Abstract
The aim of this study was to reveal the sites of yeast contamination in dairy production and perform taxonomic characterization of potential yeast spoilers in cheese making. Occurrence of spoilage yeasts was followed throughout the manufacture of white-brined cheese at a Danish dairy, including the areas of milk pasteurization, curd processing, and packaging (26 sites in total). Spoilage yeasts were isolated from whey, old cheese curd, and air samples in viable counts of 1.48–6.27 log CFU/mL, 5.44 log CFU/g, and 1.02 log CFU/m3, respectively. Yeast isolates were genotypically classified using (GTG)5-PCR fingerprinting and identified by sequencing of the D1/D2 region of the 26S rRNA gene. The largest yeast heterogeneity was found in old curd collected under the turning machine of molds, where 11 different yeast species were identified. The most frequently isolated yeast species were Candida intermedia, Kluyveromyces marxianus, and Pichia kudriavzevii. The less abundant yeast species included Candida auris, Candida parapsilosis, Candida pseudoglaebosa, Candida sojae, Cutaneotrichosporon curvatus, Cutaneotrichosporon moniliiforme, Papiliotrema flavescens, Rhodotorula mucilaginosa, Vanrija humicola, and Wickerhamiella sorbophila. The awareness on occurrence and taxonomy of spoilage yeasts in cheese production will contribute to a knowledge-based control of contaminating yeasts and quality management of cheese at the dairies.
1. Introduction
White-brined cheeses, such as Feta, Domiati, and Halloumi, originate from the Mediterranean region and the Middle East [1]. Different varieties of white-brined cheese have traditionally been made from ovine, caprine, bovine or cow milk, and nowadays, they are industrially produced or at an artisanal scale in Europe, Turkey, Northern Africa, and some regions of Asia and South America. The main production steps of white-brined cheese involve milk pasteurization, addition of starter cultures and rennet, followed by milk coagulation and curd formation. Subsequently, the curd is drained, salted, and ripened into brine to obtain the final product [2,3,4].
Yeasts are the major spoilage microorganisms in white-brined cheeses. Depending on the species, yeasts can readily utilize milk carbohydrates, such as lactose and galactose, organic acids, and proteins, and grow at dairy-relevant conditions, i.e., refrigerated temperatures, acidic pH, reduced water activity, and high salt concentrations [5,6,7]. The most common yeast spoilers in white-brined cheeses belong to the genera Candida, Cryptococcus, Debaryomyces, Geotrichum, Kluyveromyces, Pichia, Rhodotorula, Saccharomyces, Torulospora, Trichosporon, Yarrowia, and Zygosaccharomyces [7,8,9]. When grown in high numbers (typically 5–6 log CFU/g), the enzymatic activities of yeasts lead to various quality defects, such as off-flavors, discoloration, the swelling of the cans, and the softening of the cheese texture [1,5,7,9,10].
The common sources of yeast contamination in dairy plants include production facilities, raw materials, brine, air, wooden shelves of ripening, and personnel [5,6,7,8,11]. Yeast occurrence is mostly attributed to their survival through sanitizers, as well as cross-contamination via processing environment and air [6,9,12]. Various studies have demonstrated that aerosols, in form of droplets or dust, can be easily dispersed by air flow, introducing a major way of yeast transmission in dairy environment [13]. Stobnicka-Kupiec et al. [14] identified Candida spp., Cryptococcus spp., Debaryomyces hansenii, Geotrichum candidum, Rhodotorula spp., and Yarrowia lipolytica in the air and surface samples of commercial and traditional Polish dairy plants (with expertise in milk, butter, cream, yoghurt, and cheese production; not including mold-ripened cheese), where the highest concentration of yeasts and molds was found on the worktops in the milk reception and cheese production areas and in the air samples. An earlier study revealed that the major fungal loads in the air of outdoor and indoor locations of a Greek dairy plant was presented by Cladosporium spp., Penicillium spp., and some unidentified yeasts [15].
Traditional dairy products are especially prone to microbial contamination, mostly due to the usage of unpasteurized milk [1,14,16]. At commercial dairies, production processes are based on specific management systems, including GMP (Good Manufacturing Practices), GHP (Good Hygiene Practices), SOP (Standard Operating Practices), and the universal Hazard Analysis and Critical Control Point (HACCP) system, targeting prevention and control of food contamination [9,14,15,17,18]. Despite of the implementation of the quality-management systems, yeast spoilage is still of concern in the dairy industry.
Identification of yeasts at species and strain level is essential to trace the ways of yeast contamination in dairy production and evaluate their spoilage potential. Most of the published studies revealed the spots of contamination at specific dairies and quantified the yeast loads, while only a few of them proceeded with taxonomic characterization of the isolated yeasts [14,16,17]. The aim of this study was therefore to reveal the hotspot areas of yeast contamination throughout production of white-brined cheese at a Danish dairy and characterize the taxonomic diversity of potential yeast spoilers.
2. Materials and Methods
2.1. Sample Collection
Sampling was performed during two visits at a Danish dairy production of white-brined cheese, in November 2019 (Trial 1) and January 2020 (Trial 2), referred to as T1 and T2, respectively (Table 1 and Table 2). In T1, 47 samples were collected from 24 different sites in production of white-brined cheese including milk, curd, cheese, brine, ingredients, air, and swab from the cheese cutter. The second visit (T2) was mostly carried out as a supplementary sampling at the hotspots areas revealed in T1. In total, 22 samples from 11 different sites were collected in T2 including curd, whey, cheese, and air.

Table 1.
Viable counts of yeasts in milk, whey, and cheese samples from white-brined cheese production.
Table 2.
Viable counts of yeasts in environmental samples from white-brined cheese production.
