
Yeast Peptides Improve the Intestinal Barrier Function and Alleviate Weaning Stress by Changing the Intestinal Microflora Structure of Weaned Lambs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Regions, Animals, and Experimental Design
2.2. Feed Intake and Growth Performance Measurement
2.3. Blood Sample Collection and Measurement
2.4. Jejunum Sampling, Processing, and Histomorphology Analysis
2.5. DNA Extraction, PCR Amplification, and 16S rRNA Sequencing
2.6. Sequence Analysis
2.7. Statistical Analysis
3. Results
3.1. Feed Intake and Growth Performance
3.2. Serum Antibodies Concentrations
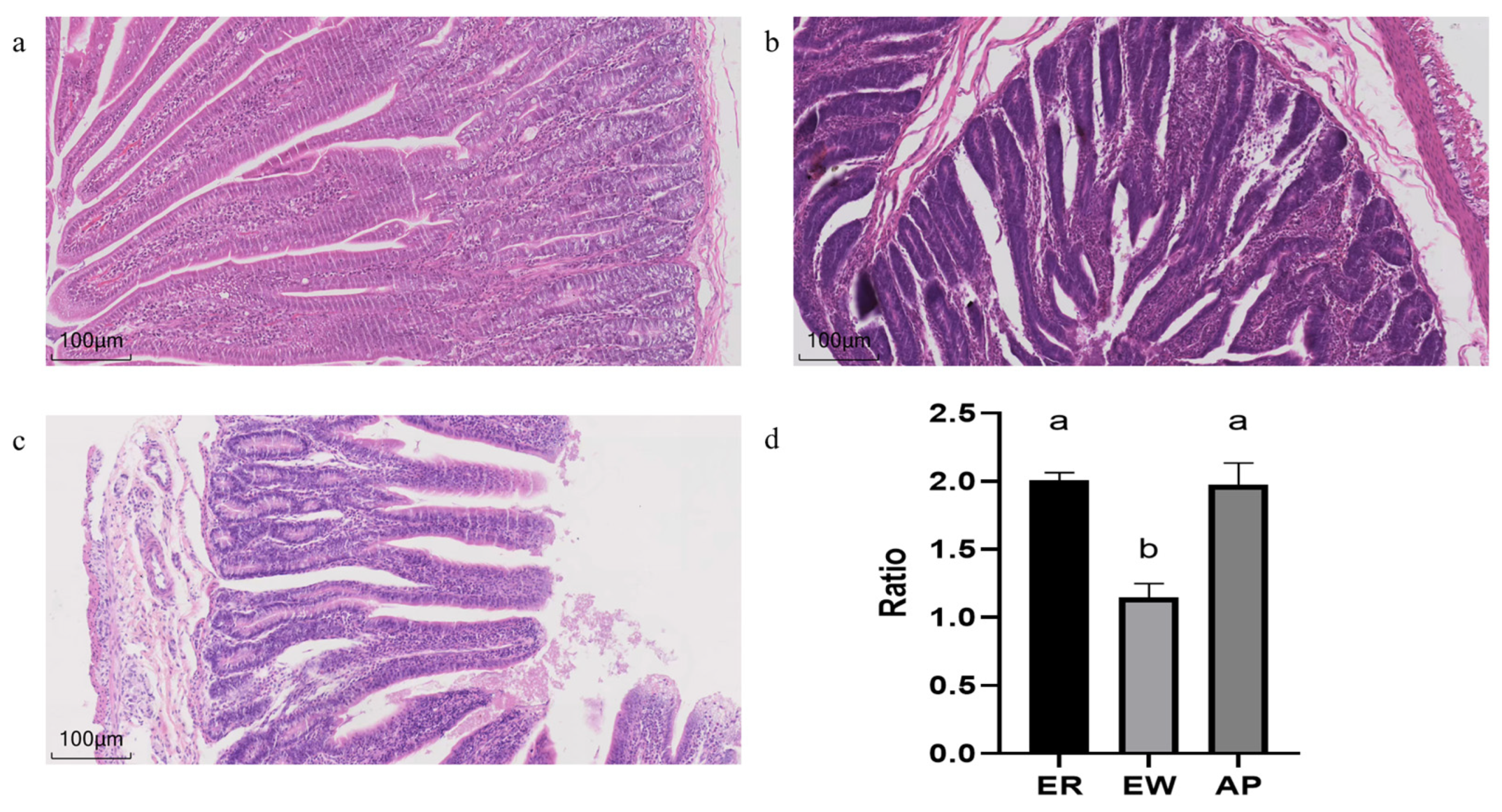
3.3. Small Intestinal Morphology
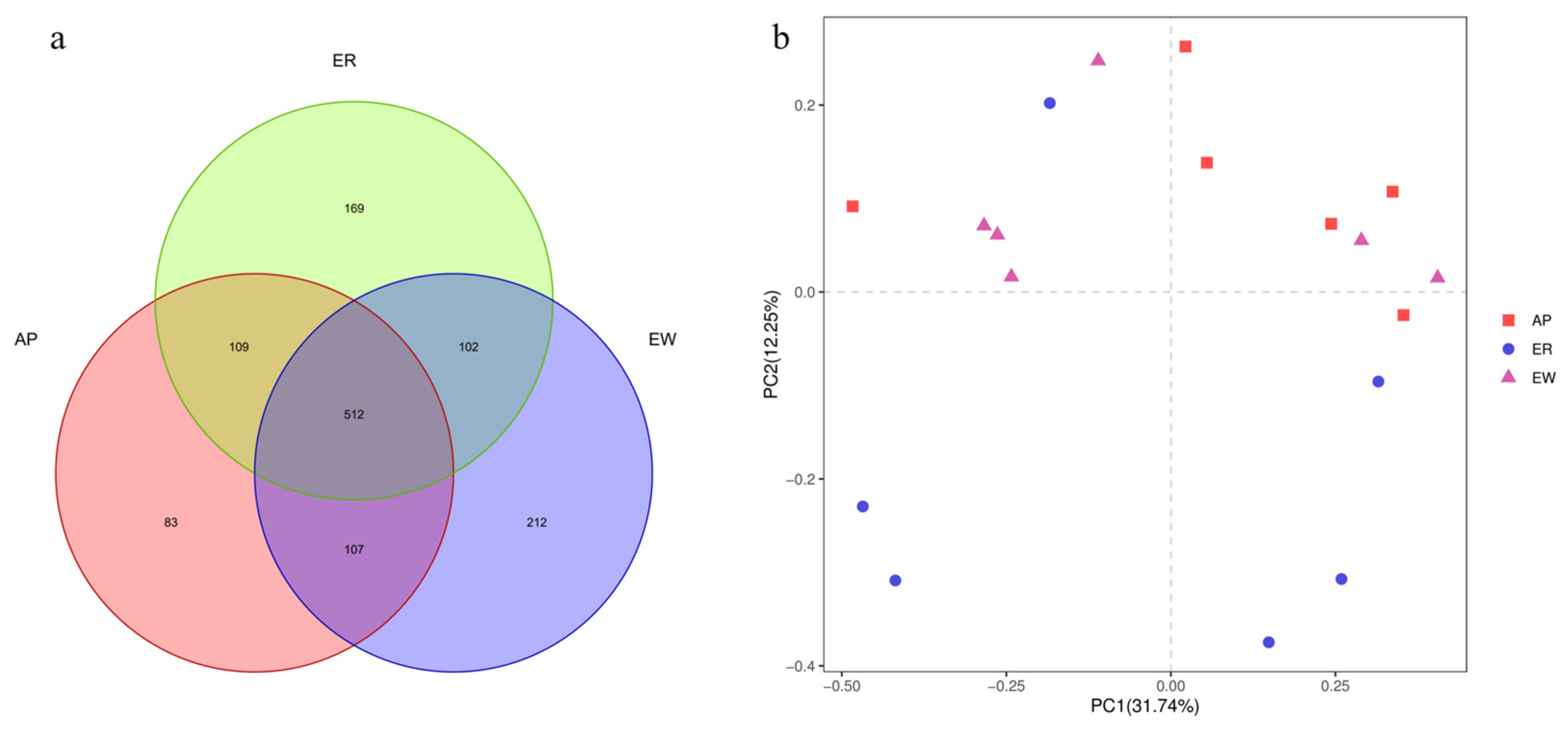
3.4. Taxonomic Composition of Gastrointestinal Bacteria
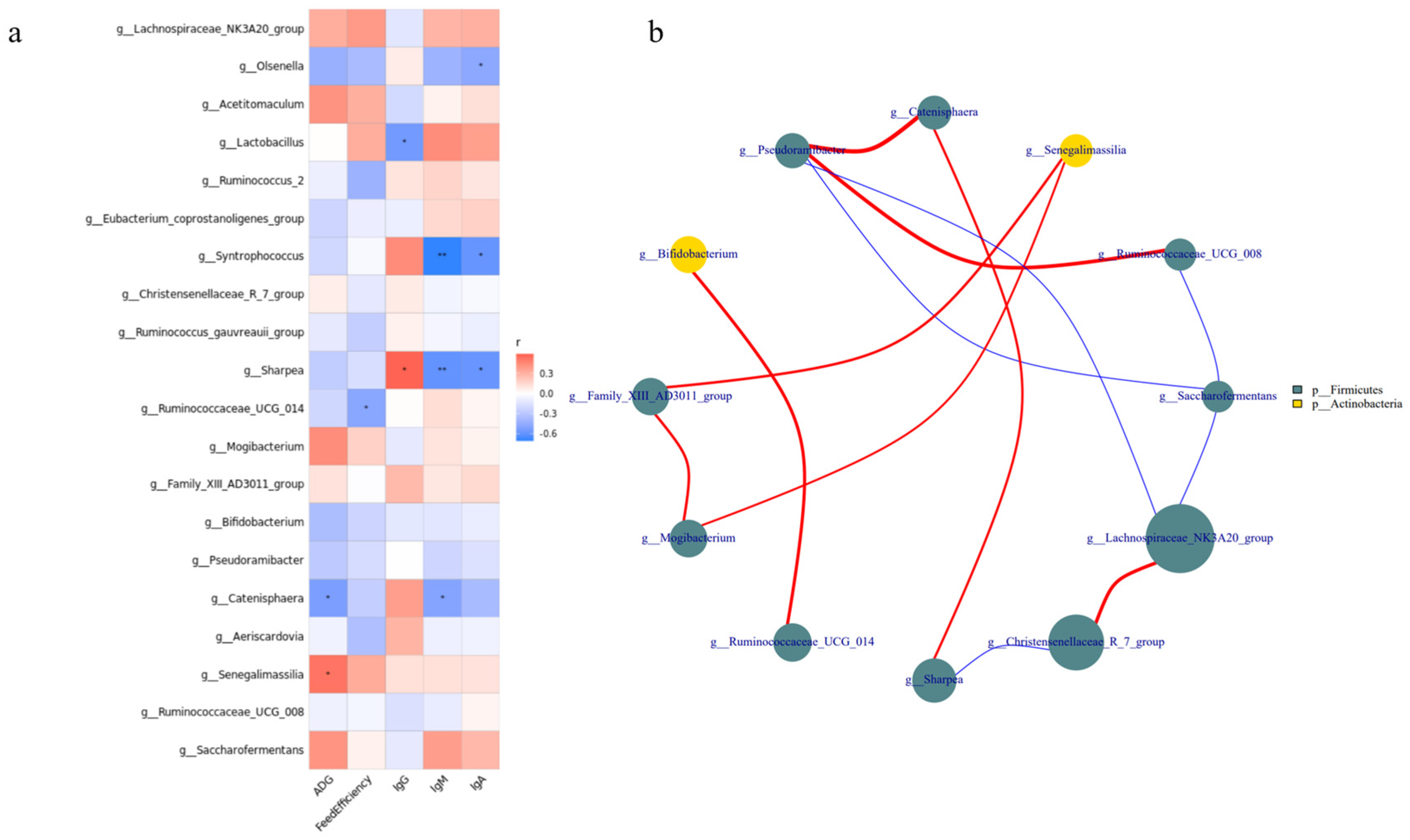
3.5. Growth Performance and Serum Immune Indicators Correlate with Bacterial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Han, L.; Tao, H.; Kang, L.; Wang, S.; Diao, Q.; Han, D.; Cui, K. Transcriptome and iTRAQ-Based Proteome Reveal the Molecular Mechanism of Intestinal Injury Induced by Weaning Ewe’s Milk in Lambs. Front. Vet. Sci. 2022, 9, 809188. [Google Scholar] [CrossRef]
- Belanche, A.; Cooke, J.; Jones, E.; Worgan, H.J.; Newbold, C.J. Short- and long-term effects of conventional and artificial rearing strategies on the health and performance of growing lambs. Animal 2019, 13, 740–749. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Novais, A.K.; Deschene, K.; Martel-Kennes, Y.; Roy, C.; Laforest, J.P.; Lessard, M.; Matte, J.J.; Lapointe, J. Weaning differentially affects mitochondrial function, oxidative stress, inflammation and apoptosis in normal and low birth weight piglets. PLoS ONE 2021, 16, e0247188. [Google Scholar] [CrossRef] [PubMed]
