
Phytoplankton Surveys in the Arctic Fram Strait Demonstrate the Tiny Eukaryotic Alga Micromonas and Other Picoprasinophytes Contribute to Deep Sea Export

 , , ,
, , , 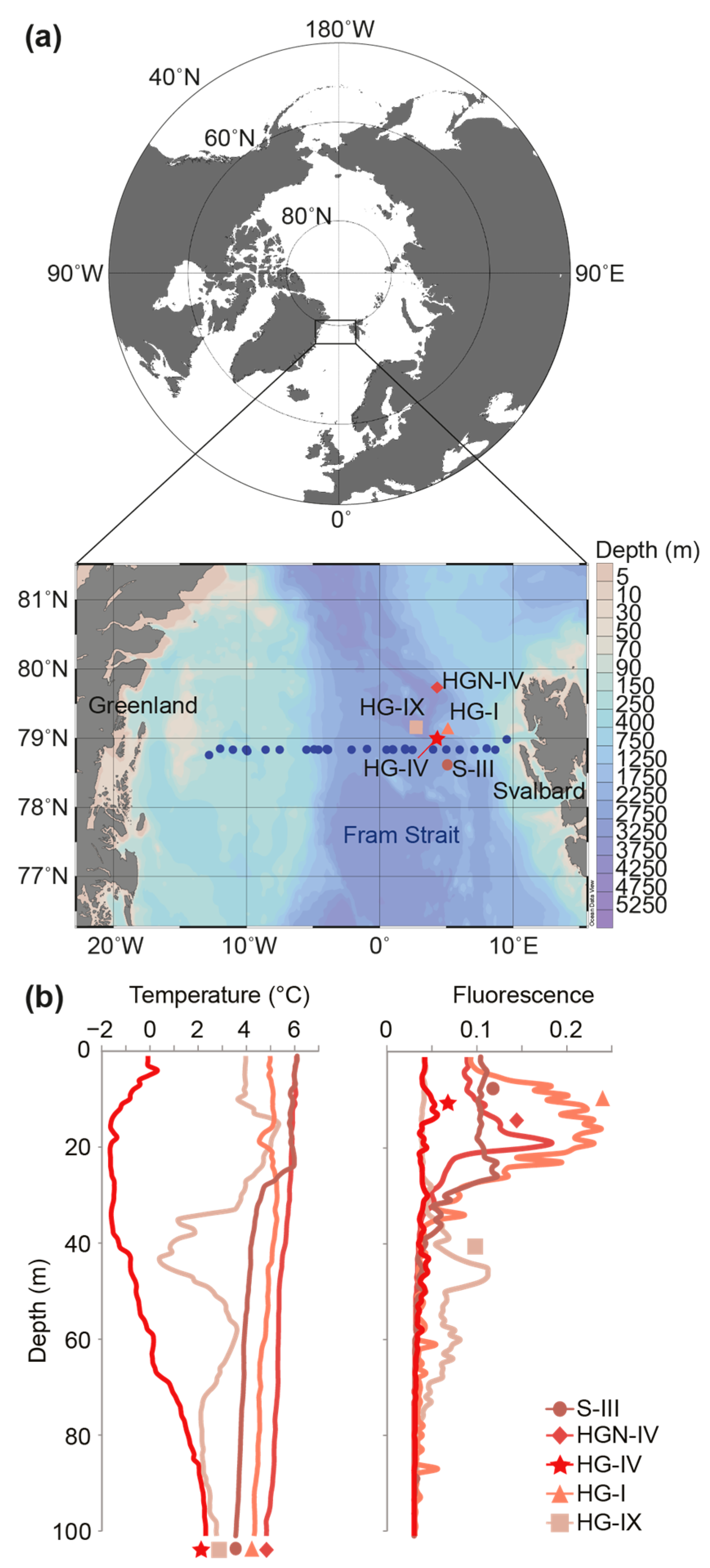{kind=link}
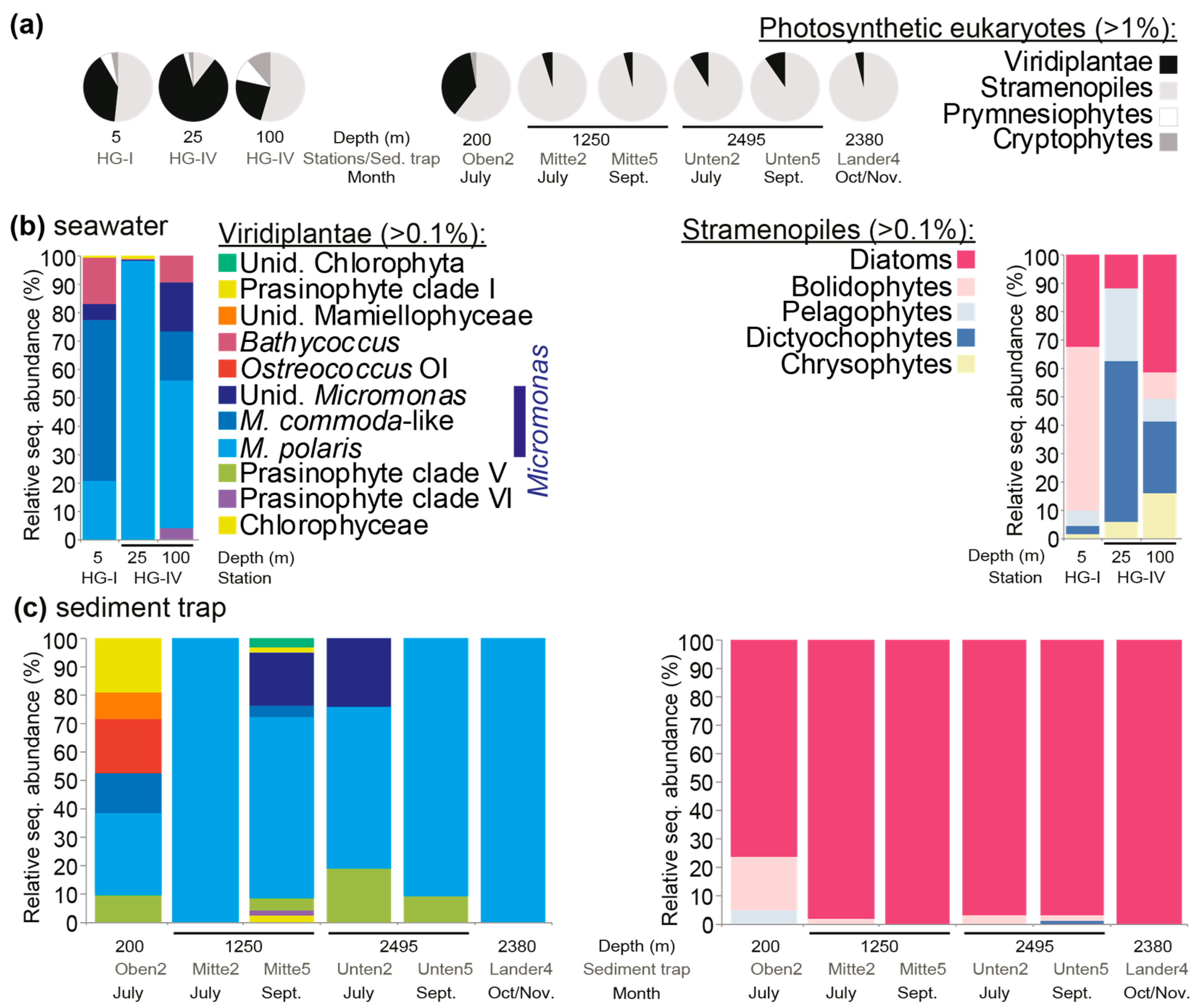{kind=link}
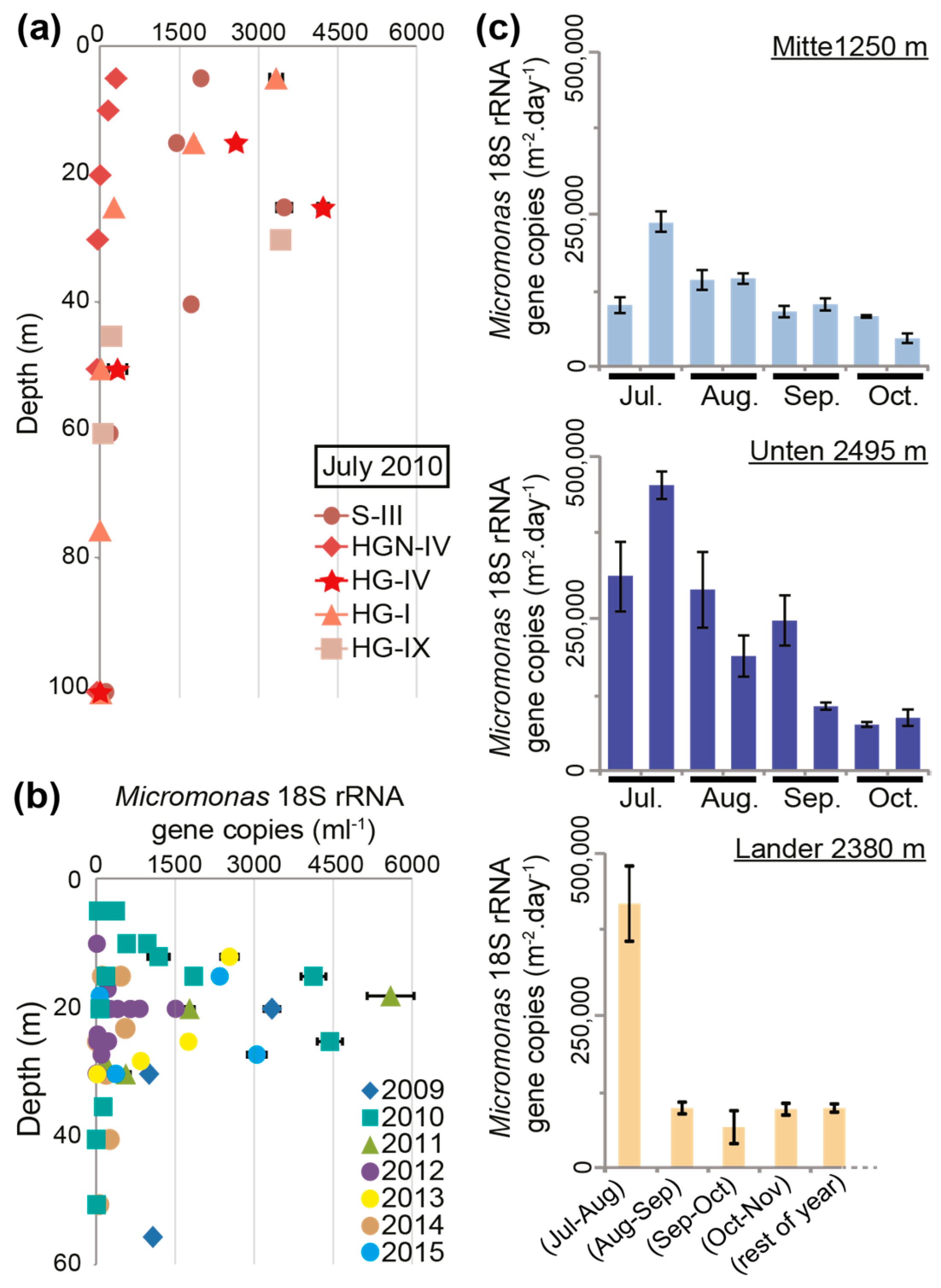{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Isolation
2.3. High-Throughput Sequencing and Analysis
2.4. Statistical Analyses
2.5. Quantitative PCR
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sieburth, J.M.; Smetacek, V.; Lenz, J. Pelagic Ecosystem Structure: Heterotrophic Compartments of the Plankton and Their Relationship to Plankton Size Fractions 1. Limnol. Oceanogr. 1978, 23, 1256–1263. [Google Scholar] [CrossRef]
- Bolaños, L.M.; Karp-Boss, L.; Choi, C.J.; Worden, A.Z.; Graff, J.R.; Haëntjens, N.; Chase, A.P.; Della Penna, A.; Gaube, P.; Morison, F.; et al. Small Phytoplankton Dominate Western North Atlantic Biomass. ISME J. 2020, 14, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, C.; Vincent, W.F.; Bonilla, S.; Roy, S.; Martineau, M.-J.; Terrado, R.; Potvin, M.; Massana, R.; Pedrós-Alió, C. Distribution, Phylogeny, and Growth of Cold-Adapted Picoprasinophytes in Arctic Seas. J. Phycol. 2007, 43, 78–89. [Google Scholar] [CrossRef]
- Metfies, K.; von Appen, W.-J.; Kilias, E.; Nicolaus, A.; Nöthig, E.-M. Biogeography and Photosynthetic Biomass of Arctic Marine Pico-Eukaroytes during Summer of the Record Sea Ice Minimum 2012. PLoS ONE 2016, 11, e0148512. [Google Scholar] [CrossRef]
- Lovejoy, C.; Potvin, M. Microbial Eukaryotic Distribution in a Dynamic Beaufort Sea and the Arctic Ocean. J. Plankton Res. 2011, 33, 431–444. [Google Scholar] [CrossRef]
- Simmons, M.P.; Bachy, C.; Sudek, S.; van Baren, M.J.; Sudek, L.; Ares, M.; Worden, A.Z. Intron Invasions Trace Algal Speciation and Reveal Nearly Identical Arctic and Antarctic Micromonas Populations. Mol. Biol. Evol. 2015, 32, 2219–2235. [Google Scholar] [CrossRef]
