
Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020

 , , , , , , , , , , , , , add
Show full author list
, , , , , , , , , , , , , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
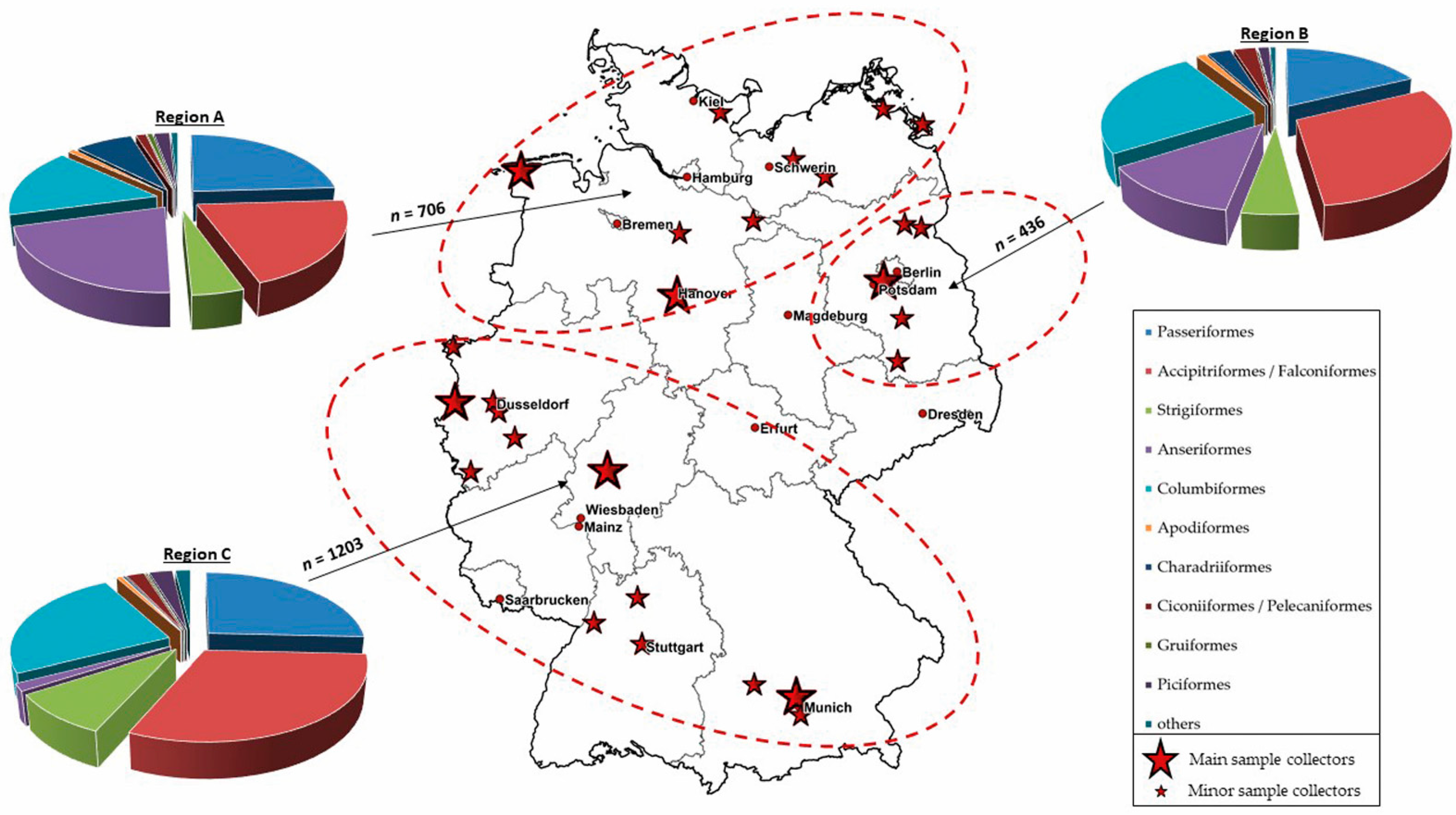
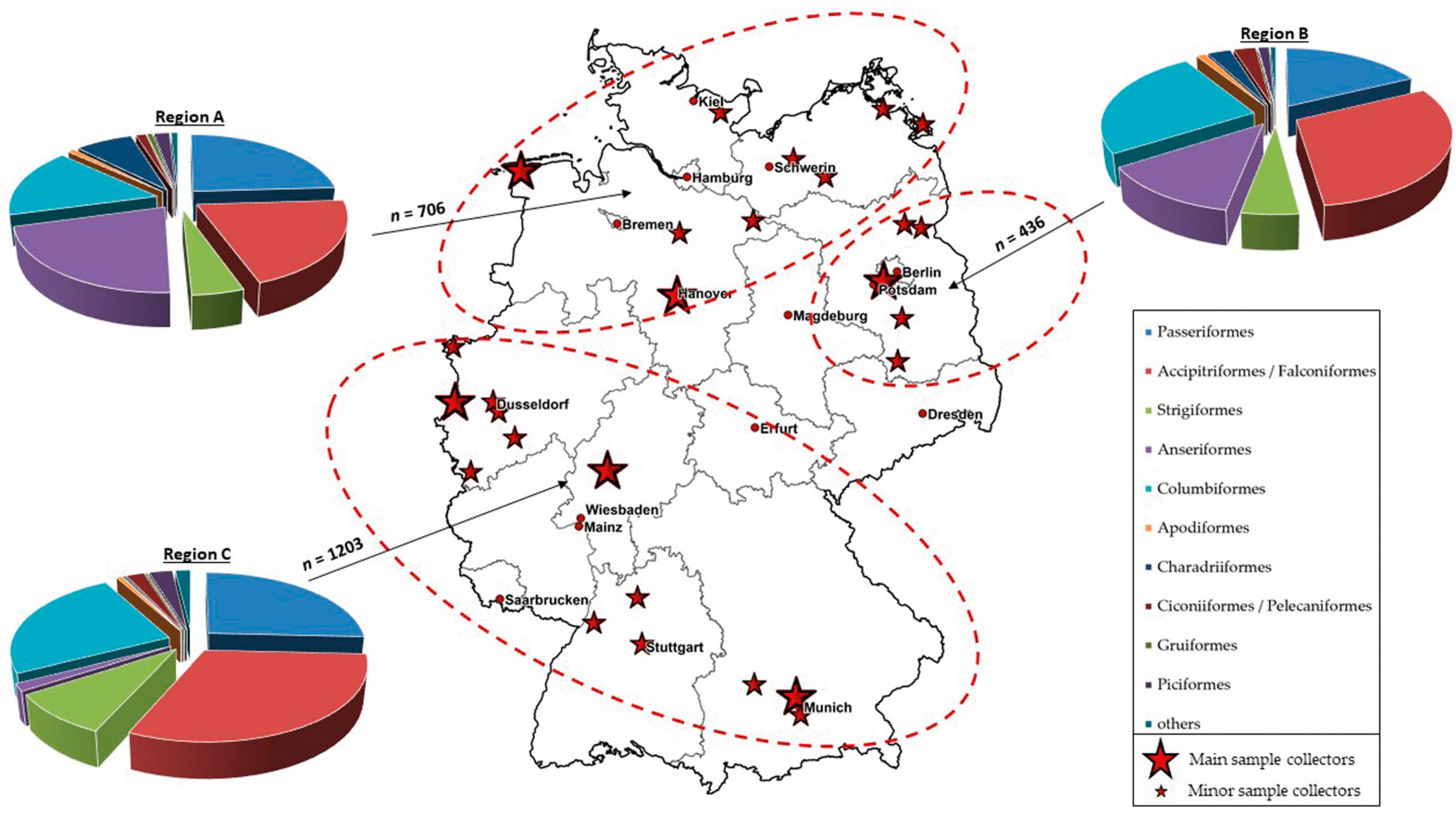
2.1. Sample Collection
2.2. Molecular Investigations
2.3. Sequencing and Phylogenetic Analysis
2.4. Serological Investigations
2.5. Statistical Analyses
2.6. Maps
2.7. Ethical Statement
3. Results
3.1. Results of the RT-qPCR
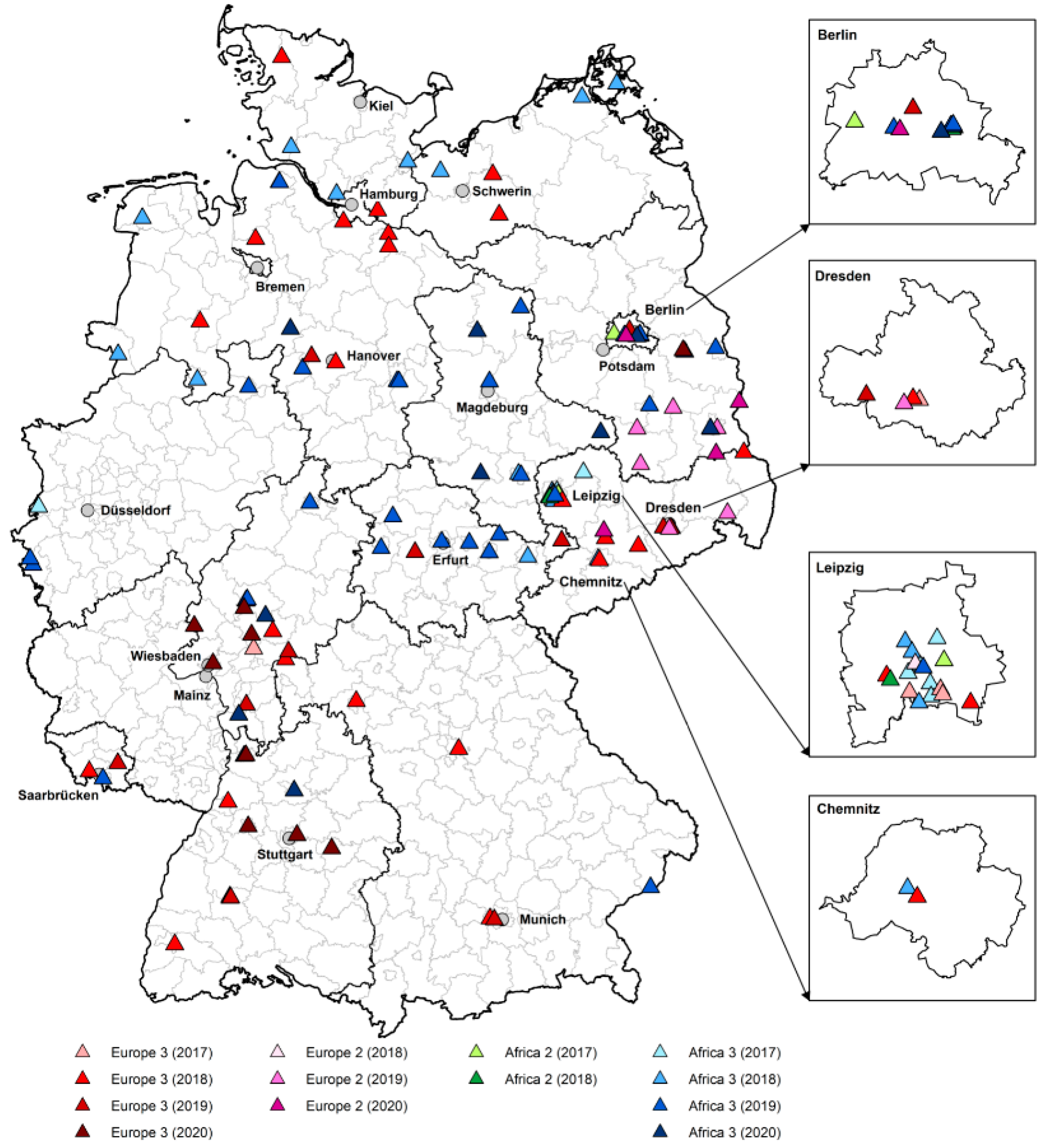
3.2. Phylogenetic Analysis of USUV RNA Positive Birds
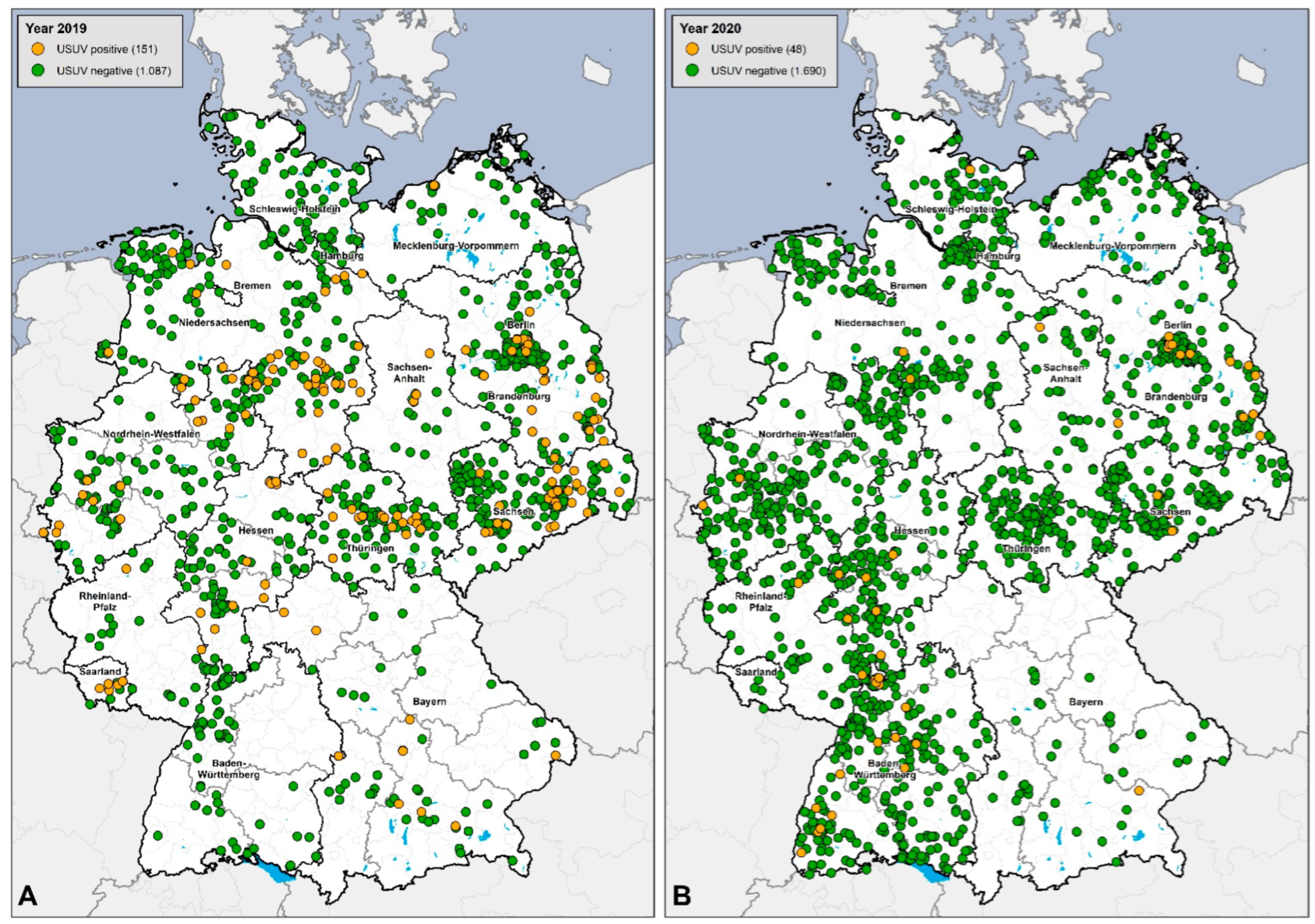
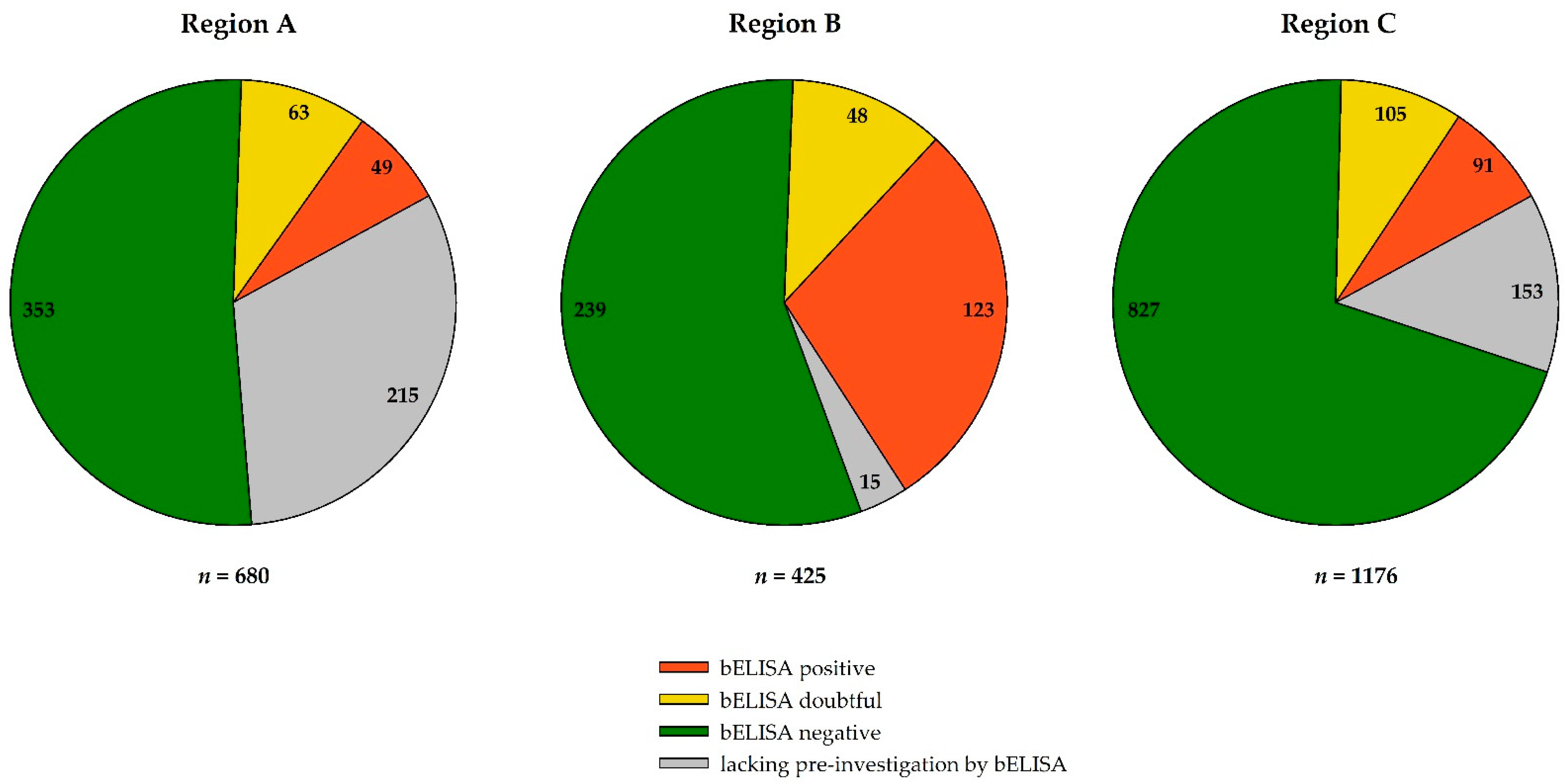
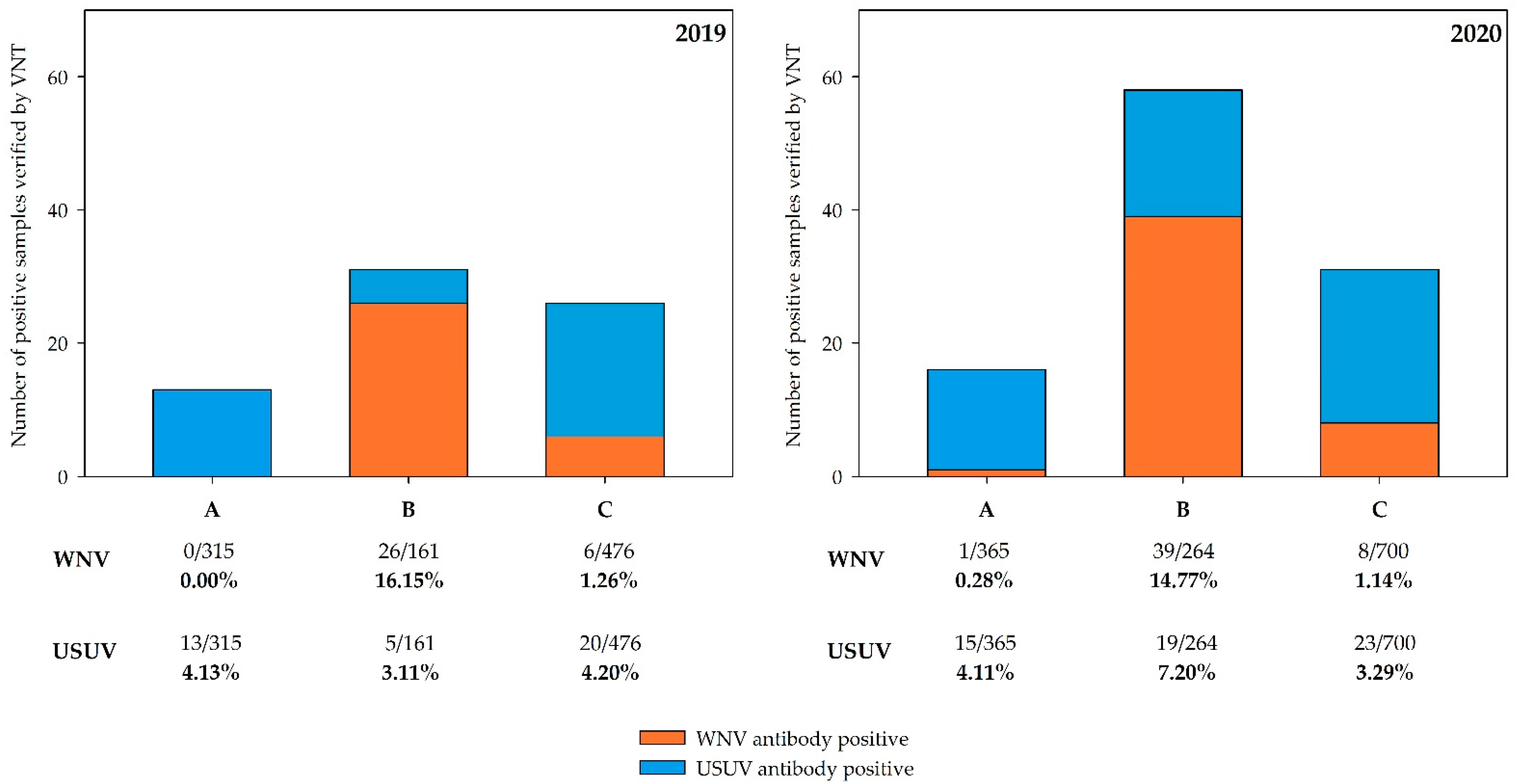
3.3. Serological Results
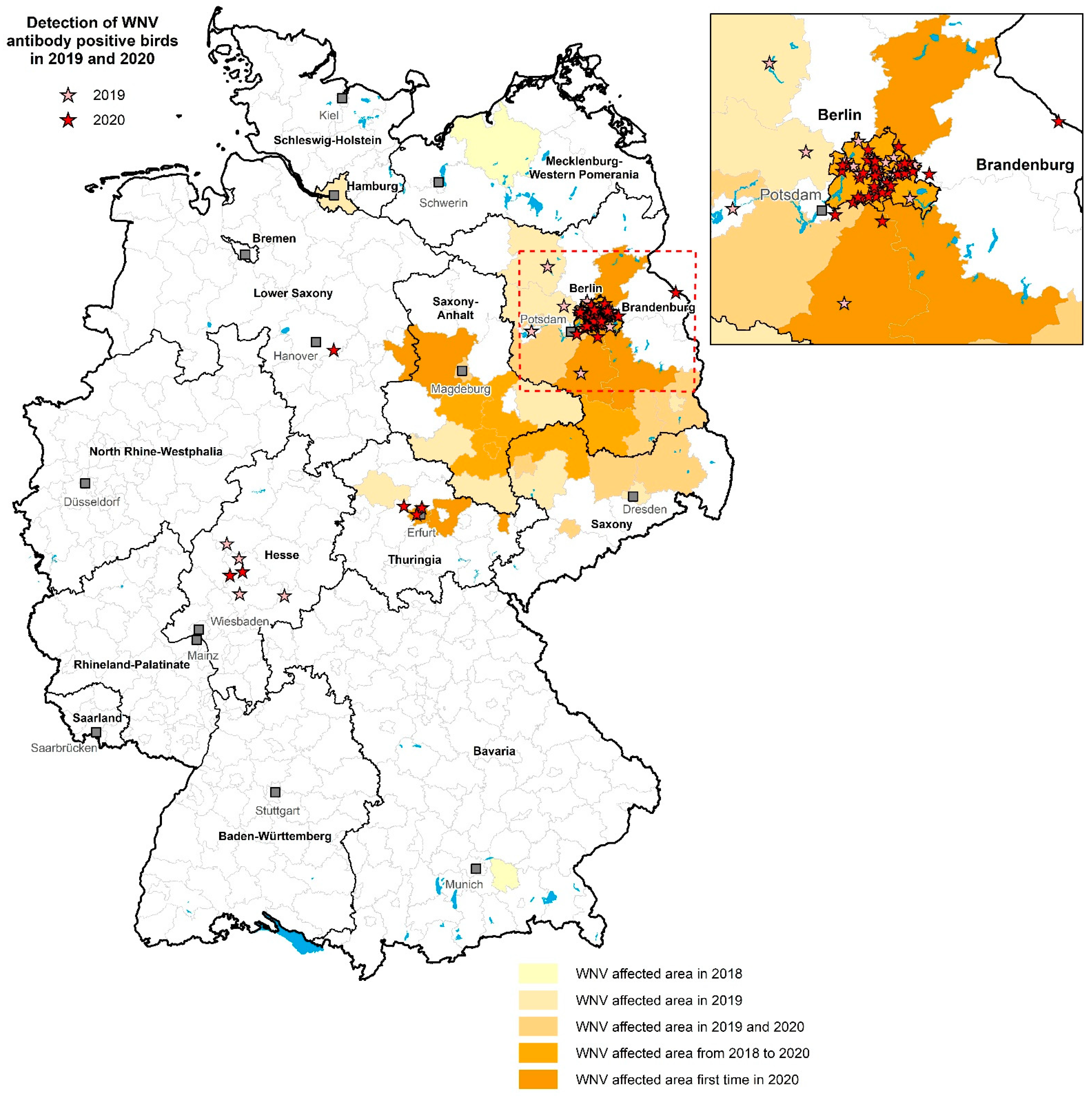
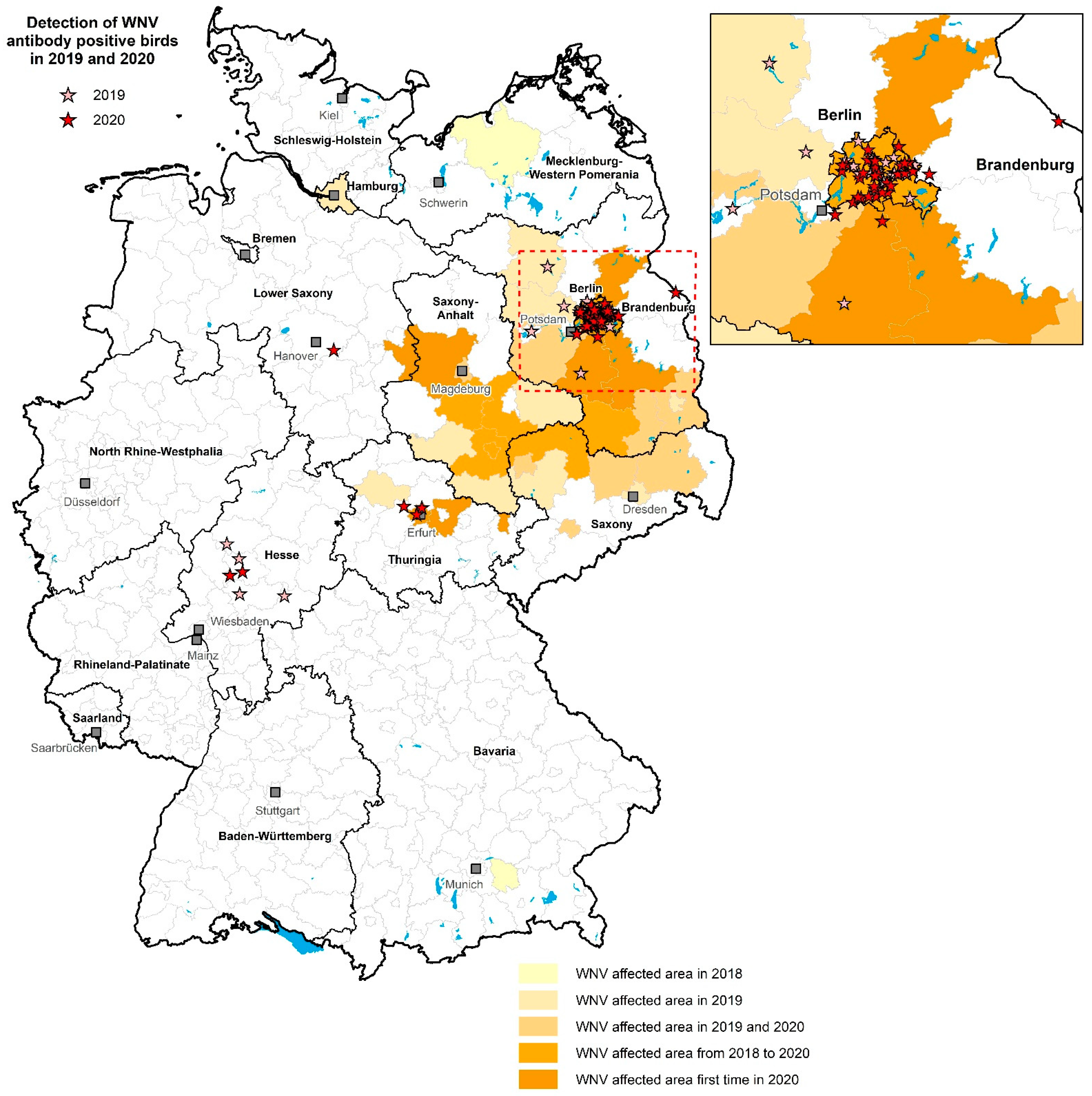
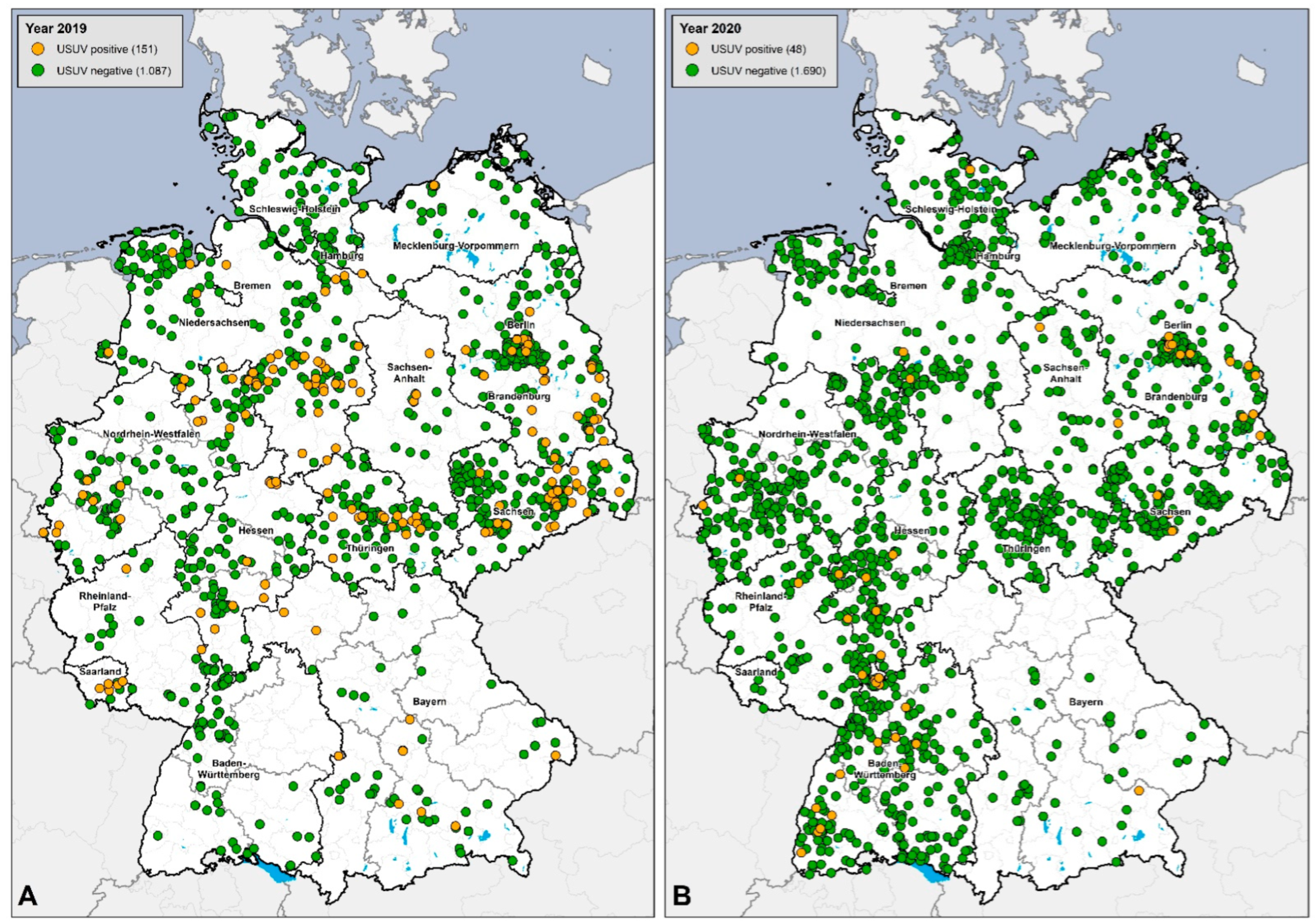
3.4. Regional Distribution of WNV- and USUV-Antibody-Positive Birds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hubálek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [CrossRef]
- Vilibic-Cavlek, T.; Savic, V.; Petrovic, T.; Toplak, I.; Barbic, L.; Petric, D.; Tabain, I.; Hrnjakovic-Cvjetkovic, I.; Bogdanic, M.; Klobucar, A.; et al. Emerging Trends in the Epidemiology of West Nile and Usutu Virus Infections in Southern Europe. Front. Vet. Sci. 2019, 6, 437. [Google Scholar] [CrossRef] [Green Version]
- Zannoli, S.; Sambri, V. West Nile Virus and Usutu Virus Co-Circulation in Europe: Epidemiology and Implications. Microorganisms 2019, 7, 184. [Google Scholar] [CrossRef] [Green Version]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [Green Version]
- Roesch, F.; Fajardo, A.; Moratorio, G.; Vignuzzi, M. Usutu Virus: An Arbovirus on the Rise. Viruses 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, S.; Kim, B.-S.; Chipman, P.R.; Rossmann, M.G.; Kuhn, R.J. Structure of West Nile virus. Science 2003, 302, 248. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, P.; Becher, P.; Bukh, J.; Gould, E.A.; Meyers, G.; Monath, T.; Muerhoff, S.; Pletnev, A.; Rico-Hesse, R.; Smith, D.B.; et al. ICTV Virus Taxonomy Profile: Flaviviridae. J. Gen. Virol. 2017, 98, 2–3. [Google Scholar] [CrossRef]
