
Organic and Inorganic Amendments Shape Bacterial Indicator Communities That Can, In Turn, Promote Rice Yield

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
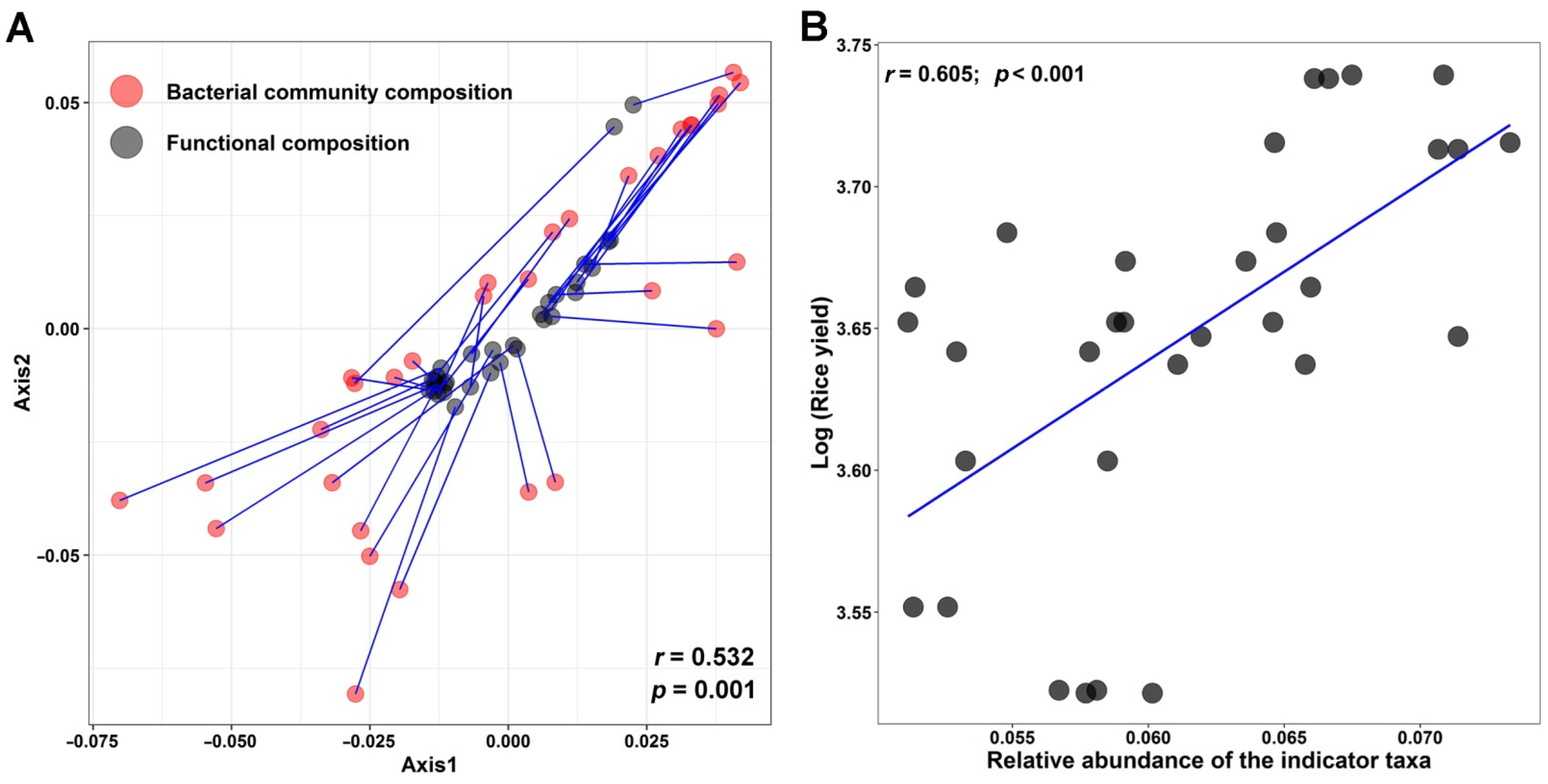{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Field Site and Experiment Description
2.2. Soil Sampling and Analysis
2.3. Soil Enzyme Activities and Microbial Biomass Analysis
2.4. DNA Extraction, PCR Amplification, and MiSeq Sequencing
2.5. Processing High-Throughput Sequencing Data
2.6. Bacterial Community Function Predictions
2.7. Random Forest Model
2.8. Statistical Analysis
3. Results
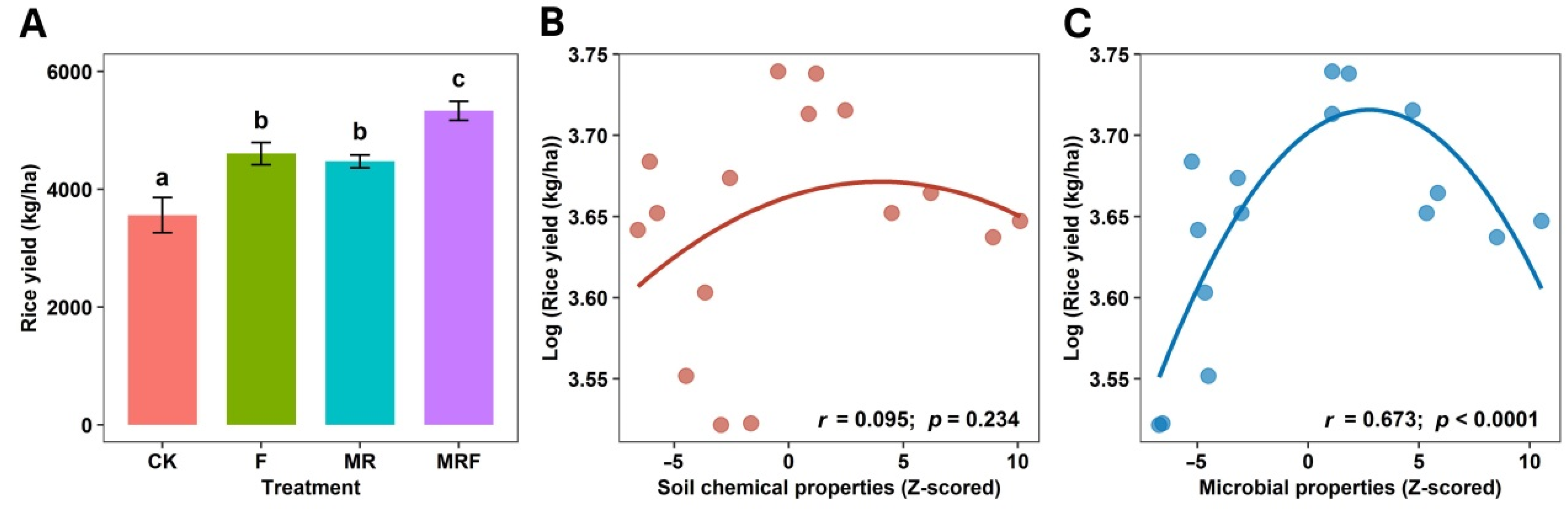
3.1. Crop Yield, Soil Chemical and Biological Properties under Different Fertilization Regimes
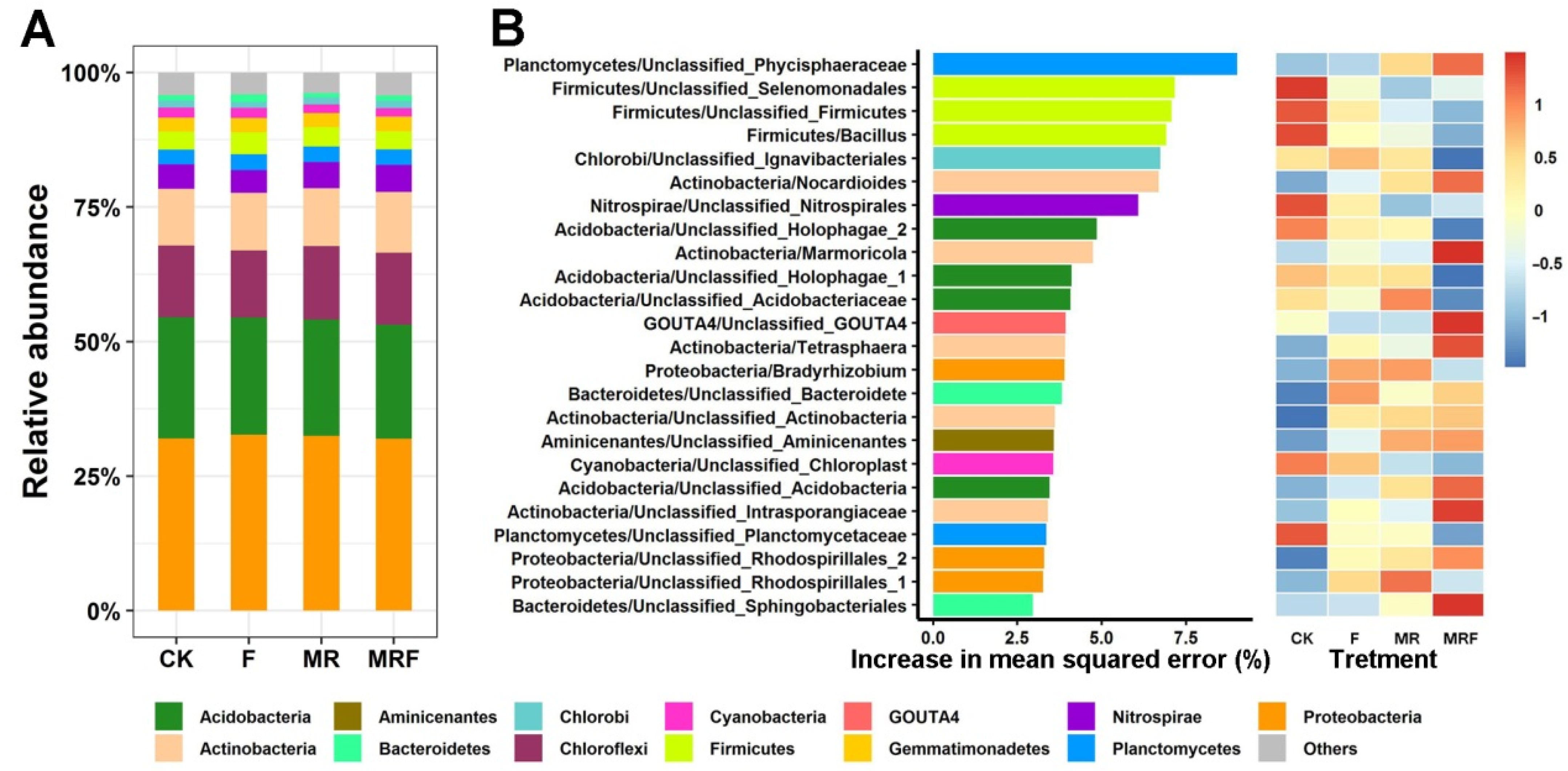
3.2. Taxonomic Distribution of the Bacterial Communities under Different Fertilization Regimes
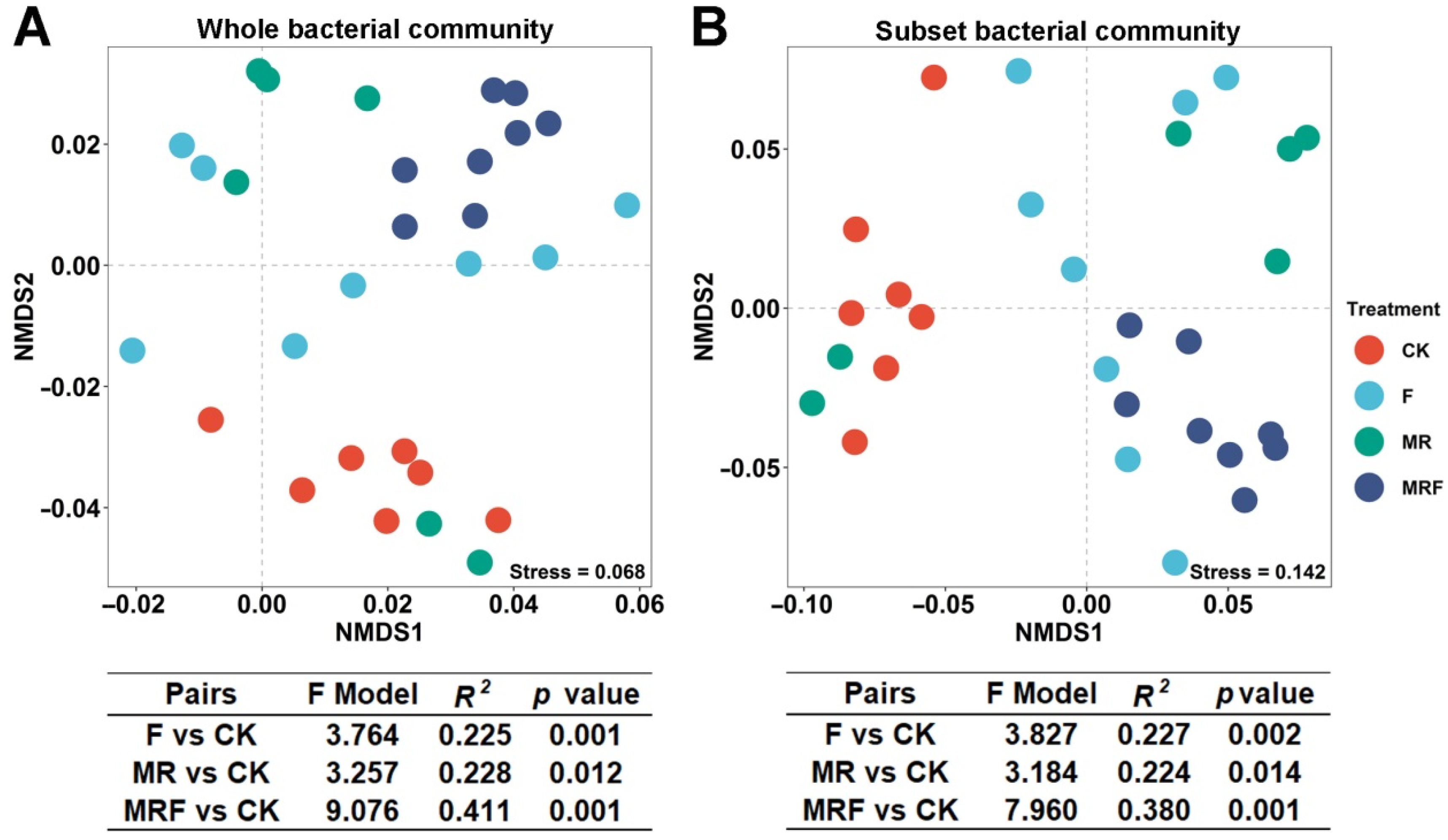
3.3. Variation in Bacterial Community Composition among Different Fertilization Regimes
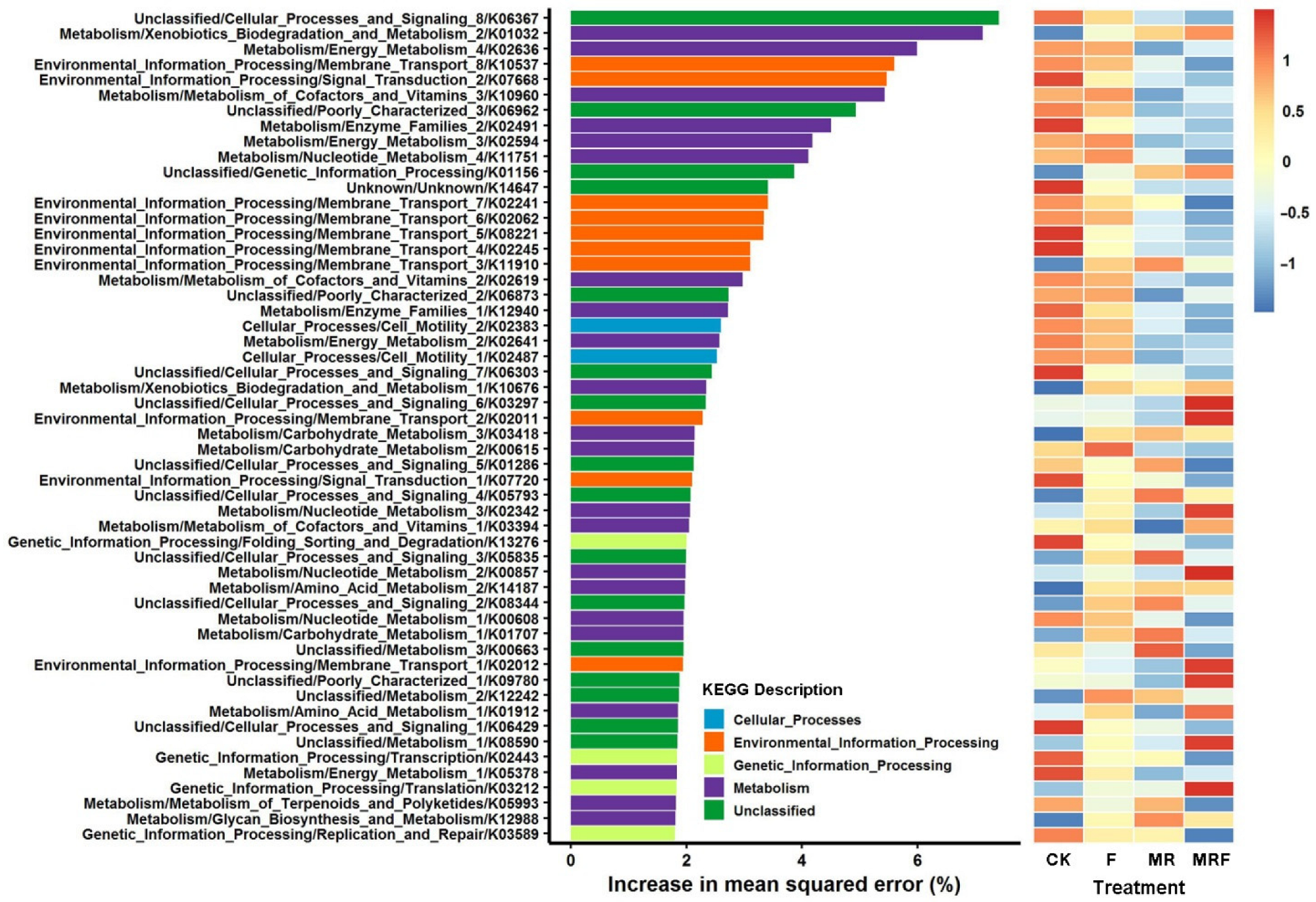
3.4. Dynamic Change in Compositions of Bacterial Metabolic Functions under Different Fertilizations
3.5. Relationship between Bacterial Indicator Community, Community Function, Soil Nutrient Cycling, and Crop Yield
4. Discussion
4.1. Microbial Properties of the Soil Community Reflect Crop Yield Better Than Soil Chemical Characteristics
4.2. Organic and Inorganic Amendments Indirectly Affect Crop Yield by Shaping the Bacterial Indicator Communities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, Z.H.; Liu, Q.S.; Li, Z.Y.; Cheng, W.; Sun, J.M.; Guo, Z.H.; Li, Y.M.; Zhou, J.Q.; Meng, D.L.; Li, H.B.; et al. Environmental factors shaping the diversity of bacterial communities that promote rice production. BMC Microbiol. 2018, 18, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food dsemand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savci, S. An agricultural pollutant: Chemical fertilizer. Int. J. Environ. Sci. Dev. 2012, 3, 73. [Google Scholar] [CrossRef] [Green Version]
