

Effect of Trehalose and Lactose Treatments on the Freeze-Drying Resistance of Lactic Acid Bacteria in High-Density Culture

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Medium, and Growth Conditions
2.2. High-Density Culture of Strains
2.3. Cell Preparation and Freeze-Drying
2.3.1. Cell Preparation
2.3.2. Freeze-Drying
2.4. Determination of Freeze-Drying Survival Rate
2.5. Scanning Electron Microscopy
2.6. Intracellular Contents Determination
2.6.1. Extraction of Intracellular Contents
2.6.2. Measurement of Intracellular Sugars
2.7. Extracellular Polysaccharide Determination
2.7.1. Extraction of Extracellular Polysaccharides
2.7.2. Determination of Extracellular Polysaccharides
2.8. Statistical Analysis
3. Results
3.1. Determination of the Freeze-Drying Survival of LAB with Different Sugars as the Sole Carbon Source
3.2. Effects of Mixed Sugars on the Freeze-Drying Survival Rate of LAB
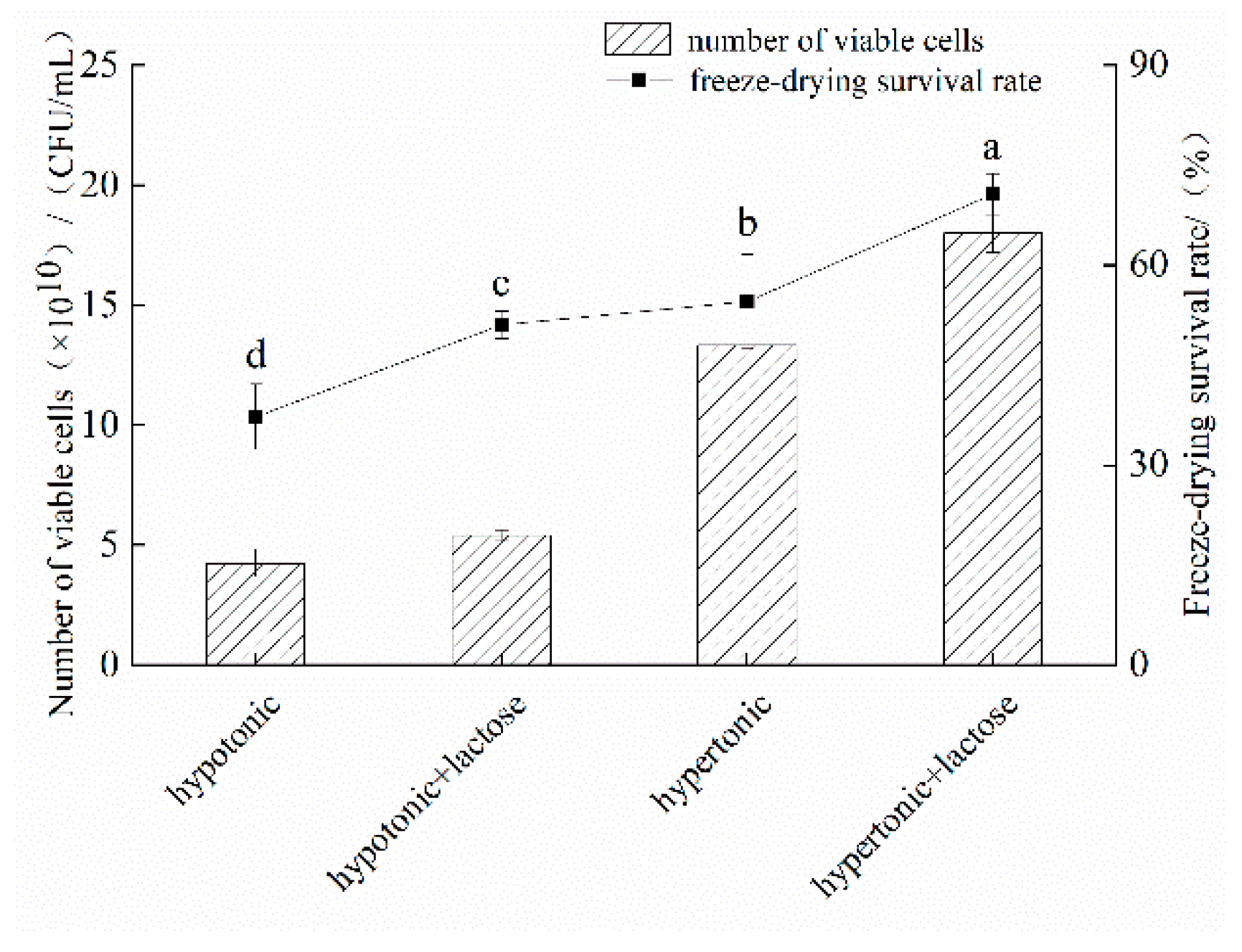
3.3. Effects of Trehalose Addition on the Freeze-Drying Resistance of L. fermentum fXJCJ6-1 in Different Conditions
3.4. Effects of Lactose Addition on the Freeze-Drying Resistance of L. brevis 173-1-2 in Different Conditions
3.5. Potential Mechanisms for the Increase in Freeze-Drying Survival of L. fermentum FXJCJ6-1 and L. reuteri CCFM1040 by Lactose Addition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Fatheree, N.Y.; Mangalat, N.; Rhoads, J.M. Human-derived probiotic Lactobacillus reuteri strains differentially reduce intestinal inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G1087–G1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segawa, S.; Nakakita, Y.; Takata, Y.; Wakita, Y.; Takafurni, K.A.; Hirotaka, K.A.; Wataria, J.; Yasui, H. Effect of oral administration of heat-killed Lactobacillus brevis SBC8803 on total and ovalbumin-specific immunoglobulin E production through the improvement of Th1/Th2 balance. Int. J. Food Microbiol. 2008, 121, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Santivarangkna, C.; Wenning, M.; Foerst, P.; Kulozik, U. Damage of cell envelope of Lactobacillus helveticus during vacuum drying. J. Appl. Microbiol. 2007, 102, 748–756. [Google Scholar] [CrossRef]
- Onwe, R.O.; Onwosi, C.O.; Ezugworie, F.N.; Ekwealor, C.C.; Okonkwo, C.C. Microbial trehalose boosts the ecological fitness of biocontrol agents, the viability of probiotics during long-term storage and plants tolerance to environmental-driven abiotic stress. Sci. Total Environ. 2022, 806, 150432. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, M.; Liu, Y.; Wang, Y.; Chen, Y.; Guan, W.; Li, X.; Wang, Y. Improved viability of Lactobacillus plantarum embedded in whey protein concentrate/pullulan/trehalose hydrogel during freeze drying. Carbohydr. Polym. 2021, 260, 117843. [Google Scholar] [CrossRef]