- Fall, G.; Di Paola, N.; Faye, M.; Dia, M.; Freire, C.C.D.M.; Loucoubar, C.; Zanotto, P.M.d.A.; Faye, O.; Sall, A.A. Biological and phylogenetic characteristics of West African lineages of West Nile virus. PLoS Negl. Trop. Dis. 2017, 11, e0006078. [Google Scholar] [CrossRef]
- Cadar, D.; Lühken, R.; van der Jeugd, H.; Garigliany, M.; Ziegler, U.; Keller, M.; Lahoreau, J.; Lachmann, L.; Becker, N.; Kik, M.; et al. Widespread activity of multiple lineages of Usutu virus, western Europe, 2016. Euro Surveill. 2017, 22, 30452. [Google Scholar] [CrossRef]
- Weissenböck, H.; Bakonyi, T.; Rossi, G.; Mani, P.; Nowotny, N. Usutu virus, Italy, 1996. Emerg. Infect. Dis. 2013, 19, 274–277. [Google Scholar] [CrossRef]
- Weissenböck, H.; Kolodziejek, J.; Fragner, K.; Kuhn, R.; Pfeffer, M.; Nowotny, N. Usutu virus activity in Austria, 2001–2002. Microbes Infect. 2003, 5, 1132–1136. [Google Scholar] [CrossRef]
- Bakonyi, T.; Erdélyi, K.; Ursu, K.; Ferenczi, E.; Csörgo, T.; Lussy, H.; Chvala, S.; Bukovsky, C.; Meister, T.; Weissenböck, H.; et al. Emergence of Usutu virus in Hungary. J. Clin. Microbiol. 2007, 45, 3870–3874. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, H.W.; Bakonyi, T.; Weissenböck, H.; Hatt, J.-M.; Eulenberger, U.; Robert, N.; Hoop, R.; Nowotny, N. Emergence and establishment of Usutu virus infection in wild and captive avian species in and around Zurich, Switzerland—Genomic and pathologic comparison to other central European outbreaks. Vet. Microbiol. 2011, 148, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Manarolla, G.; Bakonyi, T.; Gallazzi, D.; Crosta, L.; Weissenböck, H.; Dorrestein, G.M.; Nowotny, N. Usutu virus in wild birds in northern Italy. Vet. Microbiol. 2010, 141, 159–163. [Google Scholar] [CrossRef] [Green Version]