- Malhi, S.S.; Nyborg, M.; Solberg, E.D.; McConkey, B.; Dyck, M.; Puurveen, D. Long-term straw management and N fertilizer rate effects on quantity and quality of organic C and N and some chemical properties in two contrasting soils in Western Canada. Biol. Fert. Soils 2011, 47, 785–800. [Google Scholar] [CrossRef]
- Wang, W.; Chen, C.L.; Wu, X.H.; Xie, K.J.; Yin, C.M.; Hou, H.J.; Xie, X.L. Effects of reduced chemical fertilizer combined with straw retention on greenhouse gas budget and crop production in double rice fields. Biol. Fert. Soils 2019, 55, 89–96. [Google Scholar] [CrossRef]
- Tejada, M.; Garcia, C.; Gonzalez, J.L.; Hernandez, M.T. Use of organic amendment as a strategy for saline soil remediation: Influence on the physical, chemical and biological properties of soil. Soil Biol. Biochem. 2006, 38, 1413–1421. [Google Scholar] [CrossRef]
- Xin, X.L.; Zhang, J.B.; Zhu, A.N.; Zhang, C.Z. Effects of long-term (23 years) mineral fertilizer and compost application on physical properties of fluvo-aquic soil in the North China Plain. Soil Till. Res. 2016, 156, 166–172. [Google Scholar] [CrossRef]
- Philippot, L.; Spor, A.; Henault, C.; Bru, D.; Bizouard, F.; Jones, C.M.; Sarr, A.; Maron, P.A. Loss in microbial diversity affects nitrogen cycling in soil. ISME J. 2013, 7, 1609–1619. [Google Scholar] [CrossRef]
- Jeyarajan, R.; Nakkeeran, S. Exploitation of microorganisms and viruses as biocontrol agents for crop disease management. In Biocontrol Potential and Its Exploitation in Sustainable Agriculture; Springer: Berlin/Heidelberg, Germany, 2000; pp. 95–116. [Google Scholar]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fert. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Zhan, Y.; Liu, W.; Bao, Y.; Zhang, J.; Petropoulos, E.; Li, Z.; Lin, X.; Feng, Y. Fertilization shapes a well-organized community of bacterial decomposers for accelerated paddy straw degradation. Sci. Rep. 2018, 8, 7981. [Google Scholar] [CrossRef]