- Moeser, A.J.; Pohl, C.S.; Rajput, M. Weaning stress and gastrointestinal barrier development: Implications for lifelong gut health in pigs. Anim. Nutr. 2017, 3, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Li, D.; Yin, Y.; Wang, X.; Li, P.; Dangott, L.J.; Hu, W.; Wu, G. Intrauterine Growth Restriction Affects the Proteomes of the Small Intestine, Liver, and Skeletal Muscle in Newborn Pigs. J. Nutr. 2008, 138, 60–66. [Google Scholar] [CrossRef]
- Cui, K.; Wang, B.; Zhang, N.; Tu, Y.; Ma, T.; Diao, Q. iTRAQ-based quantitative proteomic analysis of alterations in the intestine of Hu sheep under weaning stress. PLoS ONE 2018, 13, e0200680. [Google Scholar] [CrossRef]
- Pascual-Alonso, M.; Miranda-de la Lama, G.C.; Aguayo-Ulloa, L.; Ezquerro, L.; Villarroel, M.; Marin, R.H.; Maria, G.A. Effect of postweaning handling strategies on welfare and productive traits in lambs. J. Appl. Anim. Welf. Sci. 2015, 18, 42–56. [Google Scholar] [CrossRef]
- Lessard, M.; Blais, M.; Beaudoin, F.; Deschene, K.; Verso, L.L.; Bissonnette, N.; Lauzon, K.; Guay, F. Piglet weight gain during the first two weeks of lactation influences the immune system development. Vet. Immunol. Immunopathol. 2018, 206, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2015, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Maranduba, C.M.; De Castro, S.B.; Souza, G.T.; Rossato, C.; da Guia, F.C.; Valente, M.A.; Rettore, J.V.; Maranduba, C.P.; Souza, C.M.; Carmo, A.M.; et al. Intestinal Microbiota as Modulators of the Immune System and Neuroimmune System: Impact on the Host Health and Homeostasis. J. Immunol. Res. 2015, 2015, 931574. [Google Scholar] [CrossRef]