- Jöst, H.; Bialonski, A.; Maus, D.; Sambri, V.; Eiden, M.; Groschup, M.H.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation of usutu virus in Germany. Am. J. Trop. Med. Hyg. 2011, 85, 551–553. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Jöst, H.; Ziegler, U.; Eiden, M.; Höper, D.; Emmerich, P.; Fichet-Calvet, E.; Ehichioya, D.U.; Czajka, C.; Gabriel, M.; et al. Epizootic emergence of Usutu virus in wild and captive birds in Germany. PLoS ONE 2012, 7, e32604. [Google Scholar] [CrossRef]
- Ziegler, U.; Jöst, H.; Müller, K.; Fischer, D.; Rinder, M.; Tietze, D.T.; Danner, K.-J.; Becker, N.; Skuballa, J.; Hamann, H.-P.; et al. Epidemic Spread of Usutu Virus in Southwest Germany in 2011 to 2013 and Monitoring of Wild Birds for Usutu and West Nile Viruses. Vector Borne Zoonotic Dis. 2015, 15, 481–488. [Google Scholar] [CrossRef]
- Cadar, D.; Bosch, S.; Jöst, H.; Börstler, J.; Garigliany, M.-M.; Becker, N.; Schmidt-Chanasit, J. Putative Lineage of Novel African Usutu Virus, Central Europe. Emerg. Infect. Dis. 2015, 21, 1647–1650. [Google Scholar] [CrossRef]
- Ziegler, U.; Fast, C.; Eiden, M.; Bock, S.; Schulze, C.; Hoeper, D.; Ochs, A.; Schlieben, P.; Keller, M.; Zielke, D.E.; et al. Evidence for an independent third Usutu virus introduction into Germany. Vet. Microbiol. 2016, 192, 60–66. [Google Scholar] [CrossRef]
- Sieg, M.; Schmidt, V.; Ziegler, U.; Keller, M.; Höper, D.; Heenemann, K.; Rückner, A.; Nieper, H.; Muluneh, A.; Groschup, M.H.; et al. Outbreak and Cocirculation of Three Different Usutu Virus Strains in Eastern Germany. Vector Borne Zoonotic Dis. 2017, 17, 662–664. [Google Scholar] [CrossRef]
- Michel, F.; Fischer, D.; Eiden, M.; Fast, C.; Reuschel, M.; Müller, K.; Rinder, M.; Urbaniak, S.; Brandes, F.; Schwehn, R.; et al. West Nile Virus and Usutu Virus Monitoring of Wild Birds in Germany. Int. J. Environ. Res. Public Health 2018, 15, 171. [Google Scholar] [CrossRef] [Green Version]
