
Cellular Modifications of Rhodococci Exposed to Separate and Combined Effects of Pharmaceutical Pollutants

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains
2.3. Culture Conditions
2.4. Respirometry
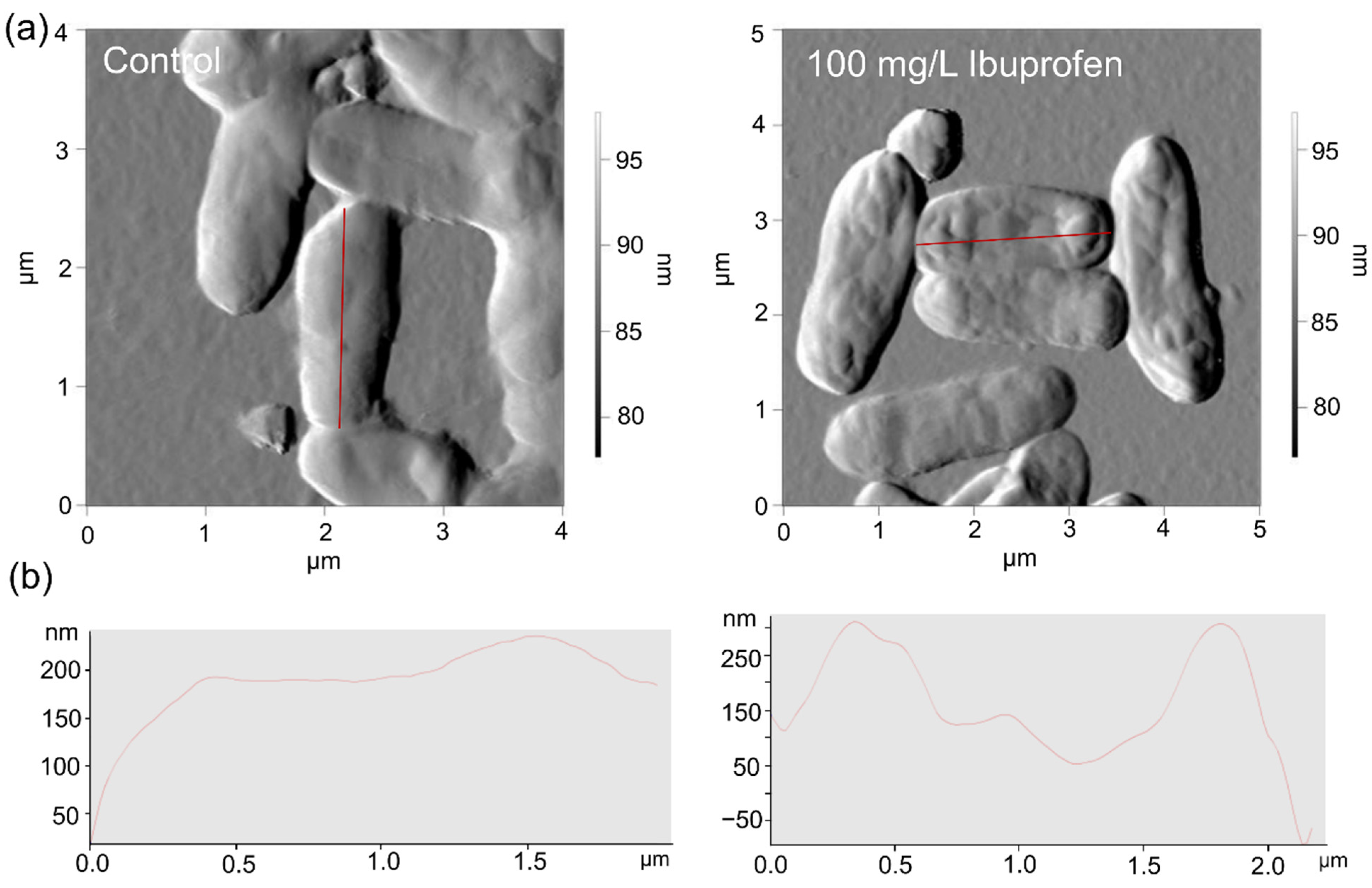
2.5. Microscopy
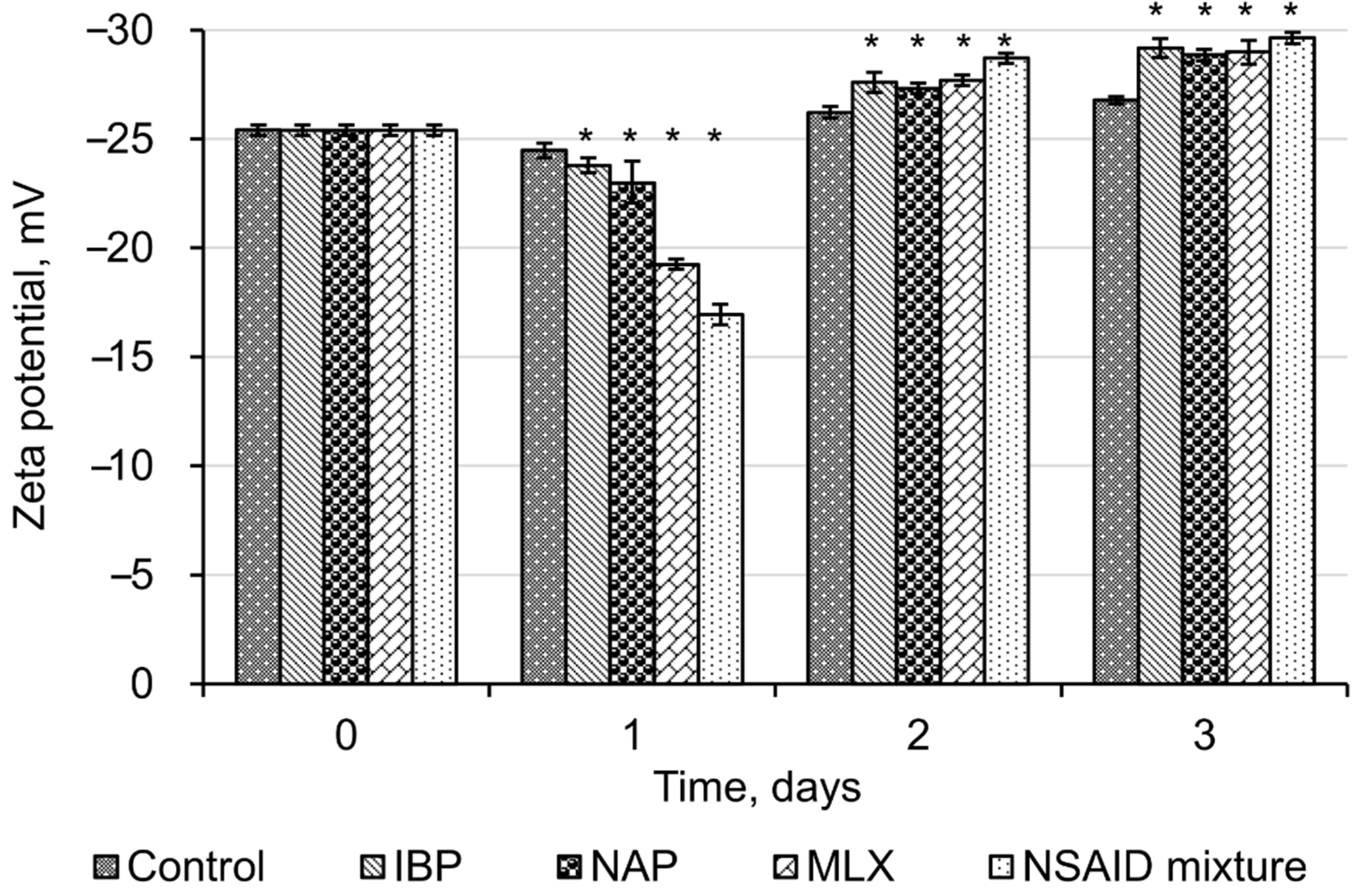
2.6. Zeta Potential
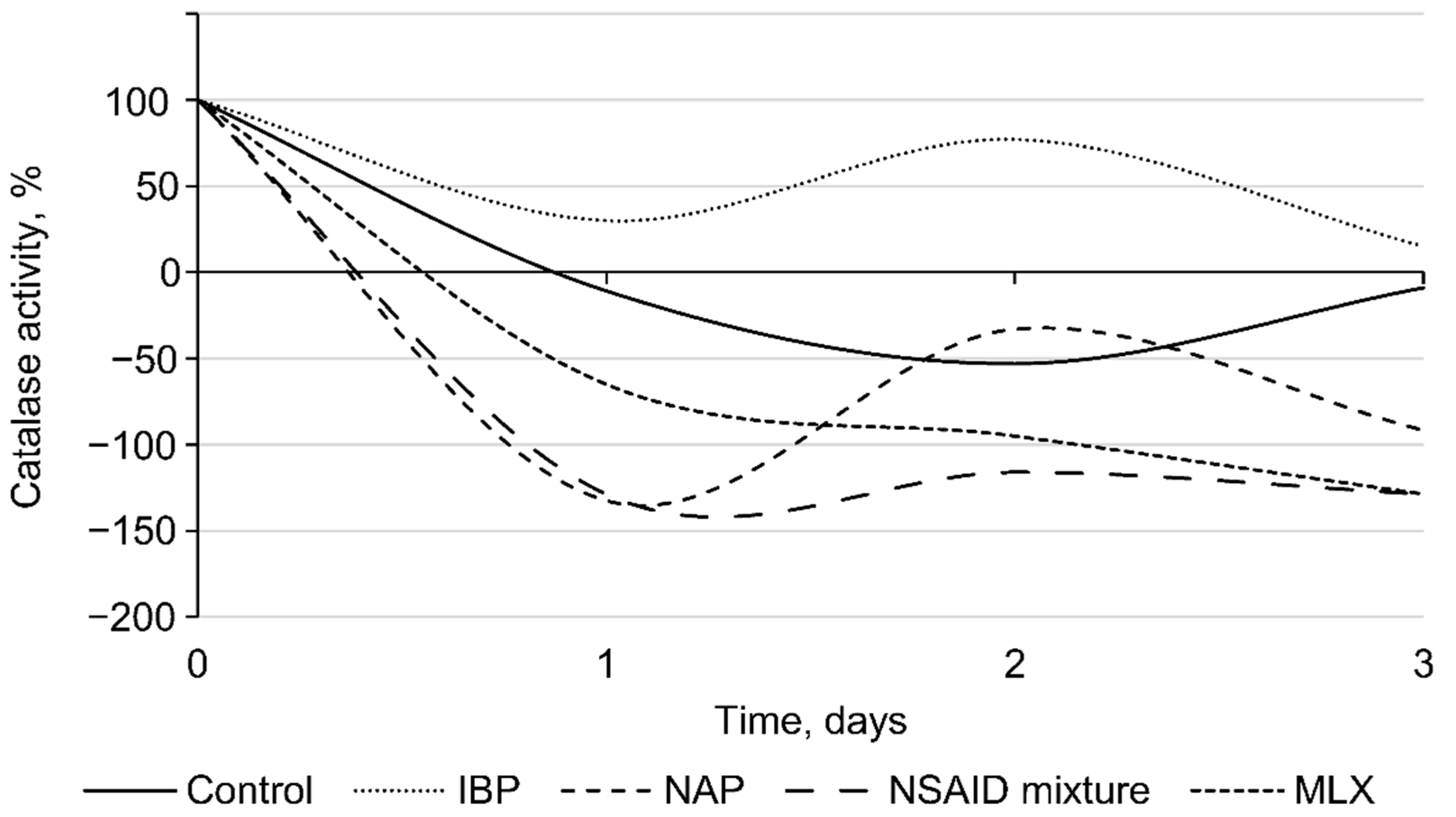
2.7. Catalase Activity
2.8. Analytical Methods
3. Results and Discussion
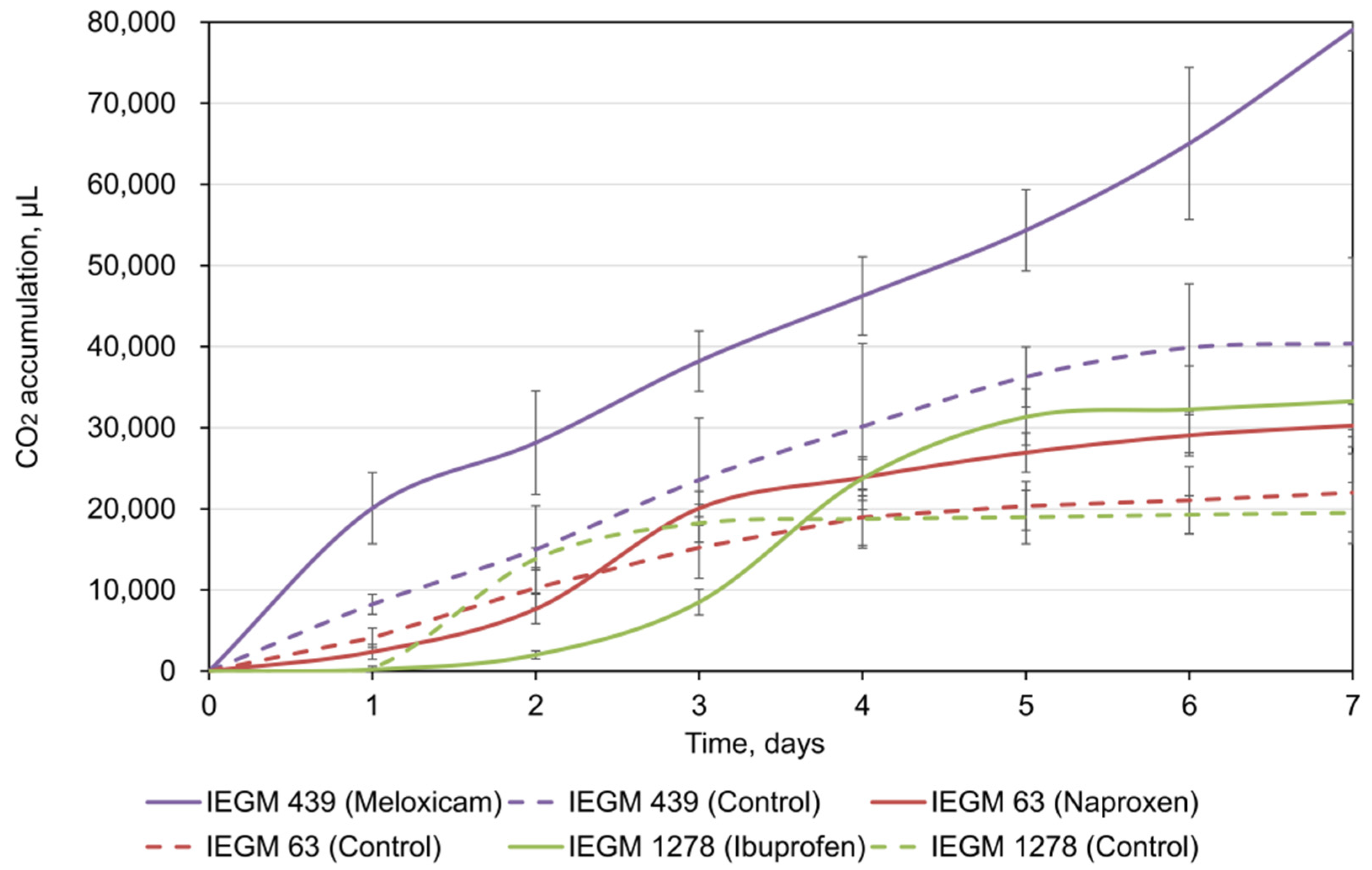
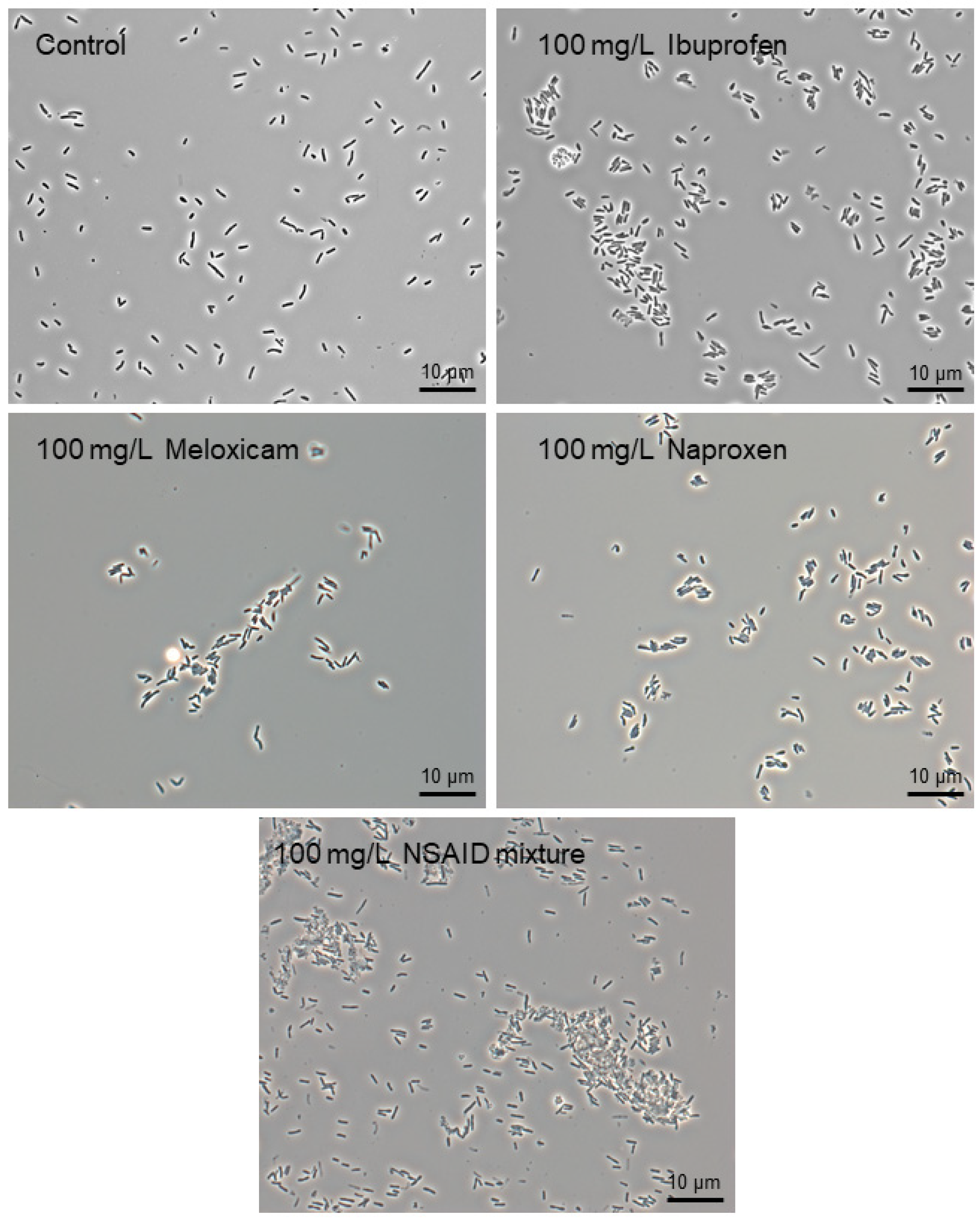
3.1. Biodegradation of Individual NSAIDs and Induced Morphological Changes of Biodegrading Strains
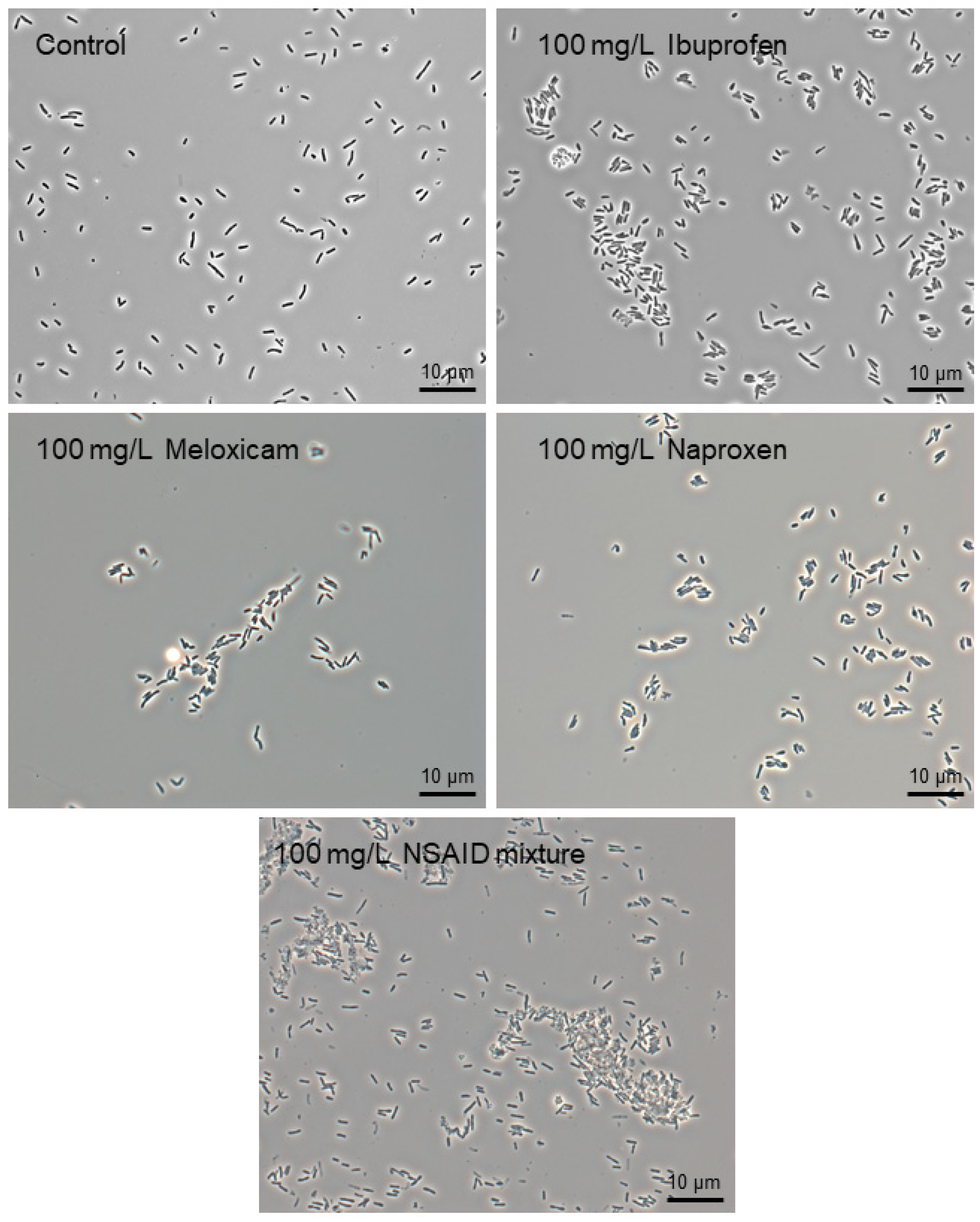
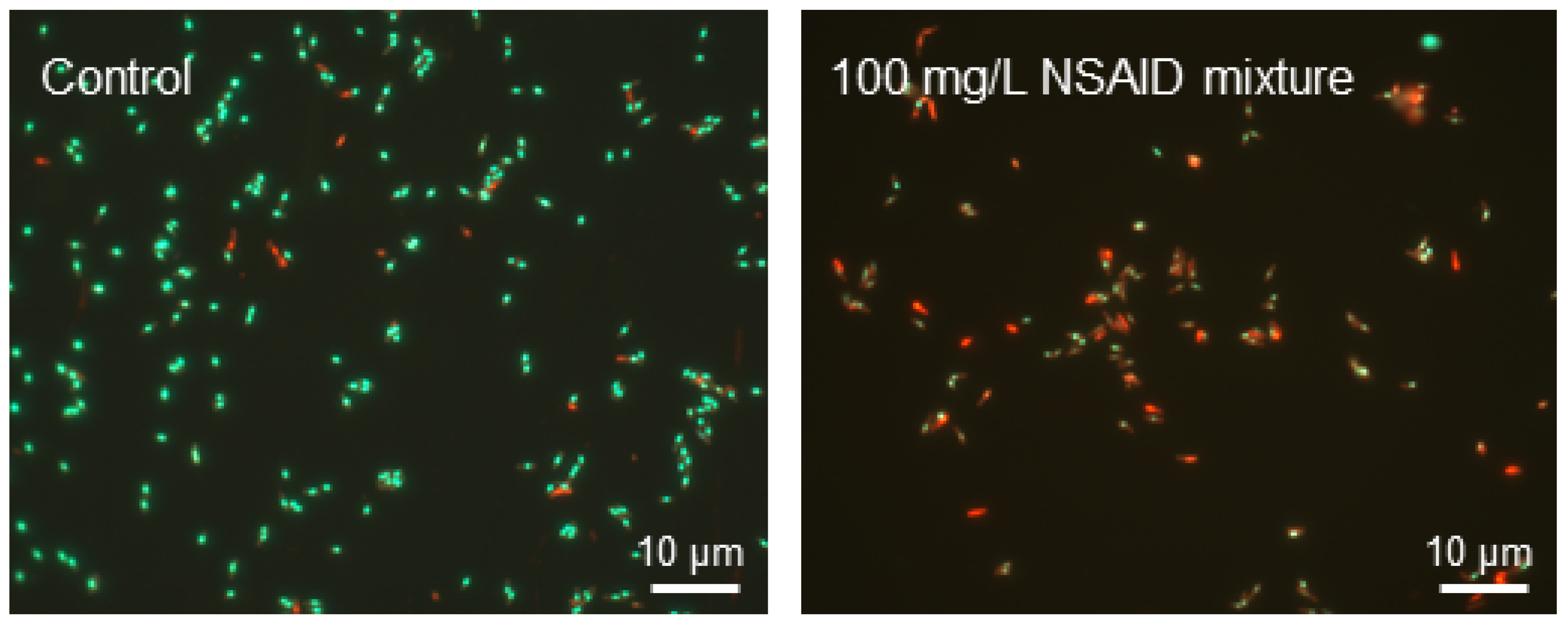
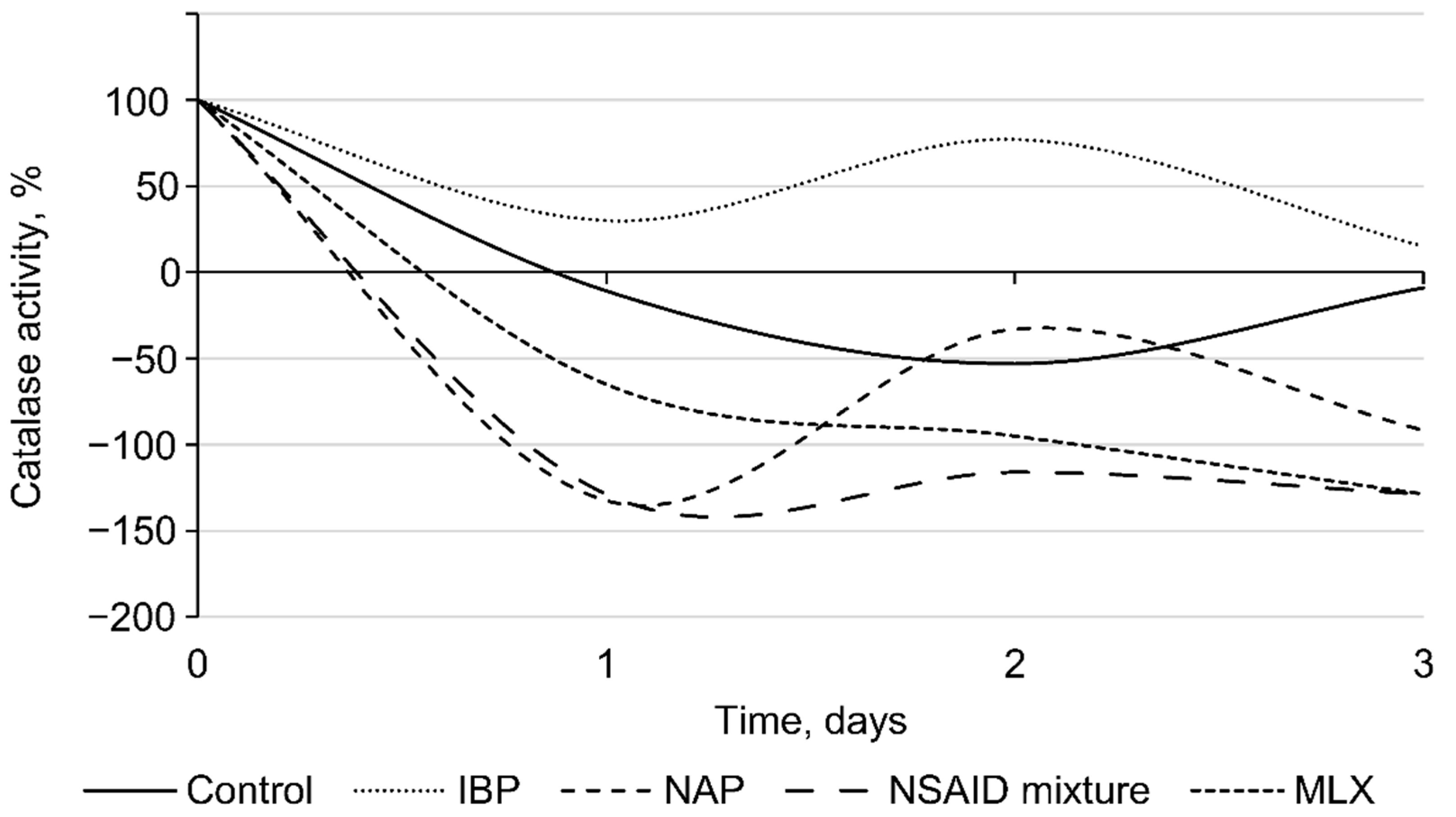
3.2. Effects of Individual NSAIDs and Their Mixture on Rhodococcus cerastii IEGM 1278 Cells
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- aus der Beek, T.; Weber, F.A.; Bergmann, A.; Hickmann, S.; Ebert, I.; Hein, A.; Küster, A. Pharmaceuticals in the environment—Global occurrences and perspectives. Environ. Toxicol. Chem. 2016, 35, 823–835. [Google Scholar] [CrossRef]
- Brodin, T.; Fick, J.; Jonsson, M.; Klaminder, J. Dilute concentrations of a psychiatric drug alter behavior of fish from natural populations. Science 2013, 339, 814–815. [Google Scholar] [CrossRef] [PubMed]
- Izadi, P.; Izadi, P.; Salem, R.; Papry, S.A.; Magdouli, S.; Pulicharla, R.; Brar, S.K. Non-steroidal anti-inflammatory drugs in the environment: Where were we and how far we have come? Environ. Pollut. 2020, 267, 115370. [Google Scholar] [CrossRef]
- Horký, P.; Grabic, R.; Grabicová, K.; Brooks, B.W.; Douda, K.; Slavik, O.; Hubená, P.; Santos, E.M.S.; Randák, T. Methamphetamine pollution elicits addiction in wild fish. J. Exp. Biol. 2021, 224, jeb242145. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jin, B.; Han, M.; Yu, Y.; Zhang, G.; Arp, H.P.H. The distribution of persistent, mobile and toxic (PMT) pharmaceuticals and personal care products monitored across Chinese water resources. J. Hazard. Mater. Lett. 2021, 2, 100026. [Google Scholar] [CrossRef]