- Simon, N.; Foulon, E.; Grulois, D.; Six, C.; Desdevises, Y.; Latimier, M.; Le Gall, F.; Tragin, M.; Houdan, A.; Derelle, E.; et al. Revision of the Genus Micromonas Manton et Parke (Chlorophyta, Mamiellophyceae), of the Type Species M. pusilla (Butcher) Manton & Parke and of the Species M. commoda van Baren, Bachy and Worden and Description of Two New Species Based on the Genetic and Phenotypic Characterization of Cultured Isolates. Protist 2017, 168, 612–635. [Google Scholar] [CrossRef]
- Demory, D.; Baudoux, A.-C.; Monier, A.; Simon, N.; Six, C.; Ge, P.; Rigaut-Jalabert, F.; Marie, D.; Sciandra, A.; Bernard, O.; et al. Picoeukaryotes of the Micromonas Genus: Sentinels of a Warming Ocean. ISME J. 2018, 13, 132–146. [Google Scholar] [CrossRef]
- Worden, A.Z.; Lee, J.-H.; Mock, T.; Rouzé, P.; Simmons, M.P.; Aerts, A.L.; Allen, A.E.; Cuvelier, M.L.; Derelle, E.; Everett, M.V.; et al. Green Evolution and Dynamic Adaptations Revealed by Genomes of the Marine Picoeukaryotes Micromonas. Science 2009, 324, 268–272. [Google Scholar] [CrossRef]
- Li, W.K.W.; McLaughlin, F.A.; Lovejoy, C.; Carmack, E.C. Smallest Algae Thrive As the Arctic Ocean Freshens. Science 2009, 326, 539. [Google Scholar] [CrossRef]
- Riley, J.S.; Sanders, R.; Marsay, C.; Le Moigne, F.A.C.; Achterberg, E.P.; Poulton, A.J. The Relative Contribution of Fast and Slow Sinking Particles to Ocean Carbon Export: Export Of Fast And Slow Sinking Poc. Glob. Biogeochem. Cycles 2012, 26, GB1026. [Google Scholar] [CrossRef]
- Guidi, L.; Stemmann, L.; Jackson, G.A.; Ibanez, F.; Claustre, H.; Legendre, L.; Picheral, M.; Gorskya, G. Effects of Phytoplankton Community on Production, Size, and Export of Large Aggregates: A World-ocean Analysis. Limnol. Oceanogr. 2009, 54, 1951–1963. [Google Scholar] [CrossRef]
- Durkin, C.A.; Estapa, M.L.; Buesseler, K.O. Observations of Carbon Export by Small Sinking Particles in the Upper Mesopelagic. Mar. Chem. 2015, 175, 72–81. [Google Scholar] [CrossRef]
- Giering, S.L.C.; Sanders, R.; Lampitt, R.S.; Anderson, T.R.; Tamburini, C.; Boutrif, M.; Zubkov, M.V.; Marsay, C.M.; Henson, S.A.; Saw, K.; et al. Reconciliation of the Carbon Budget in the Ocean’s Twilight Zone. Nature 2014, 507, 480–483. [Google Scholar] [CrossRef]