- Michel, F.; Sieg, M.; Fischer, D.; Keller, M.; Eiden, M.; Reuschel, M.; Schmidt, V.; Schwehn, R.; Rinder, M.; Urbaniak, S.; et al. Evidence for West Nile Virus and Usutu Virus Infections in Wild and Resident Birds in Germany, 2017 and 2018. Viruses 2019, 11, 674. [Google Scholar] [CrossRef] [Green Version]
- Clé, M.; Beck, C.; Salinas, S.; Lecollinet, S.; Gutierrez, S.; van de Perre, P.; Baldet, T.; Foulongne, V.; Simonin, Y. Usutu virus: A new threat? Epidemiol. Infect. 2019, 147, e232. [Google Scholar] [CrossRef] [Green Version]
- Oude Munnink, B.B.; Münger, E.; Nieuwenhuijse, D.F.; Kohl, R.; van der Linden, A.; Schapendonk, C.M.E.; van der Jeugd, H.; Kik, M.; Rijks, J.M.; Reusken, C.B.E.M.; et al. Genomic monitoring to understand the emergence and spread of Usutu virus in The Netherlands, 2016–2018. Sci. Rep. 2020, 10, 2798. [Google Scholar] [CrossRef] [Green Version]
- Vilibic-Cavlek, T.; Petrovic, T.; Savic, V.; Barbic, L.; Tabain, I.; Stevanovic, V.; Klobucar, A.; Mrzljak, A.; Ilic, M.; Bogdanic, M.; et al. Epidemiology of Usutu Virus: The European Scenario. Pathogens 2020, 9, 699. [Google Scholar] [CrossRef]
- Weidinger, P.; Kolodziejek, J.; Bakonyi, T.; Brunthaler, R.; Erdélyi, K.; Weissenböck, H.; Nowotny, N. Different dynamics of Usutu virus infections in Austria and Hungary, 2017–2018. Transbound. Emerg. Dis. 2020, 67, 298–307. [Google Scholar] [CrossRef]
- Lühken, R.; Jöst, H.; Cadar, D.; Thomas, S.M.; Bosch, S.; Tannich, E.; Becker, N.; Ziegler, U.; Lachmann, L.; Schmidt-Chanasit, J. Distribution of Usutu Virus in Germany and Its Effect on Breeding Bird Populations. Emerg. Infect. Dis. 2017, 23, 1994–2001. [Google Scholar] [CrossRef] [Green Version]
- Seidowski, D.; Ziegler, U.; von Rönn, J.A.C.; Müller, K.; Hüppop, K.; Müller, T.; Freuling, C.; Mühle, R.-U.; Nowotny, N.; Ulrich, R.G.; et al. West Nile virus monitoring of migratory and resident birds in Germany. Vector Borne Zoonotic Dis. 2010, 10, 639–647. [Google Scholar] [CrossRef]
