologies, Paris, France). The MiSeq Reagent Kit v.2 (2 × 250 paired-end reads, 15 Gb output) was used according to the manufacturer’s instructions for library preparation and sequencing on a MiSeq 2 (Illumina, San Diego, CA, USA). The quality of the obtained sequences were checked with FastQ files generated at the end of the run (MiSeq Reporter Software v.2.4, Illumina, San Diego, CA, USA) and additional PhiX Control (v.3, Illumina, San Diego, CA, USA). The corresponding pairs of sequences were then attributed to their respective samples using the individual multiplexing barcodes. Quality controls indicated a PHRED quality score of at least 30 for 94% of the reads and a median number of 65,520 ± 14,180 reads per sample and 95,860 ± 11,300 reads per sample for 16S rDNA and gyrB amplicons, respectively.
2.6. Operational Taxonomic Unit (OTU) Analysis and Accession Numbers
For each sample, paired-end sequences were then loaded in the FROGS (Find Rapidly OTUs with Galaxy Solutions, v.2.0) pipeline [29], checked for quality, and assembled. We retained merged sequences with a size of 280 ± 50 bp for gyrB and 450 ± 50 bp for the 16S rRNA gene. SWARM clustering [30] was applied on the assembled sequences using a maximal aggregation distance of three nucleotides for 16S rRNA gene sequences; for gyrB sequences, clustering was more stringent, with a maximal aggregation distance of two nucleotides in order to potentially assign OTUs to the subspecies level. After chimeras and spurious OTUs (low-abundance and low-frequency OTUs arising from sequencing artifacts) removal as described previously [28], the dataset comprised 69 OTUs for the 16S rDNA dataset and 252 OTUs for the gyrB dataset. Taxonomic assignment of 16S rDNA OTUs was performed with the Blastn+ algorithm v.2.10 [31] on the SILVA 128 SSU database [32], using a threshold of 97% identity for species assignment. For gyrB OTUs, it has been previously demonstrated that PCR amplification of gyrB can also recover the paralogous parE gene, which encodes the β subunit of DNA topoisomerase IV. In order to determine which of the two genes had been amplified for each species, the OTU sequences were blasted against the gyrB/parE databases established by Poirier and colleagues [28]. Both gyrB and parE OTUs were retained in our diversity analysis. We assigned taxonomy to the species level when 95% of a sequence matched over 90% of length coverage found in the database. The last step consisted of comparing the taxonomic assignments obtained from the 16S rDNA and the gyrB amplicon sequencing. The taxonomic assignment of the 16S rDNA-based OTUs were then homogenized and improved by those obtained from the gyrB analysis. This strategy was particularly applied to 16S rRNA gene OTUs with no clear assignment to the species-level (several species included at the threshold of 97% identity).
2.7. Beta-Diversity and Statistical Analysis
Bacterial diversity analyses were performed using the phyloseq package (v.1.24.2) of R [33]. For each of the samples, analysis of diversity was either performed at the OTU level or at the species level. In studies at the species level, OTUs with similar taxonomic assignments (both gyrB and 16S rDNA) were merged using the TAX_GLOM function of phyloseq package, and their abundance (number of reads) was averaged. This resulted in a dataset of 56 species with different taxonomic assignments. Similarly, comparative analysis between ham samples with different experimental conditions was performed after technical (16S rDNA/gyrB) and biological (biological repetition) data were merged with the phyloseq function MERGE_SAMPLES. In this case, species abundance was averaged between the four replicates. Bacterial diversity was compared among different groups of samples with permutational ANOVA, specifically using the adonis function within the vegan package [34].
3. Results
3.1. Diced Cooked Ham Selection for Microbiota Recovery
Preliminary tests were performed on diced cooked ham of different brands collected from local supermarkets. They were purchased as close as possible to the production date, based on an average shelf life of 20–25 days. They were then incubated under vacuum or air packaging at 4 °C or 8 °C. We observed a large variation of total viable counts between samples, ranging from 2 to 7 log10 CFU·g−1 one day after purchase, i.e., 2–7 days after production. No strong influence of packaging was observed on psychrophilic and mesophilic total counts, or MRS counts. Enumeration on MRS plates containing bromocresol green enabled us to estimate the putative diversity of LAB species present in dices. Only 1–3 types of colonies were detected, indicating a poor diversity among the most dominant LAB species.