- Patel, M.; Kumar, R.; Kishor, K.; Misna, T.; Pittman, C.U.; Mohan, D. Pharmaceuticals of emerging concern in aquatic systems: Chemistry, occurrence, effects, and removal methods. Chem. Rev. 2019, 119, 3510–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushik, G.; Thomas, M.A. The potential association of psychoactive pharmaceuticals in the environment with human neurological disorders. Sustain. Chem. Pharm. 2019, 13, 100148. [Google Scholar] [CrossRef]
- Tyumina, E.A.; Bazhutin, G.A.; Cartagena Gómez, A.d.P.; Ivshina, I.B. Nonsteroidal anti-inflammatory drugs as emerging contaminants. Microbiology 2020, 89, 148–163. [Google Scholar] [CrossRef]
- Wojcieszyńska, D.; Guzik, H.; Guzik, U. Non-steroidal anti-inflammatory drugs in the era of the COVID-19 pandemic in the context of the human and the environment. Sci. Total Environ. 2022, 834, 155317. [Google Scholar] [CrossRef]
- Wilkinson, J.L.; Boxall, A.B.A.; Kolpin, D.W.; Leung, K.M.Y.; Lai, R.W.S.; Galban-Malag, C.; Adell, A.D.; Mondon, J.; Metian, M.; Marchant, R.A.; et al. Pharmaceutical pollution of the world’s rivers. Proc. Natl. Acad. Sci. USA 2022, 119, e2113947119. [Google Scholar] [CrossRef]
- Barra Caracciolo, A.; Topp, E.; Grenni, P. Pharmaceuticals in the environment: Biodegradation and effects on natural microbial communities. A review. J. Pharm. Biomed. Anal. 2015, 106, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, N.K.; Ramesh, A.; Pakshirajan, K. Bacterial degradation of aromatic xenobiotic compounds: An overview on metabolic pathways and molecular approaches. In Microorganisms in Environmental Management: Microbes and Environment; Satyanarayana, T., Narain Johri, B., Prakash, A., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 201–220. [Google Scholar] [CrossRef]