- Ziegler, U.; Seidowski, D.; Globig, A.; Fereidouni, S.R.; Ulrich, R.G.; Groschup, M.H. Sentinel birds in wild-bird resting sites as potential indicators for West Nile virus infections in Germany. Arch. Virol. 2010, 155, 965–969. [Google Scholar] [CrossRef]
- Ziegler, U.; Seidowski, D.; Angenvoort, J.; Eiden, M.; Müller, K.; Nowotny, N.; Groschup, M.H. Monitoring of West Nile virus infections in Germany. Zoonoses Public Health 2012, 59 (Suppl. 2), 95–101. [Google Scholar] [CrossRef]
- Eiden, M.; Ziegler, U.; Keller, M.; Müller, K.; Granzow, H.; Jöst, H.; Schmidt-Chanasit, J.; Groschup, M.H. Isolation of sindbis virus from a hooded crow in Germany. Vector Borne Zoonotic Dis. 2014, 14, 220–222. [Google Scholar] [CrossRef]
- Ziegler, U.; Fischer, D.; Eiden, M.; Reuschel, M.; Rinder, M.; Müller, K.; Schwehn, R.; Schmidt, V.; Groschup, M.H.; Keller, M. Sindbis virus—A wild bird associated zoonotic arbovirus circulates in Germany. Vet. Microbiol. 2019, 239, 108453. [Google Scholar] [CrossRef]
- Camp, J.V.; Nowotny, N. The knowns and unknowns of West Nile virus in Europe: What did we learn from the 2018 outbreak? Expert Rev. Anti. Infect. Ther. 2020, 18, 145–154. [Google Scholar] [CrossRef]
- Ziegler, U.; Lühken, R.; Keller, M.; Cadar, D.; van der Grinten, E.; Michel, F.; Albrecht, K.; Eiden, M.; Rinder, M.; Lachmann, L.; et al. West Nile virus epizootic in Germany, 2018. Antiviral Res. 2019, 162, 39–43. [Google Scholar] [CrossRef]
- Ziegler, U.; Keller, M.; Michel, F.; Globig, A.; Denzin, N.; Eiden, M.; Fast, C.; Gethmann, J.; Bastian, M.; Groschup, M.H.; et al. Aktuelles aus dem FLI zur West-Nil-Virus-Situation. Amtstierärztlicher Dienst und Lebensmittelkontrolle 2019, 26, 78–81. [Google Scholar]
- FLI. Erster Fall von West-Nil-Virus Infektion 2019. Available online: https://www.fli.de/de/aktuelles/kurznachrichten/neues-einzelansicht/erster-fall-von-west-nil-virus-infektion-2019/ (accessed on 21 December 2021).