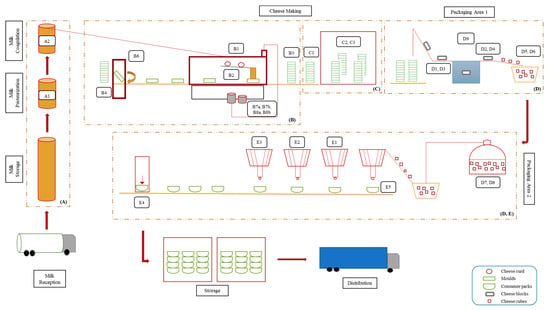
Figure 1 presents the scheme of cheese production line with highlighted sampling sites and sample codes. The production area (A) refers to the cheese vats, (B) and (C) to the mechanical tunnel and draining room, respectively, and (D) and (E) to the packaging areas. Major production steps included milk pasteurization (Sample A1), followed by the addition of rennet and starter cultures, which allowed milk coagulation and curd formation (Sample A2). Afterwards, the curd was added into molds (Samples B3 and C1), drained for 20–26 h at 13 °C (Sample C2), and cut into cubes (Sample D5). The cubes were immerged into 12% (w/v) brine for 12–72 h at 5 °C (Samples D7 and D8). Finally, the cubes were mixed with other ingredients such as oil, vegetables, and herbs (Samples E1, E2, E3, and E5), filled in “consumer pack” containers (Sample E4), and finally transported to the storage room until their distribution.
2.2. Enumeration and Isolation of Yeasts
Solid samples of curd, cheese, and ingredients in amounts of 10 g were mixed with 90 mL of Saline Peptone Diluent (SPO) (0.1% w/v peptone, 0.03% w/v Na2HSO4, 1% w/v NaCl, pH 5.6 ± 0.1) and homogenized in a Stomacher Bags Mixer (InterScience, Saint Nom la Bretêche, France) for 180 s at high speed. Whey samples were additionally incubated at 25 °C for 48 h, before yeast enumeration, as an enrichment step to promote yeast growth. The swab sample from the cube cutter (15 × 15 mm cube size, random sampling in different parts of the cube cutter) was collected using Compact Dry Swab (HyServe, Kirchheim bei München, Germany) in 1 mL SPO added 1% (w/v) NaCl. Air samples of 500 L were collected with an air sampler Sampl’air Lite (AES Laboratoire, BioMérieux, Marcy-l’Étoile, France). Serial dilutions of the samples were prepared with Pro-Media MT-11PBS diluent (Elmex, Tokyo, Japan), and plated in duplicates on Malt Yeast Glucose Peptone (MYGP) agar (0.3% w/v malt extract, 0.3% w/v yeast extract, 1% w/v D-Glucose monohydrate, 0.5% w/v peptone, 2% w/v agar, 1% w/v NaCl, supplemented with 100 mg/L of chloramphenicol and 50 mg/L of chlortetracycline, pH 5.6 ± 0.1). For CFU enumeration, plates with 20–300 colonies were selected and the results are presented as average values of log10 and their standard deviations are calculated. For brine analysis, SPO and MYGP agar, supplemented with 4% (w/v) NaCl, were used in order to mimic the brine environment. Plates were incubated under aerobic conditions at 25 °C for 3–5 days. Representative colonies, randomly selected (10–20 colonies depending on the CFU counts), were purified by streaking on MYGP agar plates and grouped based on their micro- and macromorphological characteristics (surface, margin, profile, size, and color). The number of isolates for the subsequent rep-PCR (Repetitive Extragenic Palindromic Polymerase Chain Reaction) analysis was calculated as the square root of the number of colonies from each colony group. In case of low counts (Samples B6 and D1), all colonies were isolated and purified. Selected isolates were grown in MYGP broth overnight and stored at −80 °C in 20% v/v glycerol. All reagents were purchased from Sigma A/S (Søborg, Denmark) or Merck (Søborg, Denmark), unless otherwise specified.
2.3. Molecular Characterization of Yeast Isolates
2.3.1. Rep–PCR
The total yeast DNA was extracted from the colonies grown on MYGP agar plates using the InstaGene Matrix DNA extraction kit (Bio-Rad Laboratories, Hercules, CA, USA). Rep-PCR was conducted in a 25 μL volume mixture containing 13 μL of Taq DNA Polymerase 2× Master Mix RED (Ampliqon, Odense, Denmark), 5 μL Primer GTG5 (Integrated DNA Technologies, Denmark), 4 μL sterile Milli-Q water, and 3 μL DNA. The PCR reaction was carried out in a SureCycler 8800 thermocycler (Agilent Technologies, Santa Clara, CA, USA) using the following program: initial denaturation for 7 min at 95 °C, followed by 30 cycles of 95 °C for 1 min, 45 °C for 1 min, and 65 °C for 8 min, and an elongation step of 65 °C for 16 min. The rep-PCR products were separated by 1.5% agarose gel electrophoresis (5 h, 120 V) in Tris-Borate-EDTA buffer (0.5 × TBE), using an O’GeneRuler 1 kb DNA ladder (Thermo Scientific, Roskilde, Denmark) as a reference marker. The rep-PCR profiles were clustered using Bionumerics 7.1 software (Applied Maths, BioMérieux, Schaerbeek, Belgium) based on Dice’s Coefficient of similarity with the Unweighted Pair Group Method and Arithmetic mean clustering algorithm (UPGMA).
2.3.2. 26S rRNA Gene Sequencing
Sequencing of the D1/D2 region of the 26S rRNA gene was performed for all the isolates clustered (39 in total) using the primers NL-1 (5′-GCA TAT CAA TAA GCG GAG GAA AAG-3′) and NL-4 (5′-GGT CCG TGT TTC AAG ACG G-3′), as described by Van Der Aa Kühle and Jespersen [19]. Shortly, the PCR reaction was conducted in a 50 μL volume mixture containing 25 μL of Taq DNA Polymerase 2× Master Mix RED (Ampliqon, Odense, Denmark), 5 μL Primer mix NL-1 and NL-4, 17 μL sterile Milli-Q water, and 3 μL of the total DNA from yeast. The PCR reaction was carried out in a SureCycler 8800 thermocycler (Agilent Technologies, Santa Clara, CA, USA), using the following program: initial denaturation for 5 min at 95 °C, followed by 30 cycles at 95 °C for 90 s, 53 °C for 30 s, and 72 °C for 90 s, and the final elongation step at 72 °C for 7 min. The DNA sequencing (using the same primers; NL1 and NL4) was performed by Macrogen (Amsterdam, The Netherlands). Sequences were manually corrected, assembled with CLC Genomics Workbench version 7.9.1 software (QIAGEN Digital Insights, Redwood City, CA, USA), and compared to the reported 26S rRNA gene sequences in GenBank using the Basic Local Alignment Search Tool (BLAST) algorithm. The nucleotide sequences have been deposited in GenBank under Accession Numbers OL744629–OL744667, as indicated in Table 3.