- Knelman, J.E.; Nemergut, D.R. Changes in community assembly may shift the relationship between biodiversity and ecosystem function. Front. Microbiol. 2014, 5, 424. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.Y.; Feng, Y.Z.; Stegen, J.C.; Wu, M.; Chen, R.R.; Liu, W.J.; Zhang, J.W.; Li, Z.P.; Lin, X.G. Straw chemistry links the assembly of bacterial communities to decomposition in paddy soils. Soil Biol. Biochem. 2020, 148, 107866. [Google Scholar] [CrossRef]
- Lu, P.N.; Bainard, L.D.; Ma, B.; Liu, J.H. Bio-fertilizer and rotten straw amendments alter the rhizosphere bacterial community and increase oat productivity in a saline-alkaline environment. Sci. Rep. 2020, 10, 19896. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.Y.; Guo, Z.Y.; Chen, R.R.; Wu, M.; Li, Z.P.; Lin, X.G.; Feng, Y.Z. Functional community composition has less environmental variability than taxonomic composition in straw-degrading bacteria. Biol. Fert. Soils 2020, 56, 869–874. [Google Scholar] [CrossRef]
- Louca, S.; Polz, M.F.; Mazel, F.; Albright, M.B.N.; Huber, J.A.; O’Connor, M.I.; Ackermann, M.; Hahn, A.S.; Srivastava, D.S.; Crowe, S.A.; et al. Function and functional redundancy in microbial systems. Nat. Ecol. Evol. 2018, 2, 936–943. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, J.; Lu, Q.; Fang, Z.Y.; Huang, Q.W.; Zhang, R.F.; Li, R.; Shen, B.; Shen, Q.R. Effects of organic-inorganic compound fertilizer with reduced chemical fertilizer application on crop yields, soil biological activity and bacterial community structure in a rice-wheat cropping system. Appl. Soil Ecol. 2016, 99, 1–12. [Google Scholar] [CrossRef]