- Omand, M.M.; D’Asaro, E.A.; Lee, C.M.; Perry, M.J.; Briggs, N.; Cetinić, I.; Mahadevan, A. Eddy-Driven Subduction Exports Particulate Organic Carbon from the Spring Bloom. Science 2015, 348, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.L.; Jackson, G.A. Small Phytoplankton and Carbon Export from the Surface Ocean. Science 2007, 315, 838–840. [Google Scholar] [CrossRef] [PubMed]
- Poff, K.E.; Leu, A.O.; Eppley, J.M.; Karl, D.M.; DeLong, E.F. Microbial Dynamics of Elevated Carbon Flux in the Open Ocean’s Abyss. Proc. Natl. Acad. Sci. USA 2021, 118, e2018269118. [Google Scholar] [CrossRef]
- Dall’Olmo, G.; Mork, K.A. Carbon Export by Small Particles in the Norwegian Sea. Geophys. Res. Lett. 2014, 41, 2921–2927. [Google Scholar] [CrossRef]
- Soltwedel, T.; Bauerfeind, E.; Bergmann, M.; Budaeva, N.; Hoste, E.; Jaeckisch, N.; von Juterzenka, K.; Matthiessen, J.; Mokievsky, V.; Nöthig, E.-M.; et al. HAUSGARTEN: Multidisciplinary Investigations at a Deep-Sea, Long-Term Observatory in the Arctic Ocean. Oceanography 2005, 18, 46–61. [Google Scholar] [CrossRef]
- von Appen, W.-J.; Waite, A.M.; Bergmann, M.; Bienhold, C.; Boebel, O.; Bracher, A.; Cisewski, B.; Hagemann, J.; Hoppema, M.; Iversen, M.H.; et al. Sea-Ice Derived Meltwater Stratification Slows the Biological Carbon Pump: Results from Continuous Observations. Nat. Commun. 2021, 12, 7309. [Google Scholar] [CrossRef]
- Limardo, A.J.; Sudek, S.; Choi, C.J.; Poirier, C.; Rii, Y.M.; Blum, M.; Roth, R.; Goodenough, U.; Church, M.J.; Worden, A.Z. Quantitative Biogeography of Picoprasinophytes Establishes Ecotype Distributions and Significant Contributions to Marine Phytoplankton. Environ. Microbiol. 2017, 19, 3219–3234. [Google Scholar] [CrossRef] [PubMed]
- Kremling, K.; Lentz, U.; Zeitzschel, B.; Schulz-Bull, D.E.; Duinker, J.C. New Type of Time-series Sediment Trap for the Reliable Collection of Inorganic and Organic Trace Chemical Substances. Rev. Sci. Instrum. 1996, 67, 4360–4363. [Google Scholar] [CrossRef]
- Zeitzschel, B.; Diekmann, P.; Uhlmann, L. A New Multisample Sediment Trap. Mar. Biol. 1978, 45, 285–288. [Google Scholar] [CrossRef]