Table 3.
Identification of yeasts from the white-brined cheese production by sequencing the D1/D2 region of the 26S rRNA gene.
2.3.3. Sequencing of the 5.8S rDNA-ITS Region
Sequencing of the Internal Transcribed Spacer (ITS) (ITS1-5.8S rDNA-ITS2 region) was additionally performed to differentiate closely related species of Kluyveromyces marxianus and Kluyveromyces lactis that were not differentiated by sequencing the D1/D2 region of the 26S rRNA gene [20]. For amplification of the 5.8S-ITS fragment, primers ITS-1 (5′-TCC GTA GGT GAA CCT GCG G-3′) and ITS-4 (5′-TCC TCC GCT TAT TGA TAT GC-3′) were used as previously described [19,21,22]. Shortly, the PCR reaction was conducted in a 50 μL volume mixture containing 25 μL of Taq DNA Polymerase 2× Master Mix RED (Ampliqon, Denmark), 5 μL Primer mix ITS-1 and ITS-4, 17 μL sterile Milli-Q water and 3 μL of samples’ DNA. The PCR reaction was carried out with a SureCycler 8800 (Agilent Technologies, Santa Clara, CA, USA) under the following conditions: 3 min of initial denaturation at 95 °C, followed by 30 cycles of 95 °C for 30 s, 60 °C for 40 s, 72 °C for 30 s, and the final elongation step at 72 °C for 10 min. The PCR products were treated as in Section 2.3.2. of this publication.
2.4. Phenotypic Tests
The isolates of Kluyveromyces spp. (6 isolates in total) were tested for assimilation and fermentation of glucose, lactose, maltose, galactose, sucrose, and raffinose to distinguish the species of K. marxianus and K. lactis [23]. Isolates were grown on MYGP agar for 3 days at 25 °C before the experiments. For fermentation tests, 5 mL medium (0.45% w/v yeast extract; 0.75% w/v peptone; 12 mL Bromothymol blue solution; and up to 1000 mL distilled water, pH 6.0 ± 0.2) was added into Durham tubes. The medium for assimilation tests (0.067% w/v Difco Yeast Nitrogen Base, 0.05% w/v Carbohydrate, pH 5.6 ± 0.1) was added in a volume of 0.5 mL into the respective test tubes. Solutions of glucose, lactose, maltose, galactose, sucrose (6% w/v each), and raffinose (12% w/v) were distributed in the tubes for fermentation (2.5 mL) and assimilation (0.5 mL) tests, and afterwards, yeast-culture suspension of 0.1 mL (yeast colony material mixed with 4.5 mL sterile water) was transferred into the test tubes. The tubes were incubated at 25 °C for 4 weeks and checked weekly for gas production and/or change of the color (fermentation test) and using a Wickerhams card (assimilation test) [23]. Strains of Kluyveromyces marxianus CBS 1553 and Kluyveromyces lactis CBS 845 were obtained from the CBS culture collection (Centraalbureau voor Schimmelcultures, CBS, The Netherlands) and used as positive controls.
3. Results and Discussion
3.1. Yeast Viable Counts
The viable counts of yeasts in milk (2 samples), whey (4 samples), curd (18 samples), cheese (24 samples), and brine (2 samples) are presented in Table 1. The levels of yeasts in raw materials (6 samples), air (12 samples), and swab (1 sample) are shown in Table 2. The highest yeast counts of 5.44 ± 1.07 log CFU/g were detected in the old curd under the turning machine (sample B4). Additionally, yeasts were found in recirculated whey (separated from cheese fines) at levels of 1.48 ± 0.01 log CFU/mL (sample B7b, before the enrichment step) and 6.27 ± 0.01 log CFU/mL (sample B8b, after the enrichment step). Yeast concentration in the air from the draining room (sample C3) was 1.02 ± 0.12 log CFU/m3. The yeast counts in all other production samples were below the detection limit (less than 2 log CFU/g for solid samples or less than 1 log CFU/mL, m3 or m2 for liquid, air, and swab samples, respectively). Despite undetectable levels of yeast counts, especially in the final product (E4), possibility of yeast contamination and subsequent cheese spoilage during storage cannot be excluded. Air, cheese curd, and whey have previously been recognized as the major sources of yeast contamination in dairy production. Yeast counts in curd under the turning machine in this study were close to the values reported for curd in Pecorino Crotonese cheese manufacture (3.7 log CFU/g) and Bryndza cheese produced from raw ewe milk (4.1–6.2 log CFU/g) [12,24]. Besides, the CFU counts in recirculated whey were lower than the numbers reported for whey from production of artisanal white-pickled cheese (2.5–5.6 log CFU/mL) [4,25]. The absence of yeast contamination in brine in this study is in contrast with the results reported for white-pickled and Feta cheeses (5–7 log CFU/mL) [4,25]. Discrepancies between the results can be explained by the use of fresh brine, in this study (6 weeks old with an expiration period of at least 6 months), as well as by differences in production processes and hygiene conditions between the dairies.
Supporting our results, microbial air loads of 1–3 log CFU/m3 have commonly been reported in dairy plants [18,26,27]. Until now, there is no legislation concerning the limits of airborne contaminants in dairy production, though several scientific recommendations have been published [28,29,30,31]. Thus, according to the American Public Health Association (APHA), the fungal loads in dairy facilities should not exceed the range of 1.8–2.6 log CFU/m3 [30,31]. Consequently, the levels of yeasts in the air samples in this study are in accordance with the recommended limits. However, there is no doubt that the contamination from air should be avoided, e.g., by prevention of aerosols caused by high pressure flushing.