- Yao, T.Y.; Chen, R.R.; Zhang, J.W.; Feng, Y.Z.; Huang, M.S.; Lin, X.G. Divergent patterns of microbial community composition shift under two fertilization regimes revealed by responding species. Appl. Soil Ecol. 2020, 154, 103590. [Google Scholar] [CrossRef]
- Kyuma, K. Paddy Soil Science; Kyoto University Press: Kyoto, Japan, 2004. [Google Scholar]
- Lu, R.K. Analytical Methods of Soil and Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Gosewinkel, U.; Broadbent, F.E. Conductimetric determination of soil urease activity. Commun. Soil Sci. Plan 1984, 15, 1377–1389. [Google Scholar] [CrossRef]
- Frankenberger, W.T.; Johanson, J.B. Method of measuring invertase activity in soils. Plant Soil 1983, 74, 301–311. [Google Scholar] [CrossRef]
- Alef, K.; Nannipieri, P. Methods in Applied Soil Microbiology and Biochemistry; Academic Press: New York, NY, USA, 1995; pp. 335–337. [Google Scholar]
- Serra-wittling, C.; Houot, S.; Barriuso, E. Soil enzymatic response to addition of municipal solid-waste compost. Biol. Fert. Soils 1995, 20, 226–236. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass-C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Joergensen, R.G. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEC value. Soil Biol. Biochem. 1996, 28, 25–31. [Google Scholar] [CrossRef]
- Joergensen, R.G.; Mueller, T. The fumigation-extraction method to estimate soil microbial biomass: Calibration of the kEN value. Soil Biol. Biochem. 1996, 28, 33–37. [Google Scholar] [CrossRef]
- Feng, Y.Z.; Chen, R.R.; Hu, J.L.; Zhao, F.; Wang, J.H.; Chu, H.Y.; Zhang, J.B.; Dolfing, J.; Lin, X.G. Bacillus asahii comes to the fore in organic manure fertilized alkaline soils. Soil Biol. Biochem. 2015, 81, 186–194. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Tang, H.Y.; Zhu, J.G.; Lin, X.G.; Feng, Y.Z. Effects of elevated ground-level ozone on paddy soil bacterial community and assembly mechanisms across four years. Sci. Total Environ. 2019, 654, 505–513. [Google Scholar] [CrossRef]