- Metfies, K.; Bauerfeind, E.; Wolf, C.; Sprong, P.; Frickenhaus, S.; Kaleschke, L.; Nicolaus, A.; Nöthig, E.-M. Protist Communities in Moored Long-Term Sediment Traps (Fram Strait, Arctic)–Preservation with Mercury Chloride Allows for PCR-Based Molecular Genetic Analyses. Front. Mar. Sci. 2017, 4, 301. [Google Scholar] [CrossRef]
- Beszczynska-Möller, A.; Wisotzki, A. Physical Oceanography during POLARSTERN Cruise ARK-XXV/2. PANGAEA 2010. [Google Scholar] [CrossRef]
- Hamilton, M.; Mascioni, M.; Hehenberger, E.; Bachy, C.; Yung, C.; Vernet, M.; Worden, A.Z. Spatiotemporal Variations in Antarctic Protistan Communities Highlight Phytoplankton Diversity and Seasonal Dominance by a Novel Cryptophyte Lineage. mBio 2021, 12, e02973-21. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Vergin, K.L.; Beszteri, B.; Monier, A.; Thrash, J.C.; Temperton, B.; Treusch, A.H.; Kilpert, F.; Worden, A.Z.; Giovannoni, S.J. High-Resolution SAR11 Ecotype Dynamics at the Bermuda Atlantic Time-Series Study Site by Phylogenetic Placement of Pyrosequences. ISME J. 2013, 7, 1322–1332. [Google Scholar] [CrossRef]
- Choi, C.J.; Bachy, C.; Jaeger, G.S.; Poirier, C.; Sudek, L.; Sarma, V.V.S.S.; Mahadevan, A.; Giovannoni, S.J.; Worden, A.Z. Newly Discovered Deep-Branching Marine Plastid Lineages Are Numerically Rare but Globally Distributed. Curr. Biol. 2017, 27, R15–R16. [Google Scholar] [CrossRef]
- Choi, C.J.; Jimenez, V.; Needham, D.M.; Poirier, C.; Bachy, C.; Alexander, H.; Wilken, S.; Chavez, F.P.; Sudek, S.; Giovannoni, S.J.; et al. Seasonal and Geographical Transitions in Eukaryotic Phytoplankton Community Structure in the Atlantic and Pacific Oceans. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Guillou, L.; Bachar, D.; Audic, S.; Bass, D.; Berney, C.; Bittner, L.; Boutte, C.; Burgaud, G.; de Vargas, C.; Decelle, J.; et al. The Protist Ribosomal Reference Database (PR2): A Catalog of Unicellular Eukaryote Small Sub-Unit rRNA Sequences with Curated Taxonomy. Nucl. Acids Res. 2012, 41, D597–D604. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Demir-Hilton, E.; Sudek, S.; Cuvelier, M.L.; Gentemann, C.L.; Zehr, J.P.; Worden, A.Z. Global Distribution Patterns of Distinct Clades of the Photosynthetic Picoeukaryote Ostreococcus. ISME J. 2011, 5, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Tuerena, R.E.; Hopkins, J.; Buchanan, P.J.; Ganeshram, R.S.; Norman, L.; Appen, W.; Tagliabue, A.; Doncila, A.; Graeve, M.; Ludwichowski, K.U.; et al. An Arctic Strait of Two Halves: The Changing Dynamics of Nutrient Uptake and Limitation Across the Fram Strait. Glob. Biogeochem. Cycles 2021, 35. [Google Scholar] [CrossRef]
- Šlapeta, J.; López-García, P.; Moreira, D. Global Dispersal and Ancient Cryptic Species in the Smallest Marine Eukaryotes. Mol. Biol. Evol. 2006, 23, 23–29. [Google Scholar] [CrossRef]
- Worden, A.Z. Picoeukaryote Diversity in Coastal Waters of the Pacific Ocean. Aquat. Microb. Ecol. 2006, 43, 165–175. [Google Scholar] [CrossRef]
- Nöthig, E.-M.; Bracher, A.; Engel, A.; Metfies, K.; Niehoff, B.; Peeken, I.; Bauerfeind, E.; Cherkasheva, A.; Gäbler-Schwarz, S.; Hardge, K.; et al. Summertime Plankton Ecology in Fram Strait—A Compilation of Long- and Short-Term Observations. Polar Res. 2015, 34, 23349. [Google Scholar] [CrossRef]
- Foulon, E.; Not, F.; Jalabert, F.; Cariou, T.; Massana, R.; Simon, N. Ecological Niche Partitioning in the Picoplanktonic Green Alga Micromonas pusilla: Evidence from Environmental Surveys Using Phylogenetic Probes. Environ. Microbiol. 2008, 10, 2433–2443. [Google Scholar] [CrossRef]
- Kilias, E.; Wolf, C.; Nöthig, E.-M.; Peeken, I.; Metfies, K. Protist Distribution in the Western Fram Strait in Summer 2010 Based on 454-Pyrosequencing of 18S rDNA. J. Phycol. 2013, 49, 996–1010. [Google Scholar] [CrossRef]
- Soltwedel, T.; Bauerfeind, E.; Bergmann, M.; Bracher, A.; Budaeva, N.; Busch, K.; Cherkasheva, A.; Fahl, K.; Grzelak, K.; Hasemann, C.; et al. Natural Variability or Anthropogenically-Induced Variation? Insights from 15 Years of Multidisciplinary Observations at the Arctic Marine LTER Site HAUSGARTEN. Ecol. Indic. 2015, 65, 89–102. [Google Scholar] [CrossRef]
- Lalande, C.; Nöthig, E.-M.; Bauerfeind, E.; Hardge, K.; Beszczynska-Möller, A.; Fahl, K. Lateral Supply and Downward Export of Particulate Matter from Upper Waters to the Seafloor in the Deep Eastern Fram Strait. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 114, 78–89. [Google Scholar] [CrossRef]