3.2. Phylogenetic Characterization of Yeasts
In total, 99 morphologically distinct isolates were purified from 69 samples in the white-brined cheese production. Among them, 39 isolates were clustered using rep-PCR and further identified by sequencing the D1/D2 region of the 26S rRNA gene and the 5.8S rDNA-ITS region. Figure 2 presents the phylogenetic dendrogram of these isolates based on the rep-PCR fingerprints. Using a similarity cut-off of 85%, the isolates were grouped into 13 clusters. Major clusters were comprised of species Pichia kudriavzevii (formerly Issatchenkia orientalis, anamorph Candida krusei) (Cluster 3), Candida intermedia (Cluster 8), and K. marxianus (anamorph Candida kefyr) (Cluster 10). The Candida spp., K. marxianus, P. kudriavzevii, and Wickerhamiella sorbophila (syn. Candida sorbophila) have been assigned to the division of Ascomycota, class of Saccharomycetes. Candida was the most diverse genus in this study, represented by five species, namely Candida auris, C. intermedia, Candida parapsilosis, Candida pseudoglaebosa, and Candida sojae.
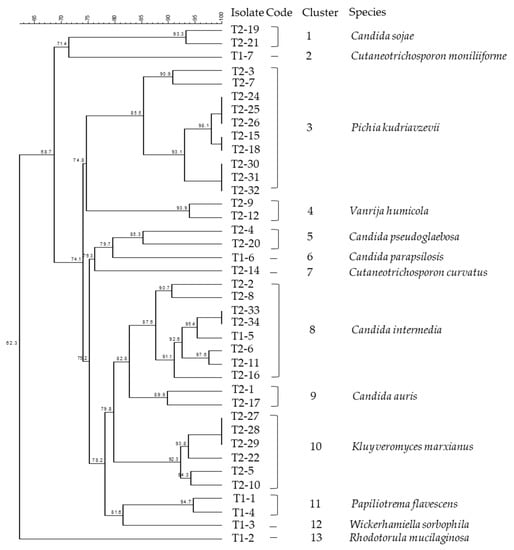
Figure 2.
Dendrogram of (GTG)5-PCR fingerprints of yeast isolates collected from the production line of white-brined cheese. Clustering was based on Dice’s coefficient of similarity with the unweighted pair-group method with arithmetic average clustering algorithm (UPGMA). Numbers of nodes in the dendrogram denote the percentage of similarity between the clusters.
Other identified species, Cutaneotrichosporon curvatus (formerly Candida curvata, syn. Cryptococcus curvatus), Cutaneotrichosporon moniliiforme (formerly Trichosporon moniliiforme), Paliliotrema flavescens (formerly Cryptococus flavescens), Rhodotorula mucilaginosa, and Vanrija humicola (formerly Cryptococcus humicola) belong to the division of Basidiomycota, classes of Microbotryomycetes and Tremellomycetes. The taxonomic identities of the isolates from production samples and their GenBank Accession Numbers are listed in Table 3. The obtained sequences were annotated to 13 yeast species showing 99.7–100% similarity to the reference GenBank sequences.
3.3. Phylogenetic Characterization of Yeasts
In total, 99 morphologically distinct isolates were purified from the white-brined cheese production. Among them, 39 isolates were clustered using rep-PCR and further identified by sequencing the D1/D2 region of the 26S rRNA gene and the 5.8S rDNA-ITS region. Figure 2 presents the phylogenetic dendrogram of these isolates based on the rep-PCR fingerprints. Using a similarity cutoff of 85%, the isolates were grouped into 13 clusters. Major clusters were comprised of species Pichia kudriavzevii (formerly Issatchenkia orientalis, anamorph Candida krusei) (Cluster 3), Candida intermedia (Cluster 8), and K. marxianus (anamorph Candida kefyr) (Cluster 10). Candida spp., K. marxianus, P. kudriavzevii, and Wickerhamiella sorbophila (syn. Candida sorbophila) were assigned to the division of Ascomycota, class of Saccharomycetes. Candida was the most diverse genus in this study, represented by five species, namely Candida auris, C. intermedia, Candida parapsilosis, Candida pseudoglaebosa, and Candida sojae. Other identified species, Cutaneotrichosporon curvatus (formerly Candida curvata, syn. Cryptococcus curvatus), Cutaneotrichosporon moniliiforme (formerly Trichosporon moniliiforme), Paliliotrema flavescens (formerly Cryptococus flavescens), Rhodotorula mucilaginosa, and Vanrija humicola (formerly Cryptococcus humicola) belong to the division of Basidiomycota, classes of Microbotryomycetes and Tremellomycetes. The taxonomic identities of the isolates from production samples and their GenBank Accession Numbers are listed in Table 3. The obtained sequences were annotated to 13 yeast species, showing 99.7–100% similarity to the reference GenBank sequences.
Due to close genotypic relatedness, the species K. marxianus and K. lactis could not be distinguished by 26S rRNA gene sequencing in this study. Based on the sequence analysis of the 5.8S rDNA-ITS region, the isolates were annotated to K. marxianus with 100% homology to GenBank. Additionally, phenotypic tests showed that the isolates could readily utilize and ferment glucose, galactose, sucrose, lactose, and weakly raffinose (Supplementary Table S1). Concurrently, no fermentation of maltose was observed during the 4-week incubation period, indicating that all Kluyveromyces isolates belong to K. marxianus [32].
3.4. Diversity of Yeasts in Cheese Production
The predominant species in white-brined cheese production were P. kudriavzevii (26% of total isolates), C. intermedia (20% of total isolates), and K. marxianus (15% of total isolates) identified from whey samples (B7b, B8b), old curd (B4), cheese on conveyor belt (D1), and air (B6). The largest abundance and diversity of yeast species (64% of the isolates) was found in the old curd (B4), presenting a nutrient rich substrate for yeast growth (Table 3). Species C. intermedia was predominantly found in the old curd (24% of curd isolates) and prevailed in cheese samples from conveyor belt (D1). Other species, isolated from the old curd, were C. auris (5%), C. pseudoglaebosa (5%), C. sojae (5%), C. curvatus (3%), P. flavescens (5%), R. mucilaginosa (3%), V. humicola (5%), and W. sorbophila (3%) (Table 3). Species C. parapsilosis (50%) and C. moniliiforme (50%) were identified in the air from the draining room (C3). Among them, the species of C. intermedia, K. lactis, P. kudriavzevii, and R. mucilaginosa are well-known contaminants in white-brined cheese [7,33,34,35].