- Hoostal, M.J.; Gabriela Bidart-Bouzat, M.; Bouzat, J.L. Local adaptation of microbial communities to heavy metal stress in polluted sediments of Lake Erie. FEMS Microbiol. Ecol. 2008, 65, 156–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruberto, L.A.M.; Vazquez, S.; Lobalbo, A.; Mac Cormack, W.P. Psychrotolerant hydrocarbon-degrading Rhodococcus strains isolated from polluted Antarctic soils. Antarct. Sci. 2005, 17, 47–56. [Google Scholar] [CrossRef]
- Song, X.; Xu, Y.; Li, G.; Zhang, Y.; Huang, T.; Hu, Z. Isolation, characterization of Rhodococcus sp. P14 capable of degrading high-molecular-weight polycyclic aromatic hydrocarbons and aliphatic hydrocarbons. Mar. Pollut. Bull. 2011, 62, 2122–2128. [Google Scholar] [CrossRef]
- Yang, C.W.; Liu, C.; Chang, B.V. Biodegradation of amoxicillin, tetracyclines and sulfonamides in wastewater sludge. Water 2020, 12, 2147. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Ivshina, I.B. Bioremediation of contaminated environments using Rhodococcus. In Biology of Rhodococcus; Alvarez, H.M., Ed.; Springer: Cham, Switzerland, 2019; pp. 231–270. [Google Scholar] [CrossRef]
- Lang, F.S.; Destain, J.; Delvigne, F.; Druart, P.; Ongena, M.; Thonart, P. Biodegradation of polycyclic aromatic hydrocarbons in mangrove sediments under different strategies: Natural attenuation, biostimulation, and bioaugmentation with Rhodococcus erythropolis T902.1. Water. Air. Soil Pollut. 2016, 227, 227. [Google Scholar] [CrossRef]
- Alvarez, H.M. Biology of Rhodococcus, 2nd ed.; Microbiology Monographs; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–386. [Google Scholar] [CrossRef]