Since our objective was to recover microbiota for further re-inoculation on cooked diced ham before HPP treatment, we aimed at collecting standardized quantities of bacteria, as diverse as possible, and enough DNA for further amplicon sequencing. For that purpose, we designed the sampling as follows: (i) diced cooked ham were sampled from supermarkets as close as possible to their production date; (ii) immediately after arrival in the laboratory, dices were reconditioned under vacuum packaging and under air to eventually increase the bacterial diversity to be collected; (iii) packs were then stored at 4 °C and analyzed every 7 days; (iv) when total viable counts reached about 7 log10 CFU·g−1, microbiotas were collected and stored for further analyses, and DNA was extracted for amplicon sequencing. Through this strategy, diced cooked ham samples from four different brands out of ten (further referred to as HAM_A to HAM_D samples) were obtained with enough bacterial diversity and DNA quality (Table 1). No significant difference in population level was observed between air and vacuum packaging, except in HAM_B, for which both PCA and MRS counts were about 1 log higher (respectively 7.97 vs. 6.53 log10 CFU·g−1 and 7.74 vs. 6.82 log10 CFU·g−1) after storage under air.

Table 1.
Description of ham specificities.
3.2. Four Different Diced Cooked Ham Samples Can Be Distinguished by the Diversity of Their Bacterial Communities
The bacterial diversity was estimated using a combination of two amplicon sequencing strategies, one using the V3–V4 region of the 16S rRNA gene and another one using an internal ~280 bp fragment of the gyrase B subunit encoding gene (gyrB) in order to improve taxonomic assignment of the OTUs to the species or sub-species level, as described previously [28]. These two strategies were also used as technical replicates for ensuring that no biases were obtained in the characterization of the four different microbiotas, as observed in Figure 1A. Only slight differences between both strategies were observed in the clustering of air samples of HAM_C and D. The average number of species detected per sample was 38 ± 5. As shown in Figure 1B, HAM_C and HAM_D samples showed a highly similar microbiota dominated by Firmicutes, in particular, Latilactobacillus sakei and Leuconostoc carnosum, two species commonly found on this type of cured meat products [35,36,37]. Furthermore, the storage type (under vacuum or air packaging) did not significantly influence the overall abundance of the identified species. On the other hand, the bacterial communities characterized from samples of HAM_A and HAM_B were significantly different from each other and from the aforementioned products, in particular with a clear dominance of Proteobacteria over Firmicutes (Figure 1C). Ham_A microbiota was mainly composed of Pseudomonadaceae and Enterobacteriaceae, with Pseudomonas lundensis and Serratia grimesii being the most abundant species for Proteobacteria and Carnobacterium maltaromaticum being the most abundant species for Firmicutes. Samples from Ham_B were revealed to contain the most original microbiota among the four types of ham, with the three most abundant 16S rDNA or gyrB OTUs being assigned to unknown species, as the identity percentage of these OTUs to gyrB genes in databases was lower than the ANI (Average Nucleotide Index) threshold of 95% classically used for bacterial species boundary [38]. The most abundant Firmicutes gyrB OTU was assigned to a Carnobacterium sp., showing only 89.0% identity (as best match) to gyrB gene of Carnobacterium funditum strain DSM 5970. Among Proteobacteria, one OTU was assigned to a Psychrobacter sp. with the closest match being 89.9% to the gyrB gene of Psychrobacter cryohalolentis FDAARGOS_308, a strain isolated from Siberian permafrost (GenBank: GCF_002208775.2). For the second Proteobacteria OTU, the best identity score found was 91.6 % with Vibrio sp. SM1977 gyrB gene, a species isolated from a coralline algae surface (GenBank: CP045699.1). It should be noted that the abundance of these two uncharacterized species (Psychrobacter sp. and Vibrio sp.) varied significantly according to the packaging type used for storage, which may correlate with their 10 times higher counts on air stored samples (Table 1), although we have no evidence for their cultivability in our growth conditions.
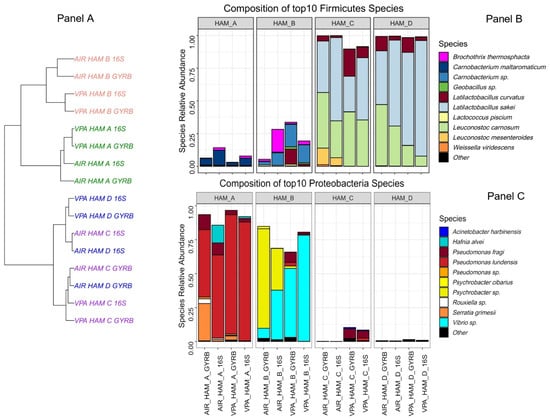
Figure 1.