- Li, C.; Wang, W.; Liu, T.; Zhang, Q.; Wang, G.; Li, F.; Li, F.; Yue, X.; Li, T. Effect of Early Weaning on the Intestinal Microbiota and Expression of Genes Related to Barrier Function in Lambs. Front. Microbiol. 2018, 9, 1431. [Google Scholar] [CrossRef]
- Mirzaei, M.; Shavandi, A.; Mirdamadi, S.; Soleymanzadeh, N.; Motahari, P.; Mirdamadi, N.; Moser, M.; Subra, G.; Alimoradi, H.; Goriely, S. Bioactive peptides from yeast: A comparative review on production methods, bioactivity, structure-function relationship, and stability. Trends Food Sci. Technol. 2021, 118, 297–315. [Google Scholar] [CrossRef]
- Mirzaei, M.; Mirdamadi, S.; Safavi, M.; Zare, D.; Hadizadeh, M.; Asadi, M.M. Synthesis, in vitro and cellular antioxidant activity evaluation of novel peptides derived from Saccharomyces cerevisiae protein hydrolysate: Structure-function relationship: Antioxidant activity and synthetic peptides. Amino Acids 2019, 51, 1167–1175. [Google Scholar] [CrossRef]
- Jung, E.Y.; Lee, H.-S.; Choi, J.W.; Ra, K.S.; Kim, M.-R.; Suh, H.J. Glucose Tolerance and Antioxidant Activity of Spent Brewer’s Yeast Hydrolysate with a High Content of Cyclo-His-Pro (CHP). J. Food Sci. 2011, 76, C272–C278. [Google Scholar] [CrossRef] [PubMed]
- Poppy, G.D.; Rabiee, A.R.; Lean, I.J.; Sanchez, W.K.; Dorton, K.L.; Morley, P.S. A meta-analysis of the effects of feeding yeast culture produced by anaerobic fermentation of Saccharomyces cerevisiae on milk production of lactating dairy cows. J. Dairy Sci. 2012, 95, 6027–6041. [Google Scholar] [CrossRef] [PubMed]
- Pas, J.; Pas, T.; Pas, C.; Dorton, K.L. Meta-analysis examining the effects of Saccharomyces cerevisiae fermentation products on feedlot performance and carcass traits 1, 2, 3. Prof. Anim. Sci. 2016, 32, 172–182. [Google Scholar]