- Jiao, S.; Chen, W.; Wang, J.; Du, N.; Li, Q.; Wei, G. Soil microbiomes with distinct assemblies through vertical soil profiles drive the cycling of multiple nutrients in reforested ecosystems. Microbiome 2018, 6, 146. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.K.; Halpern, A.L.; Beeson, K.; Tran, B.; Venter, J.C.; Martiny, J.B.H. It’s all relative: Ranking the diversity of aquatic bacterial communities. Environ. Microbiol. 2008, 10, 2200–2210. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Walsh, D.C.I. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Oksanen, J. Multivariate analysis of ecological communities in R: Vegan tutorial. R Package Version 2011, 1, 1–43. [Google Scholar]
- Conrad, J.P. The occurrence and origin of urease like activities in soils. Soil Sci. 1942, 54, 367–380. [Google Scholar] [CrossRef]
- Stemmer, M.; Gerzabek, M.H.; Kandeler, E. Invertase and xylanase activity of bulk soil and particle-size fractions during maize straw decomposition. Soil Biol. Biochem. 1999, 31, 9–18. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of phosphatase enzymes in soil. In Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling; Bünemann, E., Oberson, A., Frossard, E., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 215–243. [Google Scholar]
- Garcia, C.; Hernandez, T.; Costa, F. Potential use of dehydrogenase activity as an index of microbial activity in degraded soils. Commun. Soil Sci. Plan 1997, 28, 123–134. [Google Scholar] [CrossRef]