- Gradinger, R.R.; Baumann, M.E.M. Distribution of Phytoplankton Communities in Relation to the Large-Scale Hydrographical Regime in the Fram Strait. Mar. Biol. 1991, 111, 311–321. [Google Scholar] [CrossRef]
- Lampe, V.; Nöthig, E.-M.; Schartau, M. Spatio-Temporal Variations in Community Size Structure of Arctic Protist Plankton in the Fram Strait. Front. Mar. Sci. 2021, 7, 579880. [Google Scholar] [CrossRef]
- Lalande, C.; Bauerfeind, E.; Nöthig, E. Downward Particulate Organic Carbon Export at High Temporal Resolution in the Eastern Fram Strait: Influence of Atlantic Water on Flux Composition. Mar. Ecol. Prog. Ser. 2011, 440, 127–136. [Google Scholar] [CrossRef]
- Kilias, E.S.; Nöthig, E.-M.; Wolf, C.; Metfies, K. Picoeukaryote Plankton Composition off West Spitsbergen at the Entrance to the Arctic Ocean. J. Eukaryot. Microbiol. 2014, 61, 569–579. [Google Scholar] [CrossRef]
- Monier, A.; Worden, A.Z.; Richards, T.A. Phylogenetic Diversity and Biogeography of the Mamiellophyceae Lineage of Eukaryotic Phytoplankton across the Oceans. Environ. Microbiol. Rep. 2016, 8, 461–469. [Google Scholar] [CrossRef]
- Needham, D.M.; Fuhrman, J.A. Pronounced Daily Succession of Phytoplankton, Archaea and Bacteria Following a Spring Bloom. Nat. Microbiol. 2016, 1, 16005. [Google Scholar] [CrossRef]
- Buesseler, K.O.; Antia, A.N.; Chen, M.; Fowler, S.W.; Gardner, W.D.; Gustafsson, O.; Harada, K.; Michaels, A.F.; Rutgers van der Loeff, M.; Sarin, M.; et al. An Assessment of the Use of Sediment Traps for Estimating Upper Ocean Particle Fluxes. J. Mar. Res. 2007, 65, 345–416. [Google Scholar] [CrossRef]
- van Baren, M.J.; Bachy, C.; Reistetter, E.N.; Purvine, S.O.; Grimwood, J.; Sudek, S.; Yu, H.; Poirier, C.; Deerinck, T.J.; Kuo, A.; et al. Evidence-Based Green Algal Genomics Reveals Marine Diversity and Ancestral Characteristics of Land Plants. BMC Genom. 2016, 17, 267. [Google Scholar] [CrossRef]
