
Isolation and Genetic Characterization of Streptococcus iniae Virulence Factors in Adriatic Sturgeon (Acipenser naccarii)

 ,
,  ,
,  ,
, 
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
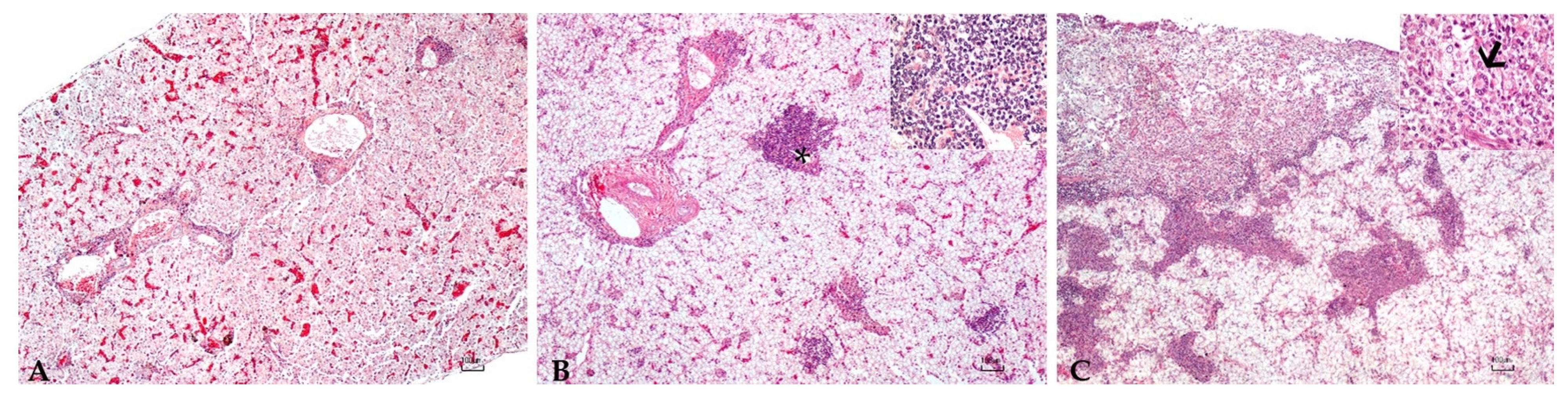
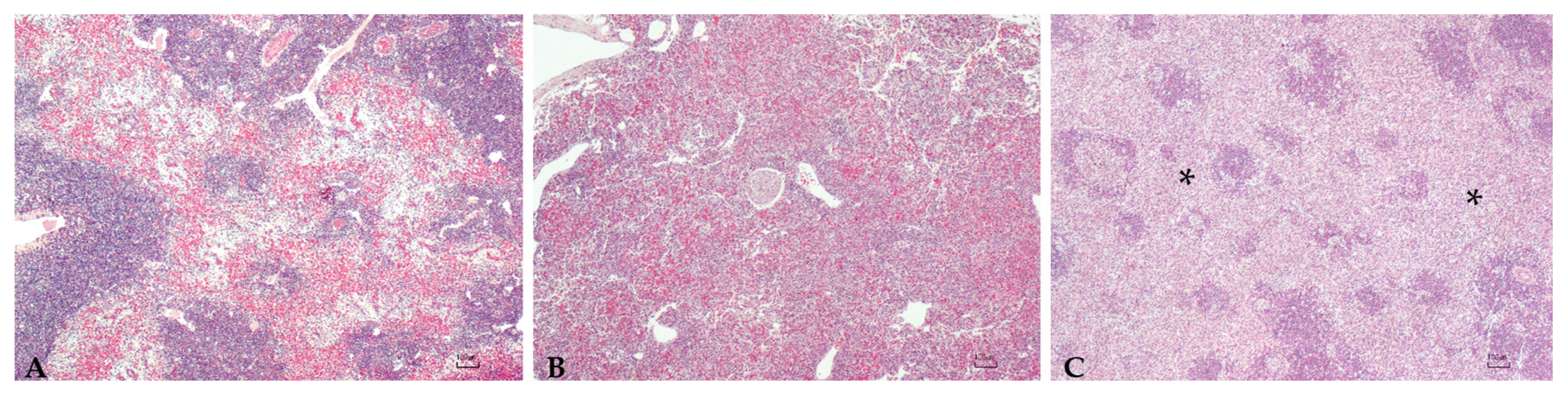
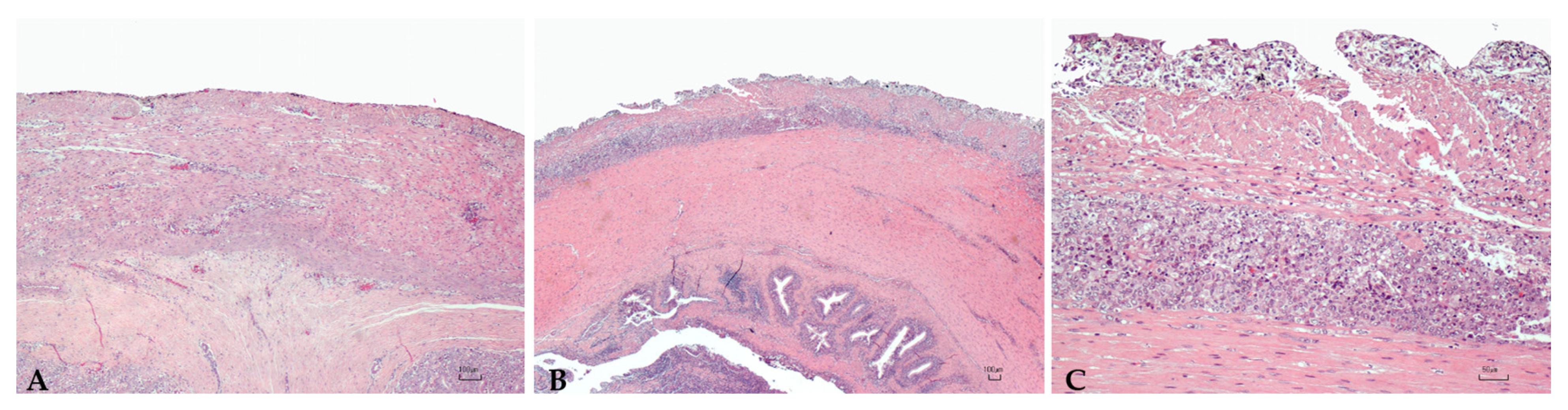
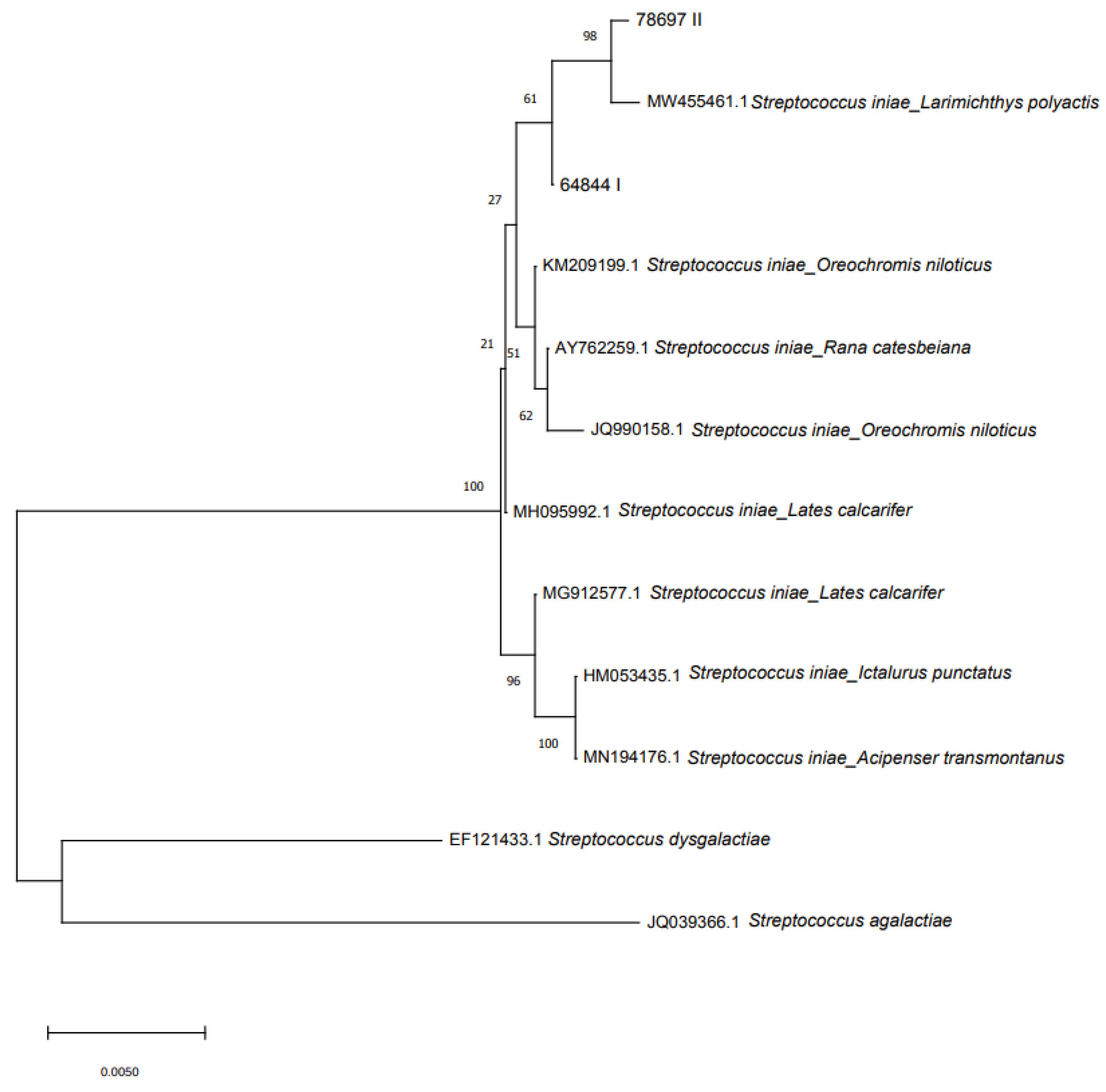
Abstract
Share and Cite
Colussi, S.; Pastorino, P.; Mugetti, D.; Antuofermo, E.; Sciuto, S.; Esposito, G.; Polinas, M.; Tomasoni, M.; Burrai, G.P.; Fernández-Garayzábal, J.F.; et al. Isolation and Genetic Characterization of Streptococcus iniae Virulence Factors in Adriatic Sturgeon (Acipenser naccarii). Microorganisms 2022, 10, 883. https://doi.org/10.3390/microorganisms10050883
Colussi S, Pastorino P, Mugetti D, Antuofermo E, Sciuto S, Esposito G, Polinas M, Tomasoni M, Burrai GP, Fernández-Garayzábal JF, et al. Isolation and Genetic Characterization of Streptococcus iniae Virulence Factors in Adriatic Sturgeon (Acipenser naccarii). Microorganisms. 2022; 10(5):883. https://doi.org/10.3390/microorganisms10050883
Chicago/Turabian StyleColussi, Silvia, Paolo Pastorino, Davide Mugetti, Elisabetta Antuofermo, Simona Sciuto, Giuseppe Esposito, Marta Polinas, Mattia Tomasoni, Giovanni Pietro Burrai, José Francisco Fernández-Garayzábal, and et al. 2022. "Isolation and Genetic Characterization of Streptococcus iniae Virulence Factors in Adriatic Sturgeon (Acipenser naccarii)" Microorganisms 10, no. 5: 883. https://doi.org/10.3390/microorganisms10050883
APA StyleColussi, S., Pastorino, P., Mugetti, D., Antuofermo, E., Sciuto, S., Esposito, G., Polinas, M., Tomasoni, M., Burrai, G. P., Fernández-Garayzábal, J. F., Acutis, P. L., Pedron, C., & Prearo, M. (2022). Isolation and Genetic Characterization of Streptococcus iniae Virulence Factors in Adriatic Sturgeon (Acipenser naccarii). Microorganisms, 10(5), 883. https://doi.org/10.3390/microorganisms10050883