- Krivoruchko, A.; Kuyukina, M.; Ivshina, I. Advanced Rhodococcus biocatalysts for environmental biotechnologies. Catalysts 2019, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Pátek, M.; Grulich, M.; Nešvera, J. Stress response in Rhodococcus strains. Biotechnol. Adv. 2021, 53, 107698. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Rychkova, M.I.; Vikhareva, E.V.; Chekryshkina, L.A.; Mishenina, I.I. Catalysis of the biodegradation of unusable medicines by alkanotrophic rhodococci. Appl. Biochem. Microbiol. 2006, 42, 392–395. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Vikhareva, E.V.; Richkova, M.I.; Mukhutdinova, A.N.; Karpenko, J.N. Biodegradation of drotaverine hydrochloride by free and immobilized cells of Rhodococcus rhodochrous IEGM 608. World J. Microbiol. Biotechnol. 2012, 28, 2997–3006. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Mukhutdinova, A.N.; Tyumina, H.A.; Vikhareva, H.V.; Suzina, N.E.; El’-Registan, G.I.; Mulyukin, A.L. Drotaverine hydrochloride degradation using cyst-like dormant cells of Rhodococcus ruber. Curr. Microbiol. 2015, 70, 307–314. [Google Scholar] [CrossRef]
- Khrenkov, A.N.; Vikhareva, E.V.; Tumilovich, E.Y.; Karpenko, Y.N.; Selyaninov, A.A.; Tyumina, E.A. Chromatographic analysis of acetylsalicylic acid in Rhodococcus cultural fluids. Moscow Univ. Chem. Bull. 2020, 75, 309–314. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Tyumina, E.A.; Kuzmina, M.V.; Vikhareva, E.V. Features of diclofenac biodegradation by Rhodococcus ruber IEGM 346. Sci. Rep. 2019, 9, 9159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazhutin, G.A.; Polygalov, M.A.; Tyumina, E.A.; Tyan, S.M.; Ivshina, I.B. Cometabolic bioconversion of ketoprofen by Rhodococcus erythropolis IEGM 746. In Science and Global Challenges of the 21st Century–Science and Technology: Proceedings of the International Perm Forum "Science and Global Challenges of the 21st Century"; Rocha, A., Isaeva, E., Eds.; Springer: Cham, Switzerland, 2022; pp. 404–410. [Google Scholar] [CrossRef]