Comparative bacterial community composition between the four brands of cooked ham analyzed. Panel (A) Cooked ham samples unsupervised clustering tree based on Bray-Curtis distance and Ward algorithm. Samples are colored according to the cooked ham brand. Both air (AIR) and vacuum packed (VPA) samples, as well as 16S rDNA-based or gyrB–based, analyses are shown. Panels (B,C) Barplot composition of the top 10 species identified among Firmicutes and Proteobacteria phyla, respectively. Species are plotted according to their relative abundance in the percentage of the whole microbiota. Novel genus nomenclature was applied for Latilactobacillus species (L. sakei and L. curvatus), as recently proposed by Zheng and colleagues [39]. Other refers to species not belonging to the top 10.
3.3. Combined Effect of HPP and Biopreservation on Bacterial Community Dynamics
For the further steps of our analysis dedicated to monitoring the effect of HPP and biopreservation by L. lactis CH-HP15 on the ham microbiota dynamics during storage at 8 °C, we decided to focus on the two different microbiotas from HAM_A and HAM_B. This was based on the fact that these two microbiotas showed a community structure enriched in bacterial species with high spoilage potential (Proteobacteria) in comparison to microbiotas of HAM_C and HAM_D enriched in Latilactobacillus; those latter species being generally rather considered as positive microbial components [20,21]. In addition, we chose HAM_B microbiota because of the unexpected presence of the two uncharacterized Psychrobacter and Vibrio species. Table 2 summarizes the estimated bacterial population level at different storage times after several processing treatments. Similarly, Figure 2 depicts the relative abundance of each species via 16S rDNA and gyrB amplicon sequencing.
Table 2.
Description of ham specificities.
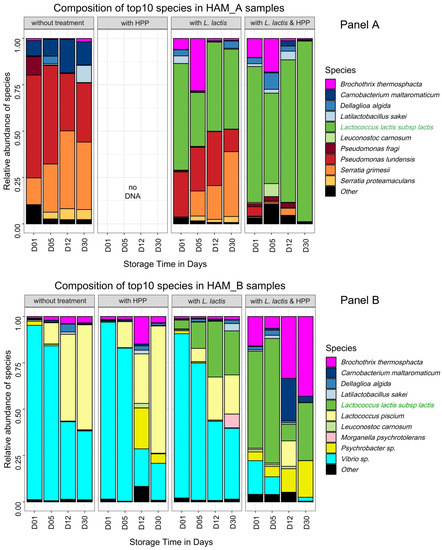
Figure 2.
Time course dynamic barplot composition of bacterial communities in nitrite-reduced cooked ham samples upon various treatments. Data for HAM_A and HAM_B samples are shown in Panels (A,B), respectively. Species are plotted according to their relative abundance in percentage of the whole microbiota. The L. lactis strain CH-HP15 used for biopreservation is highlighted in green in the left legend. Each bar plot is the mean of two biological replicates, each replicate value being the average of both 16S rDNA and gyrB amplicon sequencing data. As described in Figure 1 for Latilactobacillus species, novel genus nomenclature was applied for Dellaglioa algida (former Lactobacillus algidus), as recently proposed by Zheng and colleagues [39]. Other refers to species not belonging to the top 10.
The control samples without any processing treatment revealed that at day 1 after inoculation, the microbial communities were similar to the microbiota from HAM_A and HAM_B analyzed in Figure 1, showing that the dices prepared for the challenge-tests were indeed poorly contaminated and that the microbiota recovered with no strong bias from freezing. HAM_A microbiota grew rapidly under vacuum from ~4 log10 CFU·g−1 to reach ~7 log10 CFU·g−1 at day 5 and ~8 log10 CFU·g−1 at day 30. In contrast, HAM_B microbiota showed more reduced growth dynamics, reaching at most ~7 log10 CFU·g−1 during the whole storage period. Interestingly, the structure of the communities also changed during storage: in HAM_A samples, P. lundensis, a species abundant at the beginning of storage, was progressively overgrown by S. grimesii whereas, C. maltaromaticum, the third most abundant species, remained at a stable proportion within the community; in HAM_B, the dominant Vibrio sp. was also overgrown over time by Lactococcus piscium, although this latter species was largely subdominant (<0.1%) in the initial microbiota.