- Simmons, M.P.; Sudek, S.; Monier, A.; Limardo, A.J.; Jimenez, V.; Perle, C.R.; Elrod, V.A.; Pennington, J.T.; Worden, A.Z. Abundance and Biogeography of Picoprasinophyte Ecotypes and Other Phytoplankton in the Eastern North Pacific Ocean. Appl. Environ. Microbiol. 2016, 82, 1693–1705. [Google Scholar] [CrossRef] [PubMed]
- Sherr, E.B.; Sherr, B.F.; Wheeler, P.A.; Thompson, K. Temporal and Spatial Variation in Stocks of Autotrophic and Heterotrophic Microbes in the Upper Water Column of the Central Arctic Ocean. Deep Sea Res. Part I Oceanogr. Res. Pap. 2003, 50, 557–571. [Google Scholar] [CrossRef]
- Li, L.; Wang, S.; Wang, H.; Sahu, S.K.; Marin, B.; Li, H.; Xu, Y.; Liang, H.; Li, Z.; Cheng, S.; et al. The Genome of Prasinoderma coloniale Unveils the Existence of a Third Phylum within Green Plants. Nat. Ecol. Evol. 2020, 4, 1220–1231. [Google Scholar] [CrossRef]
- Kilias, E.; Kattner, G.; Wolf, C.; Frickenhaus, S.; Metfies, K. A Molecular Survey of Protist Diversity through the Central Arctic Ocean. Polar Biol. 2014, 37, 1271–1287. [Google Scholar] [CrossRef]
- Marquardt, M.; Vader, A.; Stübner, E.I.; Reigstad, M.; Gabrielsen, T.M. Strong Seasonality of Marine Microbial Eukaryotes in a High-Arctic Fjord (Isfjorden, West Spitsbergen). Appl. Environ. Microbiol. 2016, 82, 1868–1880. [Google Scholar] [CrossRef]
- Balzano, S.; Marie, D.; Gourvil, P.; Vaulot, D. Composition of the Summer Photosynthetic Pico and Nanoplankton Communities in the Beaufort Sea Assessed by T-RFLP and Sequences of the 18S RRNA Gene from Flow Cytometry Sorted Samples. ISME J. 2012, 6, 1480–1498. [Google Scholar] [CrossRef]
- Clayton, S.; Lin, Y.-C.; Follows, M.J.; Worden, A.Z. Co-Existence of Distinct Ostreococcus Ecotypes at an Oceanic Front. Limnol. Oceanogr. 2017, 62, 75–88. [Google Scholar] [CrossRef]
- Belevich, T.A.; Milyutina, I.A.; Troitsky, A.V. Seasonal Variability of Photosynthetic Microbial Eukaryotes (<3 μm) in the Kara Sea Revealed by 18S rDNA Metabarcoding of Sediment Trap Fluxes. Plants 2021, 10, 2394. [Google Scholar] [CrossRef]
- Wietz, M.; Bienhold, C.; Metfies, K.; Torres-Valdés, S.; von Appen, W.-J.; Salter, I.; Boetius, A. The Polar Night Shift: Seasonal Dynamics and Drivers of Arctic Ocean Microbiomes Revealed by Autonomous Sampling. ISME Commun. 2021, 1, 76. [Google Scholar] [CrossRef]