It has been reported that Candida spp., such as C. parapsilosis and C. sojae, can be easily distributed through the dairy-processing environment and air [6,14]. In accordance with this study, predominance of C. intermedia has been demonstrated in the cheese-production environment [36], as well as in various types of cheeses, such as Serro Minas [7], Pecorino Crotonese [12], and cheese brines [37,38]. C. sojae has occasionally been detected in yoghurts [6], while C. pseudoglaebosa has been found in raw milk from dairy farms in France [39,40]. In contrast, species C. auris is not of dairy origin but rather referred to as a human pathogen frequently isolated from the human body fluids and tissues [41,42]. Thus, curd contamination with C. auris in this study would probably rise from direct contact with personnel at the dairy.
C. parapsilosis was a rare species in this study, detected only in the air from the draining room (1 out of 2 isolates). It is a common spoilage organism in various types of cheeses (brined, ripened, Swiss-type, blue-veined), characterized by high proteolytic and lipolytic activities [7,11,43]. Similar to C. auris, C. parapsilosis is considered as an opportunistic human pathogen causing invasive candidiasis [11,44,45]. At the same time, C. parapsilosis is regularly encountered in other fermented products, having positive impact on their organoleptic properties. To the best of our knowledge, species of C. auris and C. parapsilosis have not been linked to any outbreak of infection.
Along with the old curd, K. marxianus and P. kudriavzevii were the major species detected in whey samples before and after the enrichment step (B7b and B8b). Furthermore, K. marxianus was identified in the air sample (B6). Species P. kudriavzevii is a common inhabitant of various fermented products, able to grow at low pH and high salt concentration [46,47]. Predominance of K. marxianus can be due to its ability to ferment residual lactose in cheese [20,48], and assimilate lactate produced by lactic-acid bacteria [49,50,51]. High prevalence of these species in white-brined cheese production is in agreement with several studies. Both K. marxianus and P. kudriavzevii have been reported as predominant yeast contaminants at different stages of May Bryndza cheese production from dairies in Slovakia and in Portuguese Serpa cheese [24,52,53]. In addition, K. marxianus was identified in Serro Minas and water buffalo Mozzarella cheeses [7,54], while P. kudriavzevii has previously been isolated from British Wensleydale cheese [11] and fresh curd of artisanal Canastra cheese [43].
Other less frequent species from white-brined cheese production were C. curvatus, P. flavescens, and V. humicola detected in old curd (B4), and C. moniliiforme in the air from the draining room (C3). These species formerly belonged to the genus Cryptococcus, but after a recent reclassification they were assigned to the mentioned genera [55]. In accordance with this study, a few incidences of C. curvatus have been reported in the air of dairy environments [56], whereas P. flavescens has rarely been identified in dairy matrices such as ice cream and cheese brine [38,45]. The presence of C. moniliiforme is undesirable in white-brined cheese, as it is able to form melanin-like brown pigments, leading to product discoloration [57]. Cutaneotrichosporon spp., P. flavescens, and V. humicola are not typical dairy spoilers, but rather found in various natural environments (i.e., plants, bird feces, and tree hollows) [58]. Therefore, their ability to grow in white-brined cheese needs further investigation.
The species R. mucilaginosa and W. sorbophila were detected in low frequency in cheese curd. Similarly, W. sorbophila has been found in low numbers in buffalo Mozzarella cheese [54], while R. mucilaginosa has occasionally been isolated from production of various cheeses (e.g., Serro Minas, Bryndza cheese) [7,52]. Furthermore, R. mucilaginosa is considered an emerging opportunistic pathogen causing fungemia in humans [59,60]. Likewise with other Rhodotorula species, R. mucilaginosa may impair the cheese quality, giving rise to yellow/red pigmentation on the cheese surface attributed to carotenoid production (e.g., β-carotene, torulene, torularhobin) [61]. Thus, despite the low numbers, the effect of R. mucilaginosa on cheese shelf life and safety should not be underestimated.
4. Conclusions
A high diversity of yeast species (13 in total) was found in the production of white-brined cheese at a Danish dairy. Most of the yeast species (i.e., C. intermedia, K. marxianus, P. kudriavzevii) have been characterized as typical dairy spoilers, while the others (i.e., C. auris, C. parapsilosis, Cutaneotrichosporon spp., P. flavescens, V. humicola) originate from different sources, such as plants, humans, and other environments. The contamination of white-brined cheese and consequent proliferation of yeasts might lead to quality defects such as off-flavors, discoloration, blowing, etc. Nevertheless, it can be foreseen that not all yeast species will be able to proliferate during the maturation and storage of white-brined cheeses. The mapping of yeast hotspot areas in dairy production, as well as species identification, can be beneficially used by dairies to eliminate cross contamination and prevent spoilage of the products. Additional studies are required to evaluate the ability of yeasts to survive and propagate in the final product and their effect on cheese quality and shelf-life.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms10061079/s1, Table S1. Fermentation and assimilation of carbohydrates by Kluyveromyces spp. isolates from production line of white-brined cheese.