Processing with biopreservation by L. lactis CH-HP15 led to two different effects regarding samples from HAM_A and HAM_B. In HAM_A samples, the addition of L. lactis had a strong reduction effect on the abundance of Firmicutes species, in particular on C. maltaromaticum, which thus became largely subdominant. Interestingly, B. thermosphacta was temporally more abundant and thus presumably taking benefit from this change (Figure 2A). Meanwhile, the inoculation of L. lactis had no obvious effect on Proteobacteria species whose pattern was unchanged over time compared to control. Indeed, after 30 days of storage, while the microbiota had reached a population level above 8.5 log10 CFU·g−1, P. lundensis and S. grimesii species displayed a relative abundance of about 50% of the microbiota; the remaining part of the microbiota being covered by L. lactis, although this strain had been inoculated a thousand-fold higher than the original microbiota. In HAM_B, the pattern of the dominant Vibrio sp. was also not affected in comparison to the untreated samples. Indeed, largely dominant at day 1, its relative abundance decreased over time. However, the addition of L. lactis induced a possible competition with L. piscium, resulting over time in the equilibrium of both species. More generally, L. lactis CH-HP15 was less competitive in microbiota from HAM_B samples than in microbiota from HAM_A samples.
Whether or not biopreservation was applied, HPP treatment resulted in major changes in the growth dynamics of both microbiotas. HPP treatment alone on HAM_A samples reduced drastically the viability of the original microbiota, which remained below 4 log10 CFU·g−1 after 30 days of storage (Table 2). It is likely that bacterial cells were severely injured, indeed no DNA could be recovered from the bacterial pellet despite the use of several extraction protocol trials. Therefore, diversity analysis by amplicon sequencing could not be performed. Nevertheless, samples treated with both HPP and inoculation by L. lactis enabled better recovery of DNA and could provide an overview of the HPP treatment on the HAM_A original microbiota.
Proteobacteria species were the most impacted by HPP treatment, whereas L. lactis CH-HP15 could recover rapidly from this treatment reaching progressively a population level above 7.5 log10 CFU·g−1 after 30 days, and almost a complete domination of the microbiota. Interestingly, as described above on samples only treated with L. lactis, the addition of the bioprotective microorganism increased temporally (up to 12 days of storage) the relative proportion of B. thermosphacta, thereby indicating that this latter species is quite resistant to HPP treatment. Unlike microbiota from HAM_A samples, that from HAM_B samples recovered rapidly from HPP treatment alone, almost reaching the population level of untreated samples at 30 days (6 log10 CFU·g−1 on average). The community structure was itself barely affected by the HPP treatment with only a higher proportion of Psychrobacter sp. at longer storage time in comparison to untreated samples. Processing with both HPP and L. lactis inoculation confirmed these observations as the population level, combining both initial microbiota and L. lactis, recovered rapidly (12 days) to 7.0 log10 CFU·g−1 and raised further to ~10 log10 CFU·g−1 at 30 days, which is almost 4 log higher than the level in untreated ham. However, the HPP treatment was more favorable to Firmicutes species, in the end, leading to domination of L. lactis CH-HP15 together with B. thermosphacta.
4. Discussion
The use of an appropriate HPP treatment to preserve perishable food has been the focus of many studies and reviews [22,40]. HPP is being used to enhance food safety by reducing the microbial development in final products while maintaining the best nutritional and sensorial values to a level acceptable to consumers [17]. However, the effect of HPP on microbial growth dynamics and community structures has not been widely studied, and it is not known whether hurdle strategies should be applied in combination with HPP for efficient microbiota inactivation or stabilization. Indeed, recent work carried out with a simplified ham microbiota (five species, including Listeria monocytogenes, L. sakei, B. thermosphacta, C. maltaromaticum, and Leuconostoc gelidum) showed that HPP treatment is not sufficient to inhibit growth recovery of the microbiota over long storage time [24]. This points out that there is a clear gap in our knowledge on the HPP efficiency towards various microbial communities, which may be present on cooked ham. Although the technology could be a very promising strategy to improve the safety of nitrite-reduced cooked ham, it is necessary to investigate whether it should be used as a hurdle with the combination of other strategies, such as biopreservation [25].