- Stefanello, R.F.; Nabeshima, E.H.; Iamanaka, B.T.; Ludwig, A.; Martins Fries, L.L.; Bernardi, A.O.; Copetti, M.V. Survival and stability of Lactobacillus fermentum and Wickerhamomyces anomalus strains upon lyophilisation with different cryoprotectant agents. Food Res. Int. 2019, 115, 90–94. [Google Scholar] [CrossRef]
- Crowe, J.H. Trehalose as a “chemical chaperone”: Fact and fantasy. In Molecular Aspects of the Stress Response: Chaperones, Membranes and Networks; Csermely, P., Vigh, L., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2007; Volume 594, pp. 143–158. [Google Scholar]
- Mensink, M.A.; Frijlink, H.W.; Van Der Voort Maarschalk, K.; Hinrichs, W.L.J. How sugars protect proteins in the solid state and during drying (review): Mechanisms of stabilization in relation to stress conditions. Eur. J. Pharm. Biopharm. 2017, 114, 288–295. [Google Scholar] [CrossRef]
- Finore, I.; Di Donato, P.; Mastascusa, V.; Nicolaus, B.; Poli, A. Fermentation Technologies for the Optimization of Marine Microbial Exopolysaccharide Production. Mar. Drugs 2014, 12, 3005–3024. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Kong, J.; Zhu, H.; Mao, B.; Cui, S.; Zhao, J. Lactobacillus, Bifidobacterium and Lactococcus response to environmental stress: Mechanisms and application of cross-protection to improve resistance against freeze-drying. J. Appl. Microbiol. 2022, 132, 802–821. [Google Scholar] [CrossRef]
- Schiraldi, C.; Adduci, V.; Valli, V.; Maresca, C.; Giuliano, M.; Lamberti, M.; Carteni, M.; De Rosa, M. High cell density cultivation of probiotics and lactic acid production. Biotechnol. Bioeng. 2003, 82, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.X.; Gu, L.; Zhang, J.; Wang, M.; Du, G.C.; Chen, J.; Li, H.Z. Optimisation for high cell density cultivation of Lactobacillus salivarius BBE 09-18 with response surface methodology. Int. Dairy J. 2014, 34, 230–236. [Google Scholar] [CrossRef]
- Cui, S.M.; Zhao, J.X.; Zhang, H.; Chen, W. High-density culture of Lactobacillus plantarum coupled with a lactic acid removal system with anion-exchange resins. Biochem. Eng. J. 2016, 115, 80–84. [Google Scholar] [CrossRef]
- Termont, S.; Vandenbroucke, K.; Iserentant, D.; Neirynck, S.; Steidler, L.; Remaut, E.; Rottiers, P. Intracellular accumulation of trehalose protects Lactococcus lactis from freeze-drying damage and bile toxicity and increases gastric acid resistance. Appl. Environ. Microbiol. 2006, 72, 7694–7700. [Google Scholar] [CrossRef] [Green Version]