- Ivshina, I.B.; Tyumina, E.A.; Bazhutin, G.A.; Vikhareva, E.V. Response of Rhodococcus cerastii IEGM 1278 to toxic effects of ibuprofen. PLoS ONE 2021, 16, e0260032. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Tyumina, E.A.; Bazhutin, G.A.; Vikhareva, E.V. Biodegrader of Ibuprofen. RF Patent 2762007, 25 November 2021. Priority of Invention of 25 May 2021. Bulletin of Inventions No. 35. Registered with the Public Register for Inventions, 14 December 2021. [Google Scholar]
- Catalogue of Strains of Regional Specialized Collection of Alkanotrophic Microorganisms. Available online: http://www.iegmcol/strains/index.html (accessed on 27 April 2022).
- Kuyukina, M.S.; Ivshina, I.B.; Korshunova, I.O.; Rubtsova, E.V. Assessment of bacterial resistance to organic solvents using a combined confocal laser scanning and atomic force microscopy (CLSM/AFM). J. Microbiol. Methods 2014, 107, 23–29. [Google Scholar] [CrossRef]
- Neumann, G.; Veeranagouda, Y.; Karegoudar, T.B.; Sahin, Ö.; Mäusezahl, I.; Kabelitz, N.; Kappelmeyer, U.; Heipieper, H.J. Cells of Pseudomonas putida and Enterobacter sp. adapt to toxic organic compounds by increasing their size. Extremophiles 2005, 9, 163–168. [Google Scholar] [CrossRef]
- Gogoleva, O.A.; Nemtseva, N.V.; Bukharin, O.V. Catalase activity of hydrocarbon-oxidizing bacteria. Appl. Biochem. Microbiol. 2012, 48, 552–556. [Google Scholar] [CrossRef]
- Kuyukina, M.S.; Rubtsova, E.V.; Ivshina, I.B.; Ivanov, R.V.; Lozinsky, V.I. Selective adsorption of hydrocarbon-oxidizing Rhodococcus cells in a column with hydrophobized poly(acrylamide) cryogel. J. Microbiol. Methods 2009, 79, 76–81. [Google Scholar] [CrossRef]
- Ojkic, N.; Serbanescu, D.; Banerjee, S. Surface-to-volume scaling and aspect ratio preservation in rod-shaped bacteria. Elife 2019, 8, e47033. [Google Scholar] [CrossRef]
- Harris, L.K.; Theriot, J.A. Surface area to volume ratio: A natural variable for bacterial morphogenesis. Trends Microbiol. 2018, 26, 815–832. [Google Scholar] [CrossRef] [Green Version]