- FLI. Weitere Ausbreitung des West-Nil-Virus: Fünf Infektionen bei Pferden und Erhöhte Fallzahlen bei Vögeln Festgestellt. Available online: https://www.fli.de/de/presse/pressemitteilungen/presse-einzelansicht/weitere-ausbreitung-des-west-nil-virus-fuenf-infektionen-bei-pferden-und-erhoehte-fallzahlen-bei-voege/ (accessed on 21 December 2021).
- Ziegler, U.; Santos, P.D.; Groschup, M.H.; Hattendorf, C.; Eiden, M.; Höper, D.; Eisermann, P.; Keller, M.; Michel, F.; Klopfleisch, R.; et al. West Nile Virus Epidemic in Germany Triggered by Epizootic Emergence, 2019. Viruses 2020, 12, 448. [Google Scholar] [CrossRef] [Green Version]
- Kampen, H.; Holicki, C.M.; Ziegler, U.; Groschup, M.H.; Tews, B.A.; Werner, D. West Nile Virus Mosquito Vectors (Diptera: Culicidae) in Germany. Viruses 2020, 12, 493. [Google Scholar] [CrossRef]
- Frank, C.; Lachmann, R.; Stark, K.; Schmidt-Chanasit, J.; Eisermann, P.; Lühken, R. Autochthone Infektionen mit dem West-Nil-Virus in Deutschland 2018 und 2019. Epidemiol. Bull. 2020, 3–10. [Google Scholar] [CrossRef]
- FLI. Aktuelle Situation West-Nil-Virus. Available online: https://www.fli.de/de/aktuelles/kurznachrichten/neues-einzelansicht/aktuelle-situation-west-nil-virus/ (accessed on 21 December 2021).
- RKI. Auch 2021 Mückenübertragungen von West-Nil-Virus in Deutschland zu erwarten. Epidemiol. Bull. 2021, 40–41. [Google Scholar]
- Pietsch, C.; Michalski, D.; Münch, J.; Petros, S.; Bergs, S.; Trawinski, H.; Lübbert, C.; Liebert, U.G. Autochthonous West Nile virus infection outbreak in humans, Leipzig, Germany, August to September 2020. Euro Surveill. 2020, 25, 2001786. [Google Scholar] [CrossRef]
- Eiden, M.; Vina-Rodriguez, A.; Hoffmann, B.; Ziegler, U.; Groschup, M.H. Two new real-time quantitative reverse transcription polymerase chain reaction assays with unique target sites for the specific and sensitive detection of lineages 1 and 2 West Nile virus strains. J. Vet. Diagn. Invest. 2010, 22, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Cavrini, F.; Della Pepa, M.E.; Gaibani, P.; Pierro, A.M.; Rossini, G.; Landini, M.P.; Sambri, V. A rapid and specific real-time RT-PCR assay to identify Usutu virus in human plasma, serum, and cerebrospinal fluid. J. Clin. Virol. 2011, 50, 221–223. [Google Scholar] [CrossRef]
- Eiden, M.; Gil, P.; Ziegler, U.; Rakotoarivony, I.; Marie, A.; Frances, B.; L’Ambert, G.; Simonin, Y.; Foulongne, V.; Groschup, M.H.; et al. Emergence of two Usutu virus lineages in Culex pipiens mosquitoes in the Camargue, France, 2015. Infect. Genet. Evol. 2018, 61, 151–154. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Xia, X. DAMBE7: New and Improved Tools for Data Analysis in Molecular Biology and Evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D. Phylogenetic analysis using parsimony (* and other methods). In Sinauer Associates, Version 4; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Rambaut, A. FigTree, Version 1.4.3. Computer Program Distributed by the Author. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 22 December 2021).
- Cadar, D.; Becker, N.; Campos, R.D.M.; Börstler, J.; Jöst, H.; Schmidt-Chanasit, J. Usutu virus in bats, Germany, 2013. Emerg. Infect. Dis. 2014, 20, 1771–1773. [Google Scholar] [CrossRef]
- Mayr, A.; Bachmann, P.A.; Bibrack, B.; Wittmann, G. Neutralisationstest. In Virologische Arbeitsmethoden: Band II (Serologie); Mayr, A., Bachmann, P.A., Bibrack, B., Wittmann, G., Eds.; Gustav Fischer Verlag: Jena, Germany, 1977; pp. 457–534. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.r-project.org/index.html (accessed on 12 February 2022).
- GeoBasis-DE. Available online: https://gdz.bkg.bund.de/index.php/default/open-data.html (accessed on 15 February 2022).
- Feyer, S.; Bartenschlager, F.; Bertram, C.A.; Ziegler, U.; Fast, C.; Klopfleisch, R.; Muller, K. Clinical, pathological and virological aspects of fatal West Nile virus infections in ten free-ranging goshawks (Accipiter gentilis) in Germany. In Transboundary and Emerging Diseases; Wiley: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Santos, P.D.; Michel, F.; Wylezich, C.; Höper, D.; Keller, M.; Holicki, C.M.; Szentiks, C.A.; Eiden, M.; Muluneh, A.; Neubauer-Juric, A.; et al. Co-infections: Simultaneous detections of West Nile virus and Usutu virus in birds from Germany. Transbound. Emerg. Dis. 2021. [Google Scholar] [CrossRef]
- Ziegler, U.; Santos, P.D.; Bergmann, F.; Günther, A.; Holicli, C.M.; Wylezich, C.; Höper, D.; Fast, C.; Keller, M.; Beer, M.; et al. Aktuelle West-Nil-Virus und Usutu-Virus Situation in Deutschland und erste Nachweise von Ko-Infektionen in Vögeln. In 39. Jahrestagung der DVG-Fachgruppe “AVID”, Schwerpunkt Virologie Online; Deutsche Veterinärmedizinische Gesellschaft e.V. (DVG): Gießen, Germany, 2021; pp. 57–58. [Google Scholar]
- Störk, T.; de Le Roi, M.; Haverkamp, A.-K.; Jesse, S.T.; Peters, M.; Fast, C.; Gregor, K.M.; Könenkamp, L.; Steffen, I.; Ludlow, M.; et al. Analysis of avian Usutu virus infections in Germany from 2011 to 2018 with focus on dsRNA detection to demonstrate viral infections. Sci. Rep. 2021, 11, 24191. [Google Scholar] [CrossRef]
- Hubálek, Z.; Rudolf, I.; Čapek, M.; Bakonyi, T.; Betášová, L.; Nowotny, N. Usutu virus in blackbirds (Turdus merula), Czech Republic, 2011–2012. Transbound. Emerg. Dis. 2014, 61, 273–276. [Google Scholar] [CrossRef]
- Hubálek, Z.; Wegner, E.; Halouzka, J.; Tryjanowski, P.; Jerzak, L.; Sikutová, S.; Rudolf, I.; Kruszewicz, A.G.; Jaworski, Z.; Wlodarczyk, R. Serologic survey of potential vertebrate hosts for West Nile virus in Poland. Viral Immunol. 2008, 21, 247–253. [Google Scholar] [CrossRef]
- Bażanów, B.; van Jansen Vuren, P.; Szymański, P.; Stygar, D.; Frącka, A.; Twardoń, J.; Kozdrowski, R.; Pawęska, J.T. A Survey on West Nile and Usutu Viruses in Horses and Birds in Poland. Viruses 2018, 10, 87. [Google Scholar] [CrossRef] [Green Version]
- Hönig, V.; Palus, M.; Kaspar, T.; Zemanova, M.; Majerova, K.; Hofmannova, L.; Papezik, P.; Sikutova, S.; Rettich, F.; Hubalek, Z.; et al. Multiple Lineages of Usutu Virus (Flaviviridae, Flavivirus) in Blackbirds (Turdus merula) and Mosquitoes (Culex pipiens, Cx. modestus) in the Czech Republic (2016–2019). Microorganisms 2019, 7, 568. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Erdélyi, K.; Brunthaler, R.; Dán, Á.; Weissenböck, H.; Nowotny, N. Usutu virus, Austria and Hungary, 2010–2016. Emerg. Microbes Infect. 2017, 6, e85. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Chiapponi, C.; Bonilauri, P.; Lelli, D.; Baioni, L.; Barbieri, I.; Lavazza, A.; Pongolini, S.; Dottori, M.; Moreno, A. Co-circulation of two Usutu virus strains in Northern Italy between 2009 and 2014. Infect. Genet. Evol. 2017, 51, 255–262. [Google Scholar] [CrossRef]
- Scheuch, D.E.; Schäfer, M.; Eiden, M.; Heym, E.C.; Ziegler, U.; Walther, D.; Schmidt-Chanasit, J.; Keller, M.; Groschup, M.H.; Kampen, H. Detection of Usutu, Sindbis, and Batai Viruses in Mosquitoes (Diptera: Culicidae) Collected in Germany, 2011–2016. Viruses 2018, 10, 389. [Google Scholar] [CrossRef] [Green Version]