Author Contributions
Conceptualization, L.J., N.L. and A.G.; methodology, N.L. and A.G.; formal analysis, A.G.; investigation, A.G.; data curation, A.G.; writing—original draft preparation, A.G.; writing—review and editing, N.L., S.K.L. and L.J.; supervision, L.J. and N.L.; project administration, L.J. and N.L.; funding acquisition, L.J. and N.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Danish Dairy Research Foundation (project “Improve Dairy Life”); and the TALENT Doctoral Fellowship Program (funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No. 801199).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Geronikou, A.; Srimahaeak, T.; Rantsiou, K.; Triantafillidis, G.; Larsen, N.; Jespersen, L. Occurrence of Yeasts in White-Brined Cheeses: Methodologies for Identification, Spoilage Potential and Good Manufacturing Practices. Front. Microbiol. 2020, 11, 582778. [Google Scholar] [CrossRef] [PubMed]
- Salameh, C.; Banon, S.; Hosri, C.; Scher, J. An Overview of Recent Studies on the Main Traditional Fermented Milks and White Cheeses in the Mediterranean Region. Food Rev. Int. 2016, 32, 256–279. [Google Scholar] [CrossRef]
- Hayaloglu, A.A. Cheese Varieties Ripened Under Brine. In Cheese; Elsevier Ltd.: Malatya, Turkey, 2017; pp. 997–1040. [Google Scholar] [CrossRef]
- Šuranská, H.; Raspor, P.; Uroić, K.; Golić, N.; Kos, B.; Mihajlović, S.; Begović, J.; Šušković, J.; Topisirović, L.; Čadež, N. Characterisation of the Yeast and Mould Biota in Traditional White Pickled Cheeses by Culture-Dependent and Independent Molecular Techniques. Folia Microbiol. 2016, 61, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, M.R.; Fernández-Otero, C.; Rodríguez-Alonso, P.; Fernández-No, I.C.; Garabal, J.I.; Centeno, J.A. Characterization of Yeasts Isolated from Artisanal Short-Ripened Cows’ Cheeses Produced in Galicia (NW Spain). Food Microbiol. 2016, 53, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Buehler, A.J.; Evanowski, R.L.; Martin, N.H.; Boor, K.J.; Wiedmann, M. Internal Transcribed Spacer (ITS) Sequencing Reveals Considerable Fungal Diversity in Dairy Products. J. Dairy Sci. 2017, 100, 8814–8825. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, V.M.; Borelli, B.M.; Lara, C.A.; Soares, M.A.; Pataro, C.; Bodevan, E.C.; Rosa, C.A. The Influence of Seasons and Ripening Time on Yeast Communities of a Traditional Brazilian Cheese. Food Res. Int. 2015, 69, 331–340. [Google Scholar] [CrossRef]
- Fröhlich-Wyder, M.T.; Arias-Roth, E.; Jakob, E. Cheese Yeasts. Yeast 2019, 36, 129–141. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Mounier, J. Diversity and Control of Spoilage Fungi in Dairy Products: An Update. Microorganisms 2017, 5, 42. [Google Scholar] [CrossRef]
- Tokak, S.; Kiliç, İ.H.; Yalçin, H.T.; Duran, T. Detection of Extracellular Lipases and Genotypic Identification from Yeast Causing Spoilage of Some Dairy Products Produced in Gaziantep. KSU J. Agric. Nat. 2019, 22, 207–212. [Google Scholar] [CrossRef]
- Banjara, N.; Suhr, M.J.; Hallen-Adams, H.E. Diversity of Yeast and Mold Species from a Variety of Cheese Types. Curr. Microbiol. 2015, 70, 792–800. [Google Scholar] [CrossRef]
- Gardini, F.; Tofalo, R.; Belletti, N.; Iucci, L.; Suzzi, G.; Torriani, S.; Guerzoni, M.E.; Lanciotti, R. Characterization of Yeasts Involved in the Ripening of Pecorino Crotonese Cheese. Food Microbiol. 2006, 23, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Brandl, H.; Fricker-Feer, C.; Ziegler, D.; Mandal, J.; Stephan, R.; Lehner, A. Distribution and Identification of Culturable Airborne Microorganisms in a Swiss Milk Processing Facility. J. Dairy Sci. 2014, 97, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Stobnicka-Kupiec, A.; Gołofit-Szymczak, M.; Górny, R. Microbial Contamination Level and Microbial Diversity of Occupational Environment in Commercial and Traditional Dairy Plants. Ann. Agric. Environ. Med. 2019, 26, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Beletsiotis, E.; Ghikas, D.; Kalantzi, K. Incorporation of Microbiological and Molecular Methods in HACCP Monitoring Scheme of Molds and Yeasts in a Greek Dairy Plant: A Case Study. Procedia Food Sci. 2011, 1, 1051–1059. [Google Scholar] [CrossRef]
- Kandasamy, S.; Park, W.S.; Yoo, J.; Yun, J.; Kang, H.B.; Seol, K.H.; Oh, M.H.; Ham, J.S. Characterisation of Fungal Contamination Sources for Use in Quality Management of Cheese Production Farms in Korea. Asian-Australas. J. Anim. Sci. 2020, 33, 1002–1011. [Google Scholar] [CrossRef]
- Masotti, F.; Vallone, L.; Ranzini, S.; Silvetti, T.; Morandi, S.; Brasca, M. Effectiveness of Air Disinfection by Ozonation or Hydrogen Peroxide Aerosolization in Dairy Environments. Food Control 2019, 97, 32–38. [Google Scholar] [CrossRef]
- Masotti, F.; Cattaneo, S.; Stuknytė, M.; De Noni, I. Airborne Contamination in the Food Industry: An Update on Monitoring and Disinfection Techniques of Air. Trends Food Sci. Technol. 2019, 90, 147–156. [Google Scholar] [CrossRef]
- Van Der Aa Kühle, A.; Jespersen, L. The Taxonomic Position of Saccharomyces Boulardii as Evaluated by Sequence Analysis of the D1/D2 Domain of 26S RDNA, the ITS1-5.8S RDNA-ITS2 Region and the Mitochondrial Cytochrome-c Oxidase II Gene. Syst. Appl. Microbiol. 2003, 26, 564–571. [Google Scholar] [CrossRef]