- Phoon, P.Y.; Galindo, F.G.; Vicente, A.; Deimek, P. Pulsed electric field in combination with vacuum impregnation with trehalose improves the freezing tolerance of spinach leaves. J. Food Eng. 2008, 88, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Van, T. Correlation of Intracellular Trehalose Concentration with Desiccation Resistance of Soil Escherichia coli Populations. Appl. Environ. Microbiol. 2012, 78, 7407–7413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowe, J.H.; Crowe, L.M.; Carpenter, J.F.; Aurell Wistrom, C. Stabilization of dry phospholipid bilayers and proteins by sugars. Biochem. J. 1987, 242, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.P.; Lu, M.J.; Guo, H.F.; Li, W.; Zhang, H.P. Protective Effect of Sucrose on the Membrane Properties of Lactobacillus casei Zhang Subjected to Freeze-Drying. J. Food Prot. 2010, 73, 715–719. [Google Scholar] [CrossRef]
- Li, C.; Liu, L.B.; Liu, N. Effects of carbon sources and lipids on freeze-drying survival of Lactobacillus bulgaricus in growth media. Ann. Microbiol. 2012, 62, 949–956. [Google Scholar] [CrossRef]
- Marx, J.G.; Carpenter, S.D.; Deming, J.W. Production of cryoprotectant extracellular polysaccharide substances (EPS) by the marine psychrophilic bacterium Colwellia psychrerythraea strain 34H under extreme conditions. Can. J. Microbiol. 2009, 55, 63–72. [Google Scholar] [CrossRef]
- Gancel, F.; Novel, G. Exopolysaccharide production by Streptococcus-salivarius ssp. thermophilus cultures. 1. Conditions of production. J. Dairy Sci. 1994, 77, 685–688. [Google Scholar] [CrossRef]
- Cui, S.; Pan, Z.; Wu, S.; Mao, B.; Tang, X.; Zhang, Q.; Zhang, H.; Zhao, J. Improvement of the Lyophilization Survival Rate of Lactobacillus casei via Regulation of Its Surface Substances. Foods 2022, 11, 3468. [Google Scholar] [CrossRef] [PubMed]
- Duong, T.; Barrangou, R.; Russell, W.M.; Klaenhammer, T.R. Characterization of the tre locus and analysis of trehalose cryoprotection in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2006, 72, 1218–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Rabah, H.; Jardin, J.; Briard-Bion, V.; Parayre, S.; Maillard, M.B.; Loir, Y.l.; Chen, X.D.; Schuck, P.; Jeantet, R.; et al. Hyperconcentrated sweet whey, a new culture medium that enhances Propionibacterium freudenreichii stress tolerance. Appl. Environ. Microbiol. 2016, 82, 4641–4651. [Google Scholar] [CrossRef] [Green Version]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef]