Our strategy was to recover different microbiotas for reusing in performing challenge tests. As previously demonstrated by Raimondi and colleagues [35], the microbiota of the four types of cooked ham was poorly diverse but highly variable. Some cooked hams with poor diversity were dominated by L. sakei, Latilactobacillus curvatus, and L. carnosum, and their microbiotas were not considered further, as HPP treatments were previously demonstrated to be efficient against the growth of these species [9]. On the other hand, cooked hams from two other producers were characterized by a more diverse microbiota. These microbiotas were composed of a quite different mixture of species from Firmicutes and Proteobacteria phyla, including not yet characterized dominant species. Therefore, these two types of microbiotas were chosen as models to test the efficiency of HPP and biopreservation.
Firstly, our results demonstrate that the reduction of viable cell populations by HPP is slightly more effective on Proteobacteria species than on Firmicutes species resulting in a small shift of the dominant population from Proteobacteria to Firmicutes when HPP is applied. This finding corroborates many previous observations made by comparing individual species of Gram-negative and Gram-positive pathogenic bacteria resistance to HPP, for instance ([22] for a review). Although the cell surface morphology of both types of bacteria can explain such differences, it is interesting to notice that the recovery of cells after HPP is species (and per se microbiota) dependent. Although both microbiotas from HAM_A and HAM_B were re-inoculated at the same population level before HPP treatment, that from HAM_A, composed of P. lundensis and S. grimesii, were revealed as more sensitive (no or almost no recovery) to HPP than that from HAM_B composed of uncharacterized Psychrobacter and Vibrio species. The species or even strain-dependent resistance to HPP has already been pointed out for Firmicutes, in particular for L. monocytogenes and Staphylococcus aureus [13]. The underlying mechanisms are not fully understood as HPP resistance and recovery of bacteria also depend on the cell physiology of bacteria present on the food matrix before the treatment. As well, the matrix composition may influence bacterial recovery [41], as was shown for fat content influencing L. monocytogenes recovery from thermal inactivation [42]. Furthermore, the capacity of the bacteria to resist HPP treatment should be dissociated from the ability of the bacterial cells to recover and thrive during the storage conditions. Our data are a good illustration of this phenomenon. For instance, S. grimesii was revealed as more competitive than P. lundensis to grow during 30 days at low temperature and perhaps under vacuum packaging, leading to a switch of the two species during storage. Similarly, L. piscium, a sub-dominant species in original HAM_B microbiota, could outcompete the initial dominant Vibrio species.
Another finding from our work is that the L. lactis strain used for biopreservation is not competitive towards ham original microbiota. However, we noticed that it has perceivable effects on the reduction of other Firmicutes species (e.g., C. maltaromaticum), perhaps due to the production of nisin. Indeed, this bacterium has been shown in previous studies to harbor sensitivity to this bacteriocin in vacuum-packed meat [24,43]. Albeit the L. lactis CH-HP15 strain was inoculated with a level three orders of magnitude higher than the original ham microbiota, the strain was found to be between 25% and 50% of overall relative abundance in HAM_A and HAM_B, respectively. We previously observed that the inactivation level of this strain after HPP was more important than that of other LAB species, which was compensated by its better ability to recover and rapidly regrow after the treatment [25]. The lack of L. lactis competitiveness is probably due to the specific ecology of ham. Indeed, L. lactis CH-HP15 was shown to be able to grow in sterile diced cooked ham at 8 °C, reaching 9 log10 CFU·g−1 within 5 days with an initial inoculum of 6 log10 CFU·g−1 [25]. In the present study, the presence of species belonging to the natural microbiota and thus potentially better adapted may explain the lack of fitness of L. lactis CH-HP15. Nevertheless, the combination of a high level of inoculation of L. lactis with HPP treatment leads to an efficient stabilization of the original cooked HAM_A microbiota by limiting the growth of P. lundensis and S. grimesii. Unlike this situation, results were different for HAM_B samples for which the hurdle strategy could not trigger, over the storage time, the outcompetition of L. lactis versus the original HAM_B microbiota. Therefore, it can be concluded that for the microbiota of HAM_B, the hurdle strategy failed in stabilizing and inactivating microbial growth.
Furthermore, our results show that B. thermosphacta is a species with very strong recovery dynamics following HPP treatment. Such an observation has already been made by Teixeira and colleagues using the simplified ham microbiota described above [24]. This finding raises the question of HPP treatment benefit on B. thermosphacta containing microbiota as this species is a well-known meat spoilage micro-organism [44].