- Mi, W.H.; Sun, Y.; Xia, S.Q.; Zhao, H.T.; Mi, W.T.; Brookes, P.C.; Liu, Y.L.; Wu, L.H. Effect of inorganic fertilizers with organic amendments on soil chemical properties and rice yield in a low-productivity paddy soil. Geoderma 2018, 320, 23–29. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, X.; Bao, E.C.; Li, J.S.; Zou, Z.R.; Cao, K. Bio-organic fertilizer with reduced rates of chemical fertilization improves soil fertility and enhances tomato yield and quality. Sci. Rep. 2020, 10, 177. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.H.; Liu, X.; Wang, Z.C.; Liang, Z.W.; Wang, M.M.; Liu, M.; Suarez, D.L. Interactive effects of pH, EC and nitrogen on yields and nutrient absorption of rice (Oryza sativa L.). Agric. Water Manag. 2017, 194, 48–57. [Google Scholar] [CrossRef]
- Feng, Y.Z.; Guo, Z.Y.; Zhong, L.H.; Zhao, F.; Zhang, J.B.; Lin, X.G. Balanced fertilization decreases environmental filtering on soil bacterial community assemblage in north China. Front. Microbiol. 2017, 8, 2376. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; He, M. The application of exogenous cellulase to improve soil fertility and plant growth due to acceleration of straw decomposition. Bioresour. Technol. 2010, 101, 3724–3731. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Shi, S.H.; Tian, L.; Nasir, F.; Bahadur, A.; Batool, A.; Luo, S.S.; Yang, F.; Wang, Z.C.; Tian, C.J. Response of microbial communities and enzyme activities to amendments in saline-alkaline soils. Appl. Soil Ecol. 2019, 135, 16–24. [Google Scholar] [CrossRef]
- Liu, X.M.; Li, Q.; Liang, W.J.; Jiang, Y. Distribution of soil enzyme activities and microbial biomass along a latitudinal gradient in farmlands of Songliao Plain, Northeast China. Pedosphere 2008, 18, 431–440. [Google Scholar] [CrossRef]
- Priyadharsini, P.; Muthukumar, T. Interactions between arbuscular mycorrhizal fungi and potassium-solubilizing microorganisms on agricultural productivity. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V.S., Maurya, B.R., Verma, J.P., Meena, R.S., Eds.; Springer: New Delhi, India, 2016; pp. 111–125. [Google Scholar]