- Kampen, H.; Tews, B.A.; Werner, D. First Evidence of West Nile Virus Overwintering in Mosquitoes in Germany. Viruses 2021, 13, 2463. [Google Scholar] [CrossRef]
- Holicki, C.M.; Ziegler, U.; Raileanu, C.; Kampen, H.; Werner, D.; Schulz, J.; Silaghi, C.; Groschup, M.H.; Vasic, A. West Nile Virus Lineage 2 Vector Competence of Indigenous Culex and Aedes Mosquitoes from Germany at Temperate Climate Conditions. Viruses 2020, 12, 561. [Google Scholar] [CrossRef]
- Holicki, C.M.; Scheuch, D.E.; Ziegler, U.; Lettow, J.; Kampen, H.; Werner, D.; Groschup, M.H. German Culex pipiens biotype molestus and Culex torrentium are vector-competent for Usutu virus. Parasit. Vectors 2020, 13, 625. [Google Scholar] [CrossRef]
- Erdélyi, K.; Ursu, K.; Ferenczi, E.; Szeredi, L.; Rátz, F.; Skáre, J.; Bakonyi, T. Clinical and pathologic features of lineage 2 West Nile virus infections in birds of prey in Hungary. Vector Borne Zoonotic Dis. 2007, 7, 181–188. [Google Scholar] [CrossRef]
- Jiménez-Clavero, M.A.; Sotelo, E.; Fernandez-Pinero, J.; Llorente, F.; Blanco, J.M.; Rodriguez-Ramos, J.; Perez-Ramirez, E.; Höfle, U. West Nile virus in golden eagles, Spain, 2007. Emerg. Infect. Dis. 2008, 14, 1489–1491. [Google Scholar] [CrossRef] [Green Version]
- Höfle, U.; Blanco, J.M.; Crespo, E.; Naranjo, V.; Jiménez-Clavero, M.A.; Sanchez, A.; de La Fuente, J.; Gortazar, C. West Nile virus in the endangered Spanish imperial eagle. Vet. Microbiol. 2008, 129, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Bakonyi, T.; Ferenczi, E.; Erdélyi, K.; Kutasi, O.; Csörgő, T.; Seidel, B.; Weissenböck, H.; Brugger, K.; Bán, E.; Nowotny, N. Explosive spread of a neuroinvasive lineage 2 West Nile virus in Central Europe, 2008/2009. Vet. Microbiol. 2013, 165, 61–70. [Google Scholar] [CrossRef]
- Csank, T.; Bhide, K.; Bencúrová, E.; Dolinská, S.; Drzewnioková, P.; Major, P.; Korytár, Ľ.; Bocková, E.; Bhide, M.; Pistl, J. Detection of West Nile virus and tick-borne encephalitis virus in birds in Slovakia, using a universal primer set. Arch. Virol. 2016, 161, 1679–1683. [Google Scholar] [CrossRef] [Green Version]
- Busquets, N.; Laranjo-González, M.; Soler, M.; Nicolás, O.; Rivas, R.; Talavera, S.; Villalba, R.; San Miguel, E.; Torner, N.; Aranda, C.; et al. Detection of West Nile virus lineage 2 in North-Eastern Spain (Catalonia). Transbound. Emerg. Dis. 2019, 66, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Hubálek, Z.; Kosina, M.; Rudolf, I.; Mendel, J.; Straková, P.; Tomešek, M. Mortality of Goshawks (Accipiter gentilis) Due to West Nile Virus Lineage 2. Vector Borne Zoonotic Dis. 2018, 18, 624–627. [Google Scholar] [CrossRef]
- Hubálek, Z.; Tomešek, M.; Kosina, M.; Šikutová, S.; Straková, P.; Rudolf, I. West Nile virus outbreak in captive and wild raptors, Czech Republic, 2018. Zoonoses Public Health 2019, 66, 978–981. [Google Scholar] [CrossRef]
- Vidaña, B.; Busquets, N.; Napp, S.; Pérez-Ramírez, E.; Jiménez-Clavero, M.Á.; Johnson, N. The Role of Birds of Prey in West Nile Virus Epidemiology. Vaccines 2020, 8, 550. [Google Scholar] [CrossRef]
- Ziegler, U.; Angenvoort, J.; Fischer, D.; Fast, C.; Eiden, M.; Rodriguez, A.V.; Revilla-Fernández, S.; Nowotny, N.; de La Fuente, J.G.; Lierz, M.; et al. Pathogenesis of West Nile virus lineage 1 and 2 in experimentally infected large falcons. Vet. Microbiol. 2013, 161, 263–273. [Google Scholar] [CrossRef]
- Chaintoutis, S.C.; Dovas, C.I.; Papanastassopoulou, M.; Gewehr, S.; Danis, K.; Beck, C.; Lecollinet, S.; Antalis, V.; Kalaitzopoulou, S.; Panagiotopoulos, T.; et al. Evaluation of a West Nile virus surveillance and early warning system in Greece, based on domestic pigeons. Comp. Immunol. Microbiol. Infect. Dis. 2014, 37, 131–141. [Google Scholar] [CrossRef]
- Napp, S.; Llorente, F.; Beck, C.; Jose-Cunilleras, E.; Soler, M.; Pailler-García, L.; Amaral, R.; Aguilera-Sepúlveda, P.; Pifarré, M.; Molina-López, R.; et al. Widespread Circulation of Flaviviruses in Horses and Birds in Northeastern Spain (Catalonia) between 2010 and 2019. Viruses 2021, 13, 2404. [Google Scholar] [CrossRef] [PubMed]
- Meister, T.; Lussy, H.; Bakonyi, T.; Sikutová, S.; Rudolf, I.; Vogl, W.; Winkler, H.; Frey, H.; Hubálek, Z.; Nowotny, N.; et al. Serological evidence of continuing high Usutu virus (Flaviviridae) activity and establishment of herd immunity in wild birds in Austria. Vet. Microbiol. 2008, 127, 237–248. [Google Scholar] [CrossRef]
- Sotelo, E.; Llorente, F.; Rebollo, B.; Camuñas, A.; Venteo, A.; Gallardo, C.; Lubisi, A.; Rodríguez, M.J.; Sanz, A.J.; Figuerola, J.; et al. Development and evaluation of a new epitope-blocking ELISA for universal detection of antibodies to West Nile virus. J. Virol. Methods 2011, 174, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Llorente, F.; García-Irazábal, A.; Pérez-Ramírez, E.; Cano-Gómez, C.; Sarasa, M.; Vázquez, A.; Jiménez-Clavero, M.Á. Influence of flavivirus co-circulation in serological diagnostics and surveillance: A model of study using West Nile, Usutu and Bagaza viruses. Transbound. Emerg. Dis. 2019, 66, 2100–2106. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.; Lowenski, S.; Durand, B.; Bahuon, C.; Zientara, S.; Lecollinet, S. Improved reliability of serological tools for the diagnosis of West Nile fever in horses within Europe. PLoS Negl. Trop. Dis. 2017, 11, e0005936. [Google Scholar] [CrossRef]
- Dobler, G. Zoonotic tick-borne flaviviruses. Vet. Microbiol. 2010, 140, 221–228. [Google Scholar] [CrossRef]
- Agüero, M.; Fernández-Pinero, J.; Buitrago, D.; Sánchez, A.; Elizalde, M.; San Miguel, E.; Villalba, R.; Llorente, F.; Jiménez-Clavero, M.A. Bagaza virus in partridges and pheasants, Spain, 2010. Emerg. Infect. Dis. 2011, 17, 1498–1501. [Google Scholar] [CrossRef]
- Beck, C.; Jimenez-Clavero, M.A.; Leblond, A.; Durand, B.; Nowotny, N.; Leparc-Goffart, I.; Zientara, S.; Jourdain, E.; Lecollinet, S. Flaviviruses in Europe: Complex circulation patterns and their consequences for the diagnosis and control of West Nile disease. Int. J. Environ. Res. Public Health 2013, 10, 6049–6083. [Google Scholar] [CrossRef] [Green Version]
- Bergmann, F.; Trachsel, D.S.; Stoeckle, S.D.; Bernis Sierra, J.; Lübke, S.; Groschup, M.H.; Gehlen, H.; Ziegler, U. Seroepidemiological Survey of West Nile Virus Infections in Horses from Berlin/Brandenburg and North Rhine-Westphalia, Germany. Viruses 2022, 14, 243. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | 2019 | 2020 | Total |
|---|---|---|---|
| Passeriformes | 232 | 327 | 559 |
| Accipitriformes/Falconiformes | 303 | 348 | 651 |
| Strigiformes | 86 | 81 | 167 |
| Anseriformes | 56 | 169 | 225 |
| Columbiformes | 169 | 346 | 515 |
| Apodiformes | 14 | 5 | 19 |
| Charadriiformes | 50 | 19 | 69 |
| Ciconiiformes/Pelecaniformes | 23 | 24 | 47 |
| Gruiformes | 5 | 3 | 8 |
| Piciformes | 20 | 34 | 54 |