5. Conclusions
From our work, we thus conclude that both HPP treatment and L. lactis-based biopreservation are strongly microbiota-dependent, and thus, the value of this strategy requires a specific assessment for each type of cooked ham production. We recommend that HPP treatment should be evaluated not only for pathogenic bacteria but also on putative spoilage bacteria in order to estimate the specific selection of these undesirable micro-organisms, in particular B. thermosphacta.
Author Contributions
Conceptualization, M.Z., S.C., M.C.-V. and S.G.; methodology, M.R. and A.R.; ham sample and microbiota collection, J.-L.M. and S.R.; nitrite reduced ham preparation, S.G., V.A. and N.M.; challenge-tests, G.C.; DNA extraction, library preparation, and amplicon sequencing, validation, S.C. and M.Z.; formal analysis, S.C.; data curation, S.C.; writing—original draft preparation, M.Z. and S.C.; writing—review and editing, M.Z. and S.C.; visualization, S.C.; supervision, M.Z. and S.C.; project administration, M.Z.; funding acquisition, M.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the French National Research Agency (ANR) for their financial support in the framework of the BLAC-HP project (ANR-14-CE20-0004).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The merged (assembled) sequences have been deposited in the Sequence Read Archive database under the accession numbers SAMN13761947 to SAMN13762161, corresponding to BioProject PRJNA599607.
Acknowledgments
The authors express their gratitude to the INRAE MIGALE bioinformatics facility (MIGALE, INRAE, 2020. Migale bioinformatics Facility, doi: 10.15454/1.5572390655343293E12) for providing computational resources and data storage. We also thank the INRA @BRIDGe platform for carrying out the MiSeq sequencing runs.
Conflicts of Interest
None. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Alahakoon, A.U.; Jayasena, D.D.; Ramachandra, S.; Jo, C. Alternatives to nitrite in processed meat: Up to date. Trends Food Sci. Technol. 2015, 45, 37–49. [Google Scholar] [CrossRef]
- Majou, D.; Christieans, S. Mechanisms of the bactericidal effects of nitrate and nitrite in cured meats. Meat Sci. 2018, 145, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Sindelar, J.J.; Milkowski, A.L. Human safety controversies surrounding nitrate and nitrite in the diet. Nitric Oxide Biol. Chem. 2012, 26, 259–266. [Google Scholar] [CrossRef] [PubMed]
- EFSA. EFSA Explains Risk Assessment: Nitrites and Nitrates Added to Food. 2017. Available online: https://op.europa.eu/en/publication-detail/-/publication/20f25e61-5562-11e7-a5ca-01aa75ed71a1 (accessed on 1 December 2021).
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Domenico, A.D.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; et al. Re-evaluation of potassium nitrite (E 249) and sodium nitrite (E 250) as food additives. EFSA J. 2017, 15, e04786. [Google Scholar] [CrossRef]
- Mortensen, A.; Aguilar, F.; Crebelli, R.; Domenico, A.D.; Dusemund, B.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; Lambré, C.; et al. Re-evaluation of sodium nitrate (E 251) and potassium nitrate (E 252) as food additives. EFSA J. 2017, 15, e04787. [Google Scholar] [CrossRef]
- Roobab, U.; Khan, A.W.; Lorenzo, J.M.; Arshad, R.N.; Chen, B.-R.; Zeng, X.-A.; Bekhit, A.E.-D.; Suleman, R.; Aadil, R.M. A systematic review of clean-label alternatives to synthetic additives in raw and processed meat with a special emphasis on high-pressure processing (2018–2021). Food Res. Int. 2021, 150, 110792. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Serra-Castelló, C.; Dalgaard, P.; Garriga, M.; Jofré, A. New insights on Listeria monocytogenes growth in pressurised cooked ham: A piezo-stimulation effect enhanced by organic acids during storage. Int. J. Food Microbiol. 2019, 290, 150–158. [Google Scholar] [CrossRef]
- Han, Y.; Jiang, Y.; Xu, X.; Sun, X.; Xu, B.; Zhou, G. Effect of high pressure treatment on microbial populations of sliced vacuum-packed cooked ham. Meat Sci. 2011, 88, 682–688. [Google Scholar] [CrossRef]