- Mirzaei, M.; Mirdamadi, S.; Safavi, M. Antioxidant activity and protective effects of Saccharomyces cerevisiae peptide fractions against H2O2-induced oxidative stress in Caco-2 cells. J. Food Meas. Charact. 2019, 13, 2654–2662. [Google Scholar] [CrossRef]
- Du, D.; Feng, L.; Chen, P.; Jiang, W.; Zhang, Y.; Liu, W.; Zhai, R.; Hu, Z. Effects of Saccharomyces cerevisiae Cultures on Performance and Immune Performance of Dairy Cows During Heat Stress. Front. Vet. Sci. 2022, 9, 851184. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, J.; Hou, Y.; Hu, S.Q. Production of yeast hydrolysates by Bacillus subtilis derived enzymes and antihypertensive activity in spontaneously hypertensive rats. Food Biotechnol. 2020, 34, 262–281. [Google Scholar] [CrossRef]
- Amorim, M.M.; Pereira, J.O.; Monteiro, K.M.; Ruiz, A.L.; Carvalho, J.O.E.; Pinheiro, H.; Pintado, M. Antiulcer and antiproliferative properties of spent brewer’s yeast peptide extracts for incorporation into foods. Food Funct. 2016, 7, 2331–2337. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M.; Bourtzis, K. Development of a Prokaryotic Universal Primer for Simultaneous Analysis of Bacteria and Archaea Using Next-Generation Sequencing. PLoS ONE 2014, 45, e105592. [Google Scholar] [CrossRef]
- Han, L.; Xue, W.; Cao, H.; Chen, X.; Qi, F.; Ma, T.; Tu, Y.; Diao, Q.; Zhang, C.; Cui, K. Comparison of Rumen Fermentation Parameters and Microbiota of Yaks From Different Altitude Regions in Tibet, China. Front. Microbiol. 2021, 12, 807512. [Google Scholar] [CrossRef] [PubMed]