| Cuculiformes | 1 | 0 | 1 |
| Suliformes | 1 | 1 | 2 |
| Psittaciformes | 1 | 4 | 5 |
| Podicipediformes | 1 | 0 | 1 |
| Galliformes | 7 | 3 | 10 |
| Struthioniformes | 0 | 1 | 1 |
| Coraciiformes | 2 | 0 | 2 |
| Gaviiformes | 0 | 1 | 1 |
| Procellariiformes | 1 | 0 | 1 |
| Caprimulgiformes | 1 | 0 | 1 |
| unknown | 4 | 2 | 6 |
| Total | 977 | 1368 | 2345 |
| Year | Region | Order | Common Name | Scientific Name | No. Tested Birds | Migration Pattern | Housing | WNV RT-qPCR No. Pos. (%) | USUV RT-qPCR No. Pos. (%) |
|---|---|---|---|---|---|---|---|---|---|
| 2019 | B | Accipitriformes | Northern Goshawk | Accipiter gentilis | 18 | R, P | wild | 5 (27.8) | 0 (0) |
| Passeriformes | Eurasian Blackbird | Turdus merula | 5 | R, P | wild | 0 (0) | 1 (20) | ||
| Hooded Crow | Corvus cornix | 13 | R, (S) | wild | 1 (7.7) | 0 (0) | |||
| Pelecaniformes | Grey Heron | Ardea cinerea | 5 | R, P, S | wild | 1 (20) | 0 (0) | ||
| C | Accipitriformes | Common Buzzard | Buteo buteo | 34 | R, P, S | wild | 0 (0) | 3 (8.8) | |
| Harris’s Hawk | Parabuteo unicinctus | 14 | zoo bird | captive | 0 (0) | 1 (7.1) | |||
| Northern Goshawk | Accipiter gentilis | 8 | R, P | wild | 0 (0) | 1 (12.5) | |||
| Steppe Eagle | Aquila nipalensis | 4 | zoo bird | captive | 0 (0) | 1 (25) | |||
| European Honey Buzzard | Pernis apicorus | 3 | L | wild | 0 (0) | 1 (33.3) | |||
| Columbiformes | Common Wood Pigeon | Columba palumbus | 42 | R, P, S | wild | 0 (0) | 6 (14.3) | ||
| Passeriformes | Thrush | Turdus sp. | 4 | S, L | wild | 0 (0) | 1 (25) | ||
| Eurasian Blackbird | Turdus merula | 29 | R, P | wild | 0 (0) | 4 (13.8) | |||
| Carrion Crow | Corvus corone | 23 | R, (S) | wild | 0 (0) | 2 (8.7) | |||
| House Sparrow | Passer domesticus | 7 | R | wild | 0 (0) | 1 (14.3) | |||
| Pelecaniformes | Grey Heron | Ardea cinerea | 6 | R, P, S | wild | 0 (0) | 1 (16.7) | ||
| Strigiformes | Eurasian Tawny Owl | Strix aluco | 12 | R | wild | 0 (0) | 2 (16.7) | ||
| Great Grey Owl | Strix nebulosa | 2 | zoo bird | captive | 0 (0) | 1 (50) | |||
| In Total in 2019 | 964 | 7 (0.7) | 26 (2.7) | ||||||
| 2020 | A | Passeriformes | Eurasian Blackbird | Turdus merula | 29 | R, P | wild | 0 (0) | 1 (3.4) |
| B | Accipitriformes | Northern Goshawk | Accipiter gentilis | 31 | R, P | wild | 10 (32.3) | 1 (3.3) | |
| Columbiformes | Common Wood Pigeon | Columba palumbus | 60 | R, P, S | wild | 0 (0) | 3 (5) | ||
| Passeriformes | Hooded Crow | Corvus cornix | 32 | R, (S) | wild | 2 (6.3) | 0 (0) | ||
| Great Tit | Parus major | 1 | R, (P) | wild | 0 (0) | 1 (100) | |||
| C | Accipitriformes | Harris’s Hawk | Parabuteo unicinctus | 7 | zoo bird | captive | 0 (0) | 1 (14.3) | |
| Northern Goshawk | Accipiter gentilis | 10 | R, P | wild | 0 (0) | 1 (10) | |||
| Anseriformes | Mute Swan | Cyngus olor | 2 | R, P, S | wild | 0 (0) | 1 (50) | ||
| Columbiformes | Common Wood Pigeon | Columba palumbus | 165 | R, P, S | wild | 0 (0) | 2 (1.2) | ||
| Passeriformes | Eurasian Blackbird | Turdus merula | 37 | R, P | wild | 0 (0) | 5 (13.5) | ||
| In Total in 2020 | 1348 | 12 (0.9) | 16 (1.2) | ||||||
| Year | Order | Common Name | Scientific Name | Migration Pattern | Housing | USUV RNA Positive/Tested |
|---|---|---|---|---|---|---|
| 2019 | Eurasian Blackbird | Turdus merula | R, P | wild | 98/223 | |
| Eurasian Blue Tit | Parus caeruleus | R | wild | 1/6 | ||
| Thrush sp. | Turdus sp. | S, L | wild | 3/18 | ||
| True Finches sp. | Fringillidae | n.d. | wild | 2/14 | ||
| Passeriformes | European Greenfinch | Carduelis chloris | S | wild | 1/35 | |
| House Sparrow | Passer domesticus | R | wild | 1/13 | ||
| Great Tit | Parus major | R, (P) | wild | 2/6 | ||
| Corvis sp. | Corvus sp. | R, (P) | wild | 1/6 | ||
| Tits sp. | Parus sp. | R, (P) | wild | 2/6 | ||
| European Robin | Erithacus rubecula | R, P | wild | 1/3 | ||
| Song thrush | Turdus philomelos | R, S | wild | 5/12 | ||
| Common Starling | Sturnus vulgaris | R, P, S | wild | 4/32 | ||
| Coal Tit | Parus ater | R, S | wild | 2/3 | ||
| Great Grey Owl | Strix nebulosa | zoo | captive | 9/15 | ||
| Snowy Owl | Bubo scandiacus | zoo | captive | 3/17 | ||
| Strigiformes | Northern Hawk Owl | Surnia ulula | zoo | captive | 1/4 | |
| Eurasian Tawny Owl | Strix aluco | R | wild | 1/6 | ||
| Northern Long-eared Owl | Asio otus | R, P, S | wild | 1/2 | ||
| Owls sp. | n.d. | n.d. | captive | 3/8 | ||
| Galliformes | Eurasian Capercaillie | Tetrao urogallus | zoo | captive | 1/4 | |
| Charadriiformes | Black-tailed Gull | Larus crassirostris | zoo | captive | 2/8 | |
| Anseriformes | Red-breasted Goose | Branta ruficollis | zoo | captive | 1/2 | |
| Columbiformes | Pigeon sp. | Columba sp. | n.d. | wild | 1/70 | |
| Unknown | Unknown species | n.d. | n.d. | captive/wild | 5/101 | |
| In Total | 151/(1238) | |||||
| 2020 | Passeriformes | Eurasian Blackbird | Turdus merula | R, P | wild | 24/158 |
| Accipitriformes | Blue Tit | Parus caeruleus | R | wild | 5/156 | |
| Passeriformes | Common Chaffinch | Fringilla coelebs | R, P | wild | 2/33 | |
| Thrush sp. | Turdus sp. | S, L | wild | 4/22 | ||
| Accipitriformes | House Sparrow | Passer domesticus | R | wild | 1/22 | |
| Great Tit | Parus major | R, (P) | wild | 2/72 | ||
| Anseriformes | Tits sp. | Parus sp. | n.d. | wild | 1/20 | |
| European Robin | Erithacus rubecula | R, P | wild | 1/9 | ||
| Song Trush | Turdus philomelos | R, S | wild | 1/8 | ||
| Great Grey Owl | Strix nebulosa | zoo | captive | 2/6 | ||
| Mute Swan | Cygnus olor | R, P, S | wild | 1/13 | ||
| Penguin sp. | Spheniscus sp. | zoo | captive | 1/21 | ||
| Unknown species | n.d. | n.d. | captive/wild | 3/177 | ||
| In Total | 48/(1738) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziegler, U.; Bergmann, F.; Fischer, D.; Müller, K.; Holicki, C.M.; Sadeghi, B.; Sieg, M.; Keller, M.; Schwehn, R.; Reuschel, M.; et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms 2022, 10, 807. https://doi.org/10.3390/microorganisms10040807
Ziegler U, Bergmann F, Fischer D, Müller K, Holicki CM, Sadeghi B, Sieg M, Keller M, Schwehn R, Reuschel M, et al. Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms. 2022; 10(4):807. https://doi.org/10.3390/microorganisms10040807
Chicago/Turabian StyleZiegler, Ute, Felicitas Bergmann, Dominik Fischer, Kerstin Müller, Cora M. Holicki, Balal Sadeghi, Michael Sieg, Markus Keller, Rebekka Schwehn, Maximilian Reuschel, and et al. 2022. "Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020" Microorganisms 10, no. 4: 807. https://doi.org/10.3390/microorganisms10040807
APA StyleZiegler, U., Bergmann, F., Fischer, D., Müller, K., Holicki, C. M., Sadeghi, B., Sieg, M., Keller, M., Schwehn, R., Reuschel, M., Fischer, L., Krone, O., Rinder, M., Schütte, K., Schmidt, V., Eiden, M., Fast, C., Günther, A., Globig, A., ... Groschup, M. H. (2022). Spread of West Nile Virus and Usutu Virus in the German Bird Population, 2019–2020. Microorganisms, 10(4), 807. https://doi.org/10.3390/microorganisms10040807