- Hereu, A.; Dalgaard, P.; Garriga, M.; Aymerich, T.; Bover-Cid, S. Analysing and modelling the growth behaviour of Listeria monocytogenes on RTE cooked meat products after a high pressure treatment at 400 MPa. Int. J. Food Microbiol. 2014, 186, 84–94. [Google Scholar] [CrossRef]
- Jofré, A.; Garriga, M.; Aymerich, T. Inhibition of Listeria monocytogenes in cooked ham through active packaging with natural antimicrobials and high-pressure processing. J. Food Prot. 2007, 70, 2498–2502. [Google Scholar] [CrossRef]
- Jofré, A.; Aymerich, T.; Garriga, M. Assessment of the effectiveness of antimicrobial packaging combined with high pressure to control Salmonella sp. in cooked ham. Food Control 2008, 19, 634–638. [Google Scholar] [CrossRef]
- Jofré, A.; Garriga, M.; Aymerich, T. Inhibition of Salmonella sp. Listeria monocytogenes and Staphylococcus aureus in cooked ham by combining antimicrobials, high hydrostatic pressure and refrigeration. Meat Sci. 2008, 78, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Koseki, S.; Mizuno, Y.; Yamamoto, K. Predictive modelling of the recovery of Listeria monocytogenes on sliced cooked ham after high pressure processing. Int. J. Food Microbiol. 2007, 119, 300–307. [Google Scholar] [CrossRef] [PubMed]
- López-Caballero, M.E.; Carballo, J.; Jiménez-Colmenero, F. Microbiological changes in pressurized, prepackaged sliced cooked ham. J. Food Prot. 1999, 62, 1411–1415. [Google Scholar] [CrossRef] [Green Version]
- Marcos, B.; Jofré, A.; Aymerich, T.; Monfort, J.M.; Garriga, M. Combined effect of natural antimicrobials and high pressure processing to prevent Listeria monocytogenes growth after a cold chain break during storage of cooked ham. Food Control 2008, 19, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Pietrasik, Z.; Gaudette, N.J.; Johnston, S.P. The use of high pressure processing to enhance the quality and shelf life of reduced sodium naturally cured restructured cooked hams. Meat Sci. 2016, 116, 102–109. [Google Scholar] [CrossRef]
- Pingen, S.; Sudhaus, N.; Becker, A.; Krischek, C.; Klein, G. High pressure as an alternative processing step for ham production. Meat Sci. 2016, 118, 22–27. [Google Scholar] [CrossRef]
- Stiles, M.E. Biopreservation by lactic acid bacteria. Antonie Van Leeuwenhoek 1996, 70, 331–345. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Co-culture experiments demonstrate the usefulness of Lactobacillus sakei 10A to prolong the shelf-life of a model cooked ham. Int. J. Food Microbiol. 2006, 108, 68–77. [Google Scholar] [CrossRef]
- Vermeiren, L.; Devlieghere, F.; Debevere, J. Evaluation of meat born lactic acid bacteria as protective cultures for the biopreservation of cooked meat products. Int. J. Food Microbiol. 2004, 96, 149–164. [Google Scholar] [CrossRef]
- de Oliveira, T.L.C.; Ramos, A.L.S.; Ramos, E.M.; Piccoli, R.H.; Cristianini, M. Natural antimicrobials as additional hurdles to preservation of foods by high pressure processing. Trends Food Sci. Technol. 2015, 45, 60–85. [Google Scholar] [CrossRef]
- Liu, G.; Wang, Y.; Gui, M.; Zheng, H.; Dai, R.; Li, P. Combined effect of high hydrostatic pressure and enterocin LM-2 on the refrigerated shelf life of ready-to-eat sliced vacuum-packed cooked ham. Food Control 2012, 24, 64–71. [Google Scholar] [CrossRef]
- Teixeira, J.S.; Repková, L.; Gänzle, M.G.; McMullen, L.M. Effect of Pressure, reconstituted RTE meat microbiota, and antimicrobials on survival and post-pressure growth of Listeria monocytogenes on ham. Front. Microbiol. 2018, 9, 1979. [Google Scholar] [CrossRef]
- Ramaroson, M.; Guillou, S.; Rossero, A.; Rezé, S.; Anthoine, V.; Moriceau, N.; Martin, J.-L.; Duranton, F.; Zagorec, M. Selection procedure of bioprotective cultures for their combined use with High Pressure Processing to control spore-forming bacteria in cooked ham. Int. J. Food Microbiol. 2018, 276, 28–38. [Google Scholar] [CrossRef]