- Nihorimbere, V.; Ongena, M.; Smargiassi, M.; Thonart, P. Beneficial effect of the rhizosphere microbial community for plant growth and health. Biotechnol. Agron. Soc. Et Environ. 2011, 15, 327–337. [Google Scholar]
- Cottrell, M.T.; Kirchman, D.L. Contribution of major bacterial groups to bacterial biomass production (thymidine and leucine incorporation) in the Delaware estuary. Limnol. Oceanogr. 2003, 48, 168–178. [Google Scholar] [CrossRef]
- Singh, S.N. Microbial Degradation of Xenobiotics; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Bao, Y.Y.; Dolfing, J.; Wang, B.Z.; Chen, R.R.; Huang, M.S.; Li, Z.P.; Lin, X.G.; Feng, Y.Z. Bacterial communities involved directly or indirectly in the anaerobic degradation of cellulose. Biol. Fert. Soils 2019, 55, 201–211. [Google Scholar] [CrossRef]
- Bao, Y.Y.; Dolfing, J.; Guo, Z.Y.; Chen, R.R.; Wu, M.; Li, Z.P.; Lin, X.G.; Feng, Y.Z. Important ecophysiological roles of non-dominant Actinobacteria in plant residue decomposition, especially in less fertile soils. Microbiome 2021, 9, 84. [Google Scholar] [CrossRef]
- Qaswar, M.; Chai, R.S.; Ahmed, W.; Jing, H.; Han, T.F.; Liu, K.L.; Ye, X.X.; Xu, Y.M.; Anthonio, C.K.; Zhang, H.M. Partial substitution of chemical fertilizers with organic amendments increased rice yield by changing phosphorus fractions and improving phosphatase activities in fluvo-aquic soil. J. Soils Sediments 2020, 20, 1285–1296. [Google Scholar] [CrossRef]
- Tejada, M.; Gonzalez, J.L. Crushed cotton gin compost on soil biological properties and rice yield. Eur. J. Agron. 2006, 25, 22–29. [Google Scholar] [CrossRef]
- Zhang, X.F.; Zhu, A.N.; Xin, X.L.; Yang, W.L.; Zhang, J.B.; Ding, S.J. Tillage and residue management for long-term wheat-maize cropping in the North China Plain: I. Crop yield and integrated soil fertility index. Field Crop. Res. 2018, 221, 157–165. [Google Scholar] [CrossRef]