- Lagadinou, M.; Onisor, M.O.; Rigas, A.; Musetescu, D.V.; Gkentzi, D.; Assimakopoulos, S.F.; Panos, G.; Marangos, M. Antimicrobial properties on non-antibiotic drugs in the era of increased bacterial resistance. Antibiotics 2020, 9, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheremnykh, K.M.; Luchnikova, N.A.; Grishko, V.V.; Ivshina, I.B. Bioconversion of ecotoxic dehydroabietic acid using Rhodococcus actinobacteria. J. Hazard. Mater. 2018, 346, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Kragh, K.N.; Hutchison, J.B.; Melaugh, G.; Rodesney, C.; Roberts, A.E.L.; Irie, Y.; Jensen, P.; Diggle, S.P.; Allen, R.J.; Gordon, V.; et al. Role of multicellular aggregates in biofilm formation. MBio 2016, 7, e00237-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klebensberger, J.; Lautenschlager, K.; Bressler, D.; Wingender, J.; Philipp, B. Detergent-induced cell aggregation in subpopulations of Pseudomonas aeruginosa as a preadaptive survival strategy. Environ. Microbiol. 2007, 9, 2247–2259. [Google Scholar] [CrossRef] [Green Version]
- Farrell, A.; Quilty, B. Substrate-dependent autoaggregation of Pseudomonas putida CP1 during the degradation of mono-chlorophenols and phenol. J. Ind. Microbiol. Biotechnol. 2002, 28, 316–324. [Google Scholar] [CrossRef]
- Pal, A.; Bhattacharjee, S.; Saha, J.; Sarkar, M.; Mandal, P. Bacterial survival strategies and responses under heavy metal stress: A comprehensive overview. Crit. Rev. Microbiol. 2021, 48, 327–355. [Google Scholar] [CrossRef]
- Cai, Y.M. Non-surface attached bacterial aggregates: A ubiquitous third lifestyle. Front. Microbiol. 2020, 11, 3106. [Google Scholar] [CrossRef]
- Burel, C.; Dreyfus, R.; Purevdorj-Gage, L. Physical mechanisms driving the reversible aggregation of Staphylococcus aureus and response to antimicrobials. Sci. Rep. 2021, 11, 15048. [Google Scholar] [CrossRef]
- Wilson, W.W.; Wade, M.M.; Holman, S.C.; Champlin, F.R. Status of methods for assessing bacterial cell surface charge properties based on zeta potential measurements. J. Microbiol. Methods 2001, 43, 153–164. [Google Scholar] [CrossRef]