- Modugno, C.; Kmiha, S.; Simonin, H.; Aouadhi, C.; Diosdado Cañizares, E.; Lang, E.; André, S.; Mejri, S.; Maaroufi, A.; Perrier-Cornet, J.M. High pressure sensitization of heat-resistant and pathogenic foodborne spores to nisin. Food Microbiol. 2019, 84, 103244. [Google Scholar] [CrossRef]
- Najjari, A.; Ouzari, H.; Boudabous, A.; Zagorec, M. Method for reliable isolation of Lactobacillus sakei strains originating from Tunisian seafood and meat products. Int. J. Food Microbiol. 2008, 121, 342–351. [Google Scholar] [CrossRef]
- Poirier, S.; Rué, O.; Peguilhan, R.; Coeuret, G.; Zagorec, M.; Champomier-Vergès, M.-C.; Loux, V.; Chaillou, S. Deciphering intra-species bacterial diversity of meat and seafood spoilage microbiota using gyrB amplicon sequencing: A comparative analysis with 16S rDNA V3-V4 amplicon sequencing. PLoS ONE 2018, 13, e0204629. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinforma. Oxf. Engl. 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Mahé, F.; Rognes, T.; Quince, C.; de Vargas, C.; Dunthorn, M. Swarm: Robust and fast clustering method for amplicon-based studies. Peer J. 2014, 2, e593. [Google Scholar] [CrossRef] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 1 December 2021).
- Raimondi, S.; Luciani, R.; Sirangelo, T.M.; Amaretti, A.; Leonardi, A.; Ulrici, A.; Foca, G.; D’Auria, G.; Moya, A.; Zuliani, V.; et al. Microbiota of sliced cooked ham packaged in modified atmosphere throughout the shelf life: Microbiota of sliced cooked ham in MAP. Int. J. Food Microbiol. 2019, 289, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Vasilopoulos, C.; De Maere, H.; De Mey, E.; Paelinck, H.; De Vuyst, L.; Leroy, F. Technology-induced selection towards the spoilage microbiota of artisan-type cooked ham packed under modified atmosphere. Food Microbiol. 2010, 27, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zagdoun, M.; Coeuret, G.; N’Dione, M.; Champomier-Vergès, M.-C.; Chaillou, S. Large microbiota survey reveals how the microbial ecology of cooked ham is shaped by different processing steps. Food Microbiol. 2020, 91, 103547. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.-L.; Xie, B.-B.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhou, J.; Oren, A.; Zhang, Y.-Z. A proposed genus boundary for the prokaryotes based on genomic insights. J. Bacteriol. 2014, 196, 2210–2215. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Yamamoto, K. Food processing by high hydrostatic pressure. Biosci. Biotechnol. Biochem. 2017, 81, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Tassou, C.C.; Galiatsatou, P.; Samaras, F.J.; Mallidis, C.G. Inactivation kinetics of a piezotolerant Staphylococcus aureus isolated from high-pressure-treated sliced ham by high pressure in buffer and in a ham model system: Evaluation in selective and non-selective medium. Innov. Food Sci. Emerg. Technol. 2007, 8, 478–484. [Google Scholar] [CrossRef]
- Verheyen, D.; Govaert, M.; Seow, T.K.; Ruvina, J.; Mukherjee, V.; Baka, M.; Skåra, T.; Van Impe, J.F.M. The complex effect of food matrix fat content on thermal inactivation of Listeria monocytogenes: Case study in emulsion and gelled emulsion model systems. Front. Microbiol. 2020, 10, 3149. [Google Scholar] [CrossRef]
- La Storia, A.; Ferrocino, I.; Torrieri, E.; Di Monaco, R.; Mauriello, G.; Villani, F.; Ercolini, D. A combination of modified atmosphere and antimicrobial packaging to extend the shelf-life of beefsteaks stored at chill temperature. Int. J. Food Microbiol. 2012, 158, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Illikoud, N.; Gohier, R.; Werner, D.; Barrachina, C.; Roche, D.; Jaffrès, E.; Zagorec, M. Transcriptome and volatilome analysis during growth of Brochothrix thermosphacta in food: Role of food substrate and strain specificity for the expression of spoilage functions. Front. Microbiol. 2019, 10, 2527. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).