- Naumova, E.S.; Naumov, G.I.; Nikitina, T.N.; Sadykova, A.Z.; Kondratieva, V.I. Molecular Genetic and Physiological Differentiation of Kluyveromyces Lactis and Kluyveromyces Marxianus: Analysis of Strains from the All-Russian Collection of Microorganisms (VKM). Microbiology 2012, 81, 216–223. [Google Scholar] [CrossRef]
- Petersen, K.M.; Moller, P.L.; Jespersen, L. DNA Typing Methods for Differentiation of Debaryomyces Hansenii Strains and Other Yeasts Related to Surface Ripened Cheeses. Int. J. Food Microbiol. 2001, 69, 11–24. [Google Scholar] [CrossRef]
- Jespersen, L.; Nielsen, D.S.; Hønholt, S.; Jakobsen, M. Occurrence and Diversity of Yeasts Involved in Fermentation of West African Cocoa Beans. FEMS Yeast Res. 2005, 5, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for Isolation, Phenotypic Characterization and Maintenance of Yeasts. In The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 87–110. [Google Scholar] [CrossRef]
- Pangallo, D.; Šaková, N.; Koreňová, J.; Puškárová, A.; Kraková, L.; Valík, L.; Kuchta, T. Microbial Diversity and Dynamics during the Production of May Bryndza Cheese. Int. J. Food Microbiol. 2014, 170, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Büchl, N.R.; Seiler, H. Yeasts and Molds: Yeasts in Milk and Dairy Products. Encyclopedia of Dairy Sciences, 2nd ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 744–753. [Google Scholar] [CrossRef]
- Simon, X.; Duquenne, P. Assessment of Workers’ Exposure to Bioaerosols in a French Cheese Factory. Ann. Occup. Hyg. 2014, 58, 677–692. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salustiano, V.C.; Andrade, N.J.; Brandão, S.C.C.; Azeredo, R.M.C.; Lima, S.A.K. Microbiological Air Quality of Processing Areas in a Dairy Plant as Evaluated by the Sedimentation Technique and a One-Stage Air Sampler. Braz. J. Microbiol. 2003, 34, 255–259. [Google Scholar] [CrossRef]
- Luck, H.; Gavron, H. Quality Control in the Dairy Industry. In Dairy Microbiology—The Microbiology of Milk Products; Robinson, R.K., Ed.; Elsevier Applied Science: London, UK, 1990; pp. 345–392. [Google Scholar]
- Mostert, J.; Jooste, P. Quality Control in the Dairy Industry. In Dairy Microbiology Handbook—The Microbiology of Milk and Milk Products; Robinson, R.K., Ed.; John Wiley and Sons: New York, NY, USA, 2002; pp. 655–736. [Google Scholar]
- Sveum, W.H.; Moberg, L.J.; Rude, R.; Frank, J.F. Microbiological Monitoring of the Food Processing Environment in Compendium of Methods for the Microbiological Examination of Foods. In Compendium of Methods for the Microbiological Examination of Foods; Vanderzant, C., Splittstoeser, D.F., Eds.; APHA: Washington, DC, USA, 1992; pp. 51–75. [Google Scholar]
- Shale, K.; Lues, J.F.R. The Etiology of Bioaerosols in Food Environments. Food Rev. Int. 2007, 23, 73–90. [Google Scholar] [CrossRef]
- Lachance, M.A. Kluyveromyces van Der Walt. In The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011; Volume 2, pp. 471–481. [Google Scholar] [CrossRef]
- Rantsiou, K.; Urso, R.; Dolci, P.; Comi, G.; Cocolin, L. Microflora of Feta Cheese from Four Greek Manufacturers. Int. J. Food Microbiol. 2008, 126, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Chipilev, N.; Daskalov, H.; Stoyanchev, T. Study on the Prevalence of Lipolytic Yeasts and Moulds in Raw Cow Milk and White Brined Cheese. Bulg. J. Vet. Med. 2016, 19, 117–126. [Google Scholar] [CrossRef]
- Togay, S.O.; Capece, A.; Siesto, G.; Aksu, H.; Altunatmaz, S.S.; Aksu, F.Y.; Romano, P.; Yuceer, Y.K. Molecular Characterization of Yeasts Isolated from Traditional Turkish Cheeses. Food Sci. Technol. 2020, 40, 871–876. [Google Scholar] [CrossRef]
- Lavoie, K.; Touchette, M.; St-Gelais, D.; Labrie, S. Characterization of the Fungal Microflora in Raw Milk and Specialty Cheeses of the Province of Quebec. Dairy Sci. Technol. 2012, 92, 455–468. [Google Scholar] [CrossRef]
- Andrade, R.P.; Melo, C.N.; Genisheva, Z.; Schwan, R.F.; Duarte, W.F. Yeasts from Canastra Cheese Production Process: Isolation and Evaluation of Their Potential for Cheese Whey Fermentation. Food Res. Int. 2017, 91, 72–79. [Google Scholar] [CrossRef]
- Haastrup, M.K.; Johansen, P.; Malskær, A.H.; Castro-Mejía, J.L.; Kot, W.; Krych, L.; Arneborg, N.; Jespersen, L. Cheese Brines from Danish Dairies Reveal a Complex Microbiota Comprising Several Halotolerant Bacteria and Yeasts. Int. J. Food Microbiol. 2018, 285, 173–187. [Google Scholar] [CrossRef] [PubMed]
- Delavenne, E.; Mounier, J.; Asmani, K.; Jany, J.L.; Barbier, G.; Le Blay, G. Fungal Diversity in Cow, Goat and Ewe Milk. Int. J. Food Microbiol. 2011, 151, 247–251. [Google Scholar] [CrossRef] [PubMed]
- von Neubeck, M.; Baur, C.; Krewinkel, M.; Stoeckel, M.; Kranz, B.; Stressler, T.; Fischer, L.; Hinrichs, J.; Scherer, S.; Wenning, M. Biodiversity of Refrigerated Raw Milk Microbiota and Their Enzymatic Spoilage Potential. Int. J. Food Microbiol. 2015, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.R.; Chow, N.; Forsberg, K.; Litvintseva, A.P.; Lockhart, S.R.; Welsh, R.; Vallabhaneni, S.; Chiller, T. On the Origins of a Species: What Might Explain the Rise of Candida Auris? J. Fungi 2019, 5, 58. [Google Scholar] [CrossRef]