- Benarroch, J.M.; Asally, M. The microbiologist’s guide to membrane potential dynamics. Trends Microbiol. 2020, 28, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, M.E.; Becker, E.C.; Choe, S. MstX and a putative potassium channel facilitate biofilm formation in Bacillus subtilis. PLoS ONE 2013, 8, e60993. [Google Scholar] [CrossRef] [Green Version]
- Prindle, A.; Liu, J.; Asally, M.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Ion channels enable electrical communication in bacterial communities. Nature 2015, 527, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Prindle, A.; Humphries, J.; Gabalda-Sagarra, M.; Asally, M.; Lee, D.Y.D.; Ly, S.; Garcia-Ojalvo, J.; Süel, G.M. Metabolic Co-dependence gives rise to collective oscillations within biofilms. Nature 2015, 523, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Hagge, S.O.; Hammer, M.U.; Wiese, A.; Seydel, U.; Gutsmann, T. Calcium adsorption and displacement: Characterization of lipid monolayers and their interaction with membrane-active peptides/proteins. BMC Biochem. 2006, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Halder, S.; Yadav, K.K.; Sarkar, R.; Mukherjee, S.; Saha, P.; Haldar, S.; Karmakar, S.; Sen, T. Alteration of zeta potential and membrane permeability in bacteria: A study with cationic agents. SpringerPlus 2015, 4, 672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayala-Torres, C.; Hernández, N.; Galeano, A.; Novoa-Aponte, L.; Soto, C.Y. Zeta potential as a measure of the surface charge of mycobacterial cells. Ann. Microbiol. 2014, 64, 1189–1195. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | NSAID | Biodegradation, % |
|---|---|---|
| R. cerastii IEGM 1278 | 100 mg/L ibuprofen | 7.8 ± 3.97 |
| R. ruber IEGM 439 | 100 mg/L meloxicam | 99.3 ± 1.37 |
| R. rhodochrous IEGM 63 | 100 mg/L naproxen | 46.6 ± 3.45 |