- Cruz, B.N.; Brozak, S.; Neuer, S. Microscopy and DNA -based Characterization of Sinking Particles at the Bermuda Atlantic Time-series Study Station Point to Zooplankton Mediation of Particle Flux. Limnol. Oceanogr. 2021, 66, 3697–3713. [Google Scholar] [CrossRef]
- Potvin, M.; Lovejoy, C. PCR-Based Diversity Estimates of Artificial and Environmental 18S rRNA Gene Libraries. J. Eukaryot. Microbiol. 2009, 56, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Krumpen, T.; Gerdes, R.; Haas, C.; Hendricks, S.; Herber, A.; Selyuzhenok, V.; Smedsrud, L.; Spreen, G. Recent Summer Sea Ice Thickness Surveys in Fram Strait and Associated Ice Volume Fluxes. Cryosphere 2016, 10, 523–534. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bachy, C.; Sudek, L.; Choi, C.J.; Eckmann, C.A.; Nöthig, E.-M.; Metfies, K.; Worden, A.Z. Phytoplankton Surveys in the Arctic Fram Strait Demonstrate the Tiny Eukaryotic Alga Micromonas and Other Picoprasinophytes Contribute to Deep Sea Export. Microorganisms 2022, 10, 961. https://doi.org/10.3390/microorganisms10050961
Bachy C, Sudek L, Choi CJ, Eckmann CA, Nöthig E-M, Metfies K, Worden AZ. Phytoplankton Surveys in the Arctic Fram Strait Demonstrate the Tiny Eukaryotic Alga Micromonas and Other Picoprasinophytes Contribute to Deep Sea Export. Microorganisms. 2022; 10(5):961. https://doi.org/10.3390/microorganisms10050961
Chicago/Turabian StyleBachy, Charles, Lisa Sudek, Chang Jae Choi, Charlotte A. Eckmann, Eva-Maria Nöthig, Katja Metfies, and Alexandra Z. Worden. 2022. "Phytoplankton Surveys in the Arctic Fram Strait Demonstrate the Tiny Eukaryotic Alga Micromonas and Other Picoprasinophytes Contribute to Deep Sea Export" Microorganisms 10, no. 5: 961. https://doi.org/10.3390/microorganisms10050961
APA StyleBachy, C., Sudek, L., Choi, C. J., Eckmann, C. A., Nöthig, E.-M., Metfies, K., & Worden, A. Z. (2022). Phytoplankton Surveys in the Arctic Fram Strait Demonstrate the Tiny Eukaryotic Alga Micromonas and Other Picoprasinophytes Contribute to Deep Sea Export. Microorganisms, 10(5), 961. https://doi.org/10.3390/microorganisms10050961