- Hounsa, C.G.; Brandt, E.V.; Thevelein, J.; Hohmann, S.; Prior, B.A. Role of trehalose in survival of Saccharomyces cerevisiae under osmotic stress. Microbiology 1998, 144, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Mendes, L.; Bernardes, E.; de Araujo, P.S.; Panek, A.D.; Paschoalin, V.M.F. Preservation of frozen yeast cells by trehalose. Biotechnol. Bioeng. 1999, 65, 572–578. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Hugenholtz, J.; Zoon, P. An overview of the functionality of exopolysaccharides produced by lactic acid bacteria. Int. Dairy J. 2002, 12, 163–171. [Google Scholar] [CrossRef]
- Looijesteijn, P.J.; Trapet, L.; de Vries, E.; Abee, T.; Hugenholtz, J. Physiological function of exopolysaccharides produced by Lactococcus lactis. Int. J. Food Microbiol. 2001, 64, 71–80. [Google Scholar] [CrossRef]
- Stack, H.M.; Kearney, N.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Association of Beta-Glucan Endogenous Production with Increased Stress Tolerance of Intestinal Lactobacilli. Appl. Environ. Microbiol. 2010, 76, 500–507. [Google Scholar] [CrossRef]
- Billi, D.; Potts, M. Life and death of dried prokaryotes. Res. Microbiol. 2002, 153, 7–12. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| L. fermentum FXJCJ6-1 | L. reuteri CCFM1040 | L. brevis 173-1-2 | ||||
|---|---|---|---|---|---|---|
| Number of Active Bacteria (×1010)/ (CFU/mL) | Freeze-Drying Survival Rate/(%) | Number of Active Bacteria (×1010)/(CFU/mL) | Freeze-Drying Survival Rate/(%) | Number of Active Bacteria (×1010)/ (CFU/mL) | Freeze-Drying Survival Rate/(%) | |
| Glucose | 5.7 ± 0.29 | 32.39 ± 1.67 a | 2.31 ± 0.06 | 59.1 ± 1.57 a | 15.68 ± 0.59 | 60.52 ± 2.26 |
| Trehalose | - | - | - | - | - | - |
| Lactose | 10.88 ± 0.95 | 57.31 ± 1.74 c | 3.47 ± 0.14 | 66.78 ± 2.61 b | - | - |
| Sucrose | 7.05 ± 0.29 | 38.21 ± 1.56 b | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, S.; Hu, M.; Sun, Y.; Mao, B.; Zhang, Q.; Zhao, J.; Tang, X.; Zhang, H. Effect of Trehalose and Lactose Treatments on the Freeze-Drying Resistance of Lactic Acid Bacteria in High-Density Culture. Microorganisms 2023, 11, 48. https://doi.org/10.3390/microorganisms11010048
Cui S, Hu M, Sun Y, Mao B, Zhang Q, Zhao J, Tang X, Zhang H. Effect of Trehalose and Lactose Treatments on the Freeze-Drying Resistance of Lactic Acid Bacteria in High-Density Culture. Microorganisms. 2023; 11(1):48. https://doi.org/10.3390/microorganisms11010048
Chicago/Turabian StyleCui, Shumao, Mengyu Hu, Yuanyuan Sun, Bingyong Mao, Qiuxiang Zhang, Jianxin Zhao, Xin Tang, and Hao Zhang. 2023. "Effect of Trehalose and Lactose Treatments on the Freeze-Drying Resistance of Lactic Acid Bacteria in High-Density Culture" Microorganisms 11, no. 1: 48. https://doi.org/10.3390/microorganisms11010048