| Strain | Variant | Length, μm | Width, μm | Volume (V), μm3 | Surface Area (SA), μm2 | SA/V, μm−1 |
|---|---|---|---|---|---|---|
| Ibuprofen | ||||||
| R. cerastii IEGM 1278 | Control | 3.7 ± 0.25 | 1.0 ± 0.07 | 2.9 ± 0.41 | 12.5 ± 1.19 | 4.6 ± 0.28 |
| 100 mg/L | 2.8 ± 0.29 ** | 1.1 ± 0.10 * | 2.6 ± 0.42 | 11.5 ± 1.12 * | 4.4 ± 0.28 | |
| Meloxicam | ||||||
| R. ruber IEGM 439 | Control | 3.3 ± 0.28 | 0.9 ± 0.10 | 1.9 ± 0.46 | 10.1 ± 0.46 | 5.3 ± 0.54 |
| 100 mg/L | 2.7 ± 0.18 * | 1.0 ± 0.10 * | 2.0 ± 0.45 | 9.8 ± 1.30 | 4.9 ± 0.39 | |
| Naproxen | ||||||
| R. rhodochrous IEGM 63 | Control | 1.9 ± 0.10 | 1.0 ± 0.07 | 1.4 ± 0.21 | 7.12 ± 0.68 | 5.3 ± 0.28 |
| 100 mg/L | 1.6 ± 0.17 * | 1.1 ± 0.06 ** | 1.6 ± 0.29 * | 7.7 ± 0.91 | 4.8 ± 0.31 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivshina, I.; Bazhutin, G.; Tyan, S.; Polygalov, M.; Subbotina, M.; Tyumina, E. Cellular Modifications of Rhodococci Exposed to Separate and Combined Effects of Pharmaceutical Pollutants. Microorganisms 2022, 10, 1101. https://doi.org/10.3390/microorganisms10061101
Ivshina I, Bazhutin G, Tyan S, Polygalov M, Subbotina M, Tyumina E. Cellular Modifications of Rhodococci Exposed to Separate and Combined Effects of Pharmaceutical Pollutants. Microorganisms. 2022; 10(6):1101. https://doi.org/10.3390/microorganisms10061101
Chicago/Turabian StyleIvshina, Irina, Grigory Bazhutin, Semyon Tyan, Maxim Polygalov, Maria Subbotina, and Elena Tyumina. 2022. "Cellular Modifications of Rhodococci Exposed to Separate and Combined Effects of Pharmaceutical Pollutants" Microorganisms 10, no. 6: 1101. https://doi.org/10.3390/microorganisms10061101
APA StyleIvshina, I., Bazhutin, G., Tyan, S., Polygalov, M., Subbotina, M., & Tyumina, E. (2022). Cellular Modifications of Rhodococci Exposed to Separate and Combined Effects of Pharmaceutical Pollutants. Microorganisms, 10(6), 1101. https://doi.org/10.3390/microorganisms10061101