- Spivak, E.S.; Hanson, K.E. Candida auris: An Emerging Fungal Pathogen. J. Clin. Microbiol. 2018, 56, e01588-17. [Google Scholar] [CrossRef]
- Borelli, B.M.; Ferreira, E.G.; Lacerda, I.C.A.; Franco, G.R.; Rosa, C.A. Yeast Populations Associated with the Artisanal Cheese Produced in the Region of Serra Da Canastra, Brazil. World J. Microbiol. Biotechnol. 2006, 22, 1115–1119. [Google Scholar] [CrossRef]
- Trofa, D.; Gácser, A.; Nosanchuk, J.D. Candida Parapsilosis, an Emerging Fungal Pathogen. Clin. Microbiol. Rev. 2008, 21, 606–625. [Google Scholar] [CrossRef]
- Lima, G.B.L.; Rosa, C.A.; Johann, S.; De Lourdes Almeida Vieira, M.; De Cássia Oliveira Gomes, F. Yeasts Isolated from Tropical Fruit Ice Creams: Diversity, Antifungal Susceptibility and Adherence to Buccal Epithelial Cells. Braz. J. Food Technol. 2019, 22, e2018197. [Google Scholar] [CrossRef]
- Park, H.J.; Bae, J.H.; Ko, H.J.; Lee, S.H.; Sung, B.H.; Han, J.I.; Sohn, J.H. Low-PH Production of d-Lactic Acid Using Newly Isolated Acid Tolerant Yeast Pichia Kudriavzevii NG7. Biotechnol. Bioeng. 2018, 115, 2232–2242. [Google Scholar] [CrossRef]
- Johansen, P.G.; Owusu-Kwarteng, J.; Parkouda, C.; Padonou, S.W.; Jespersen, L. Occurrence and Importance of Yeasts in Indigenous Fermented Food and Beverages Produced in Sub-Saharan Africa. Front. Microbiol. 2019, 10, 1789. [Google Scholar] [CrossRef]
- Padilla, B.; Belloch, C.; López-Díez, J.J.; Flores, M.; Manzanares, P. Potential Impact of Dairy Yeasts on the Typical Flavour of Traditional Ewes’ and Goats’ Cheeses. Int. Dairy J. 2014, 35, 122–129. [Google Scholar] [CrossRef]
- Vasdinyei, R.; Deák, T. Characterization of Yeast Isolates Originating from Hungarian Dairy Products Using Traditional and Molecular Identification Techniques. Int. J. Food Microbiol. 2003, 86, 123–130. [Google Scholar] [CrossRef]
- Bai, M.; Qing, M.; Guo, Z.; Zhang, Y.; Chen, X.; Bao, Q.; Zhang, H.; Sun, T.S. Occurrence and Dominance of Yeast Species in Naturally Fermented Milk from the Tibetan Plateau of China. Can. J. Microbiol. 2010, 56, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D.; Liu, J.L.; Jiang, T.M.; Li, L.; Fang, G.Z.; Liu, Y.P.; Chen, L.J. Influence of Kluyveromyces Marxianus on Proteins, Peptides, and Amino Acids in Lactobacillus-Fermented Milk. Food Sci. Biotechnol. 2017, 26, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Sádecká, J.; Šaková, N.; Pangallo, D.; Koreňová, J.; Kolek, E.; Puškárová, A.; Bučková, M.; Valík, L.; Kuchta, T. Microbial Diversity and Volatile Odour-Active Compounds of Barrelled Ewes’ Cheese as an Intermediate Product That Determines the Quality of Winter Bryndza Cheese. LWT—Food Sci. Technol. 2016, 70, 237–244. [Google Scholar] [CrossRef]
- Gonçalves Dos Santos, M.T.P.; Benito, M.J.; de Guía Córdoba, M.; Alvarenga, N.; de Herrera, S.R.-M.S. Yeast Community in Traditional Portuguese Serpa Cheese by Culture-Dependent and -Independent DNA Approaches. Int. J. Food Microbiol. 2017, 262, 63–70. [Google Scholar] [CrossRef]
- Aponte, M.; Pepe, O.; Blaiotta, G. Short Communication: Identification and Technological Characterization of Yeast Strains Isolated from Samples of Water Buffalo Mozzarella Cheese. J. Dairy Sci. 2010, 93, 2358–2361. [Google Scholar] [CrossRef]
- Liu, X.Z.; Wang, Q.M.; Göker, M.; Groenewald, M.; Kachalkin, A.V.; Lumbsch, H.T.; Millanes, A.M.; Wedin, M.; Yurkov, A.M.; Boekhout, T.; et al. Towards an Integrated Phylogenetic Classification of the Tremellomycetes. Stud. Mycol. 2015, 81, 85–147. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Deniel, F.; Coton, E.; Mounier, J. Diversity of Spoilage Fungi Associated with Various French Dairy Products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef]
- Prakash, A.; Randhawa, H.S.; Khan, Z.U.; Ahmad, S.; Hagen, F.; Meis, J.F.; Chowdhary, A. Environmental Distribution of Cryptococcus Species and Some Other Yeast-like Fungi in India. Mycoses 2018, 61, 305–313. [Google Scholar] [CrossRef]
- de Oliveira Brito, M.; de Souza Bessa, M.A.; de Paula Menezes, R.; de Brito Röder, D.V.D.; Penatti, M.P.A.; Pimenta, J.P.; de Aguiar, P.A.D.F.; Pedroso, R.D.S. Isolation of Cryptococcus Species from the External Environments of Hospital and Academic Areas. J. Infect. Dev. Ctries. 2019, 13, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Gostinčar, C.; Černoša, A.; Gunde-Cimerman, N. Stress-Tolerant Yeasts: Opportunistic Pathogenicity versus Biocontrol Potential. Genes 2019, 10, 42. [Google Scholar] [CrossRef] [PubMed]
- Kot, A.M.; Błażejak, S.; Kurcz, A.; Gientka, I.; Kieliszek, M. Rhodotorula Glutinis—Potential Source of Lipids, Carotenoids, and Enzymes for Use in Industries. Appl. Microbiol. Biotechnol. 2016, 100, 6103–6117. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Ghoshal, G. Optimization of Carotenoids Production by Rhodotorula Mucilaginosa (MTCC-1403) Using Agro-Industrial Waste in Bioreactor: A Statistical Approach. Biotechnol. Rep. 2020, 25, e00407. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).