- Yao, T.X.; Zhang, W.T.; Gulaqa, A.; Cui, Y.F.; Zhou, Y.M.; Weng, W.N.; Wang, X.; Liu, Q.T.; Jin, F. Effects of peanut shell biochar on soil nutrients, soil enzyme activity, and rice yield in heavily saline-sodic paddy field. J. Soil Sci. Plant. Nutr. 2021, 21, 655–664. [Google Scholar] [CrossRef]
- Dai, X.L.; Zhou, W.; Liu, G.R.; Liang, G.Q.; He, P.; Liu, Z.B. Soil C/N and pH together as a comprehensive indicator for evaluating the effects of organic substitution management in subtropical paddy fields after application of high-quality amendments. Geoderma 2019, 337, 1116–1125. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, C.; Bao, Y.; Petropoulos, E.; Wang, Y.; Zhong, Z.; Jiang, Y.; Ye, X.; Lin, X.; Feng, Y. Organic and Inorganic Amendments Shape Bacterial Indicator Communities That Can, In Turn, Promote Rice Yield. Microorganisms 2022, 10, 482. https://doi.org/10.3390/microorganisms10020482
Qiu C, Bao Y, Petropoulos E, Wang Y, Zhong Z, Jiang Y, Ye X, Lin X, Feng Y. Organic and Inorganic Amendments Shape Bacterial Indicator Communities That Can, In Turn, Promote Rice Yield. Microorganisms. 2022; 10(2):482. https://doi.org/10.3390/microorganisms10020482
Chicago/Turabian StyleQiu, Chongwen, Yuanyuan Bao, Evangelos Petropoulos, Yiming Wang, Zhenfang Zhong, Yaozhi Jiang, Xuhong Ye, Xiangui Lin, and Youzhi Feng. 2022. "Organic and Inorganic Amendments Shape Bacterial Indicator Communities That Can, In Turn, Promote Rice Yield" Microorganisms 10, no. 2: 482. https://doi.org/10.3390/microorganisms10020482
APA StyleQiu, C., Bao, Y., Petropoulos, E., Wang, Y., Zhong, Z., Jiang, Y., Ye, X., Lin, X., & Feng, Y. (2022). Organic and Inorganic Amendments Shape Bacterial Indicator Communities That Can, In Turn, Promote Rice Yield. Microorganisms, 10(2), 482. https://doi.org/10.3390/microorganisms10020482