- Mago, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194. [Google Scholar]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Wang, Q. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Glckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Miller, G.E.; Engen, P.A.; Gillevet, P.M.; Shaikh, M.; Sikaroodi, M.; Forsyth, C.B.; Mutlu, E.; Keshavarzian, A. Lower Neighborhood Socioeconomic Status Associated with Reduced Diversity of the Colonic Microbiota in Healthy Adults. PLoS ONE 2016, 11, e0148952. [Google Scholar] [CrossRef]
- de la Fuente, J.; Tejon, D.; Rey, A.; Thos, J.; Lopez-Bote, C.J. Effect of rearing system on growth, body composition and development of digestive system in young lambs. J. Anim. Physiol. Anim. Nutr. 1997, 78, 75–83. [Google Scholar] [CrossRef]
- Wang, S.; Ma, T.; Zhao, G.; Zhang, N.; Tu, Y.; Li, F.; Cui, K.; Bi, Y.; Ding, H.; Diao, Q. Effect of Age and Weaning on Growth Performance, Rumen Fermentation, and Serum Parameters in Lambs Fed Starter with Limited Ewe–Lamb Interaction. Animals 2019, 9, 825. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, Q.; Wen, L.; Wu, C.; Yao, Z.; Yan, Z.; Li, R.; Chen, L.; Chen, F.; Xie, Z.; et al. The Effect of the Antimicrobial Peptide Plectasin on the Growth Performance, Intestinal Health, and Immune Function of Yellow-Feathered Chickens. Front. Vet. Sci. 2021, 8, 688611. [Google Scholar] [CrossRef] [PubMed]
- Palmeira, P.; Quinello, C.; Silveira-Lessa, A.L.; Zago, C.A.; Carneiro-Sampaio, M. IgG Placental Transfer in Healthy and Pathological Pregnancies. Clin. Dev. Immunol. 2012, 2012, 985646. [Google Scholar] [CrossRef] [PubMed]
- Furman-Fratczak, K.; Rzasa, A.; Stefaniak, T. The influence of colostral immunoglobulin concentration in heifer calves’ serum on their health and growth. J. Dairy. Sci. 2011, 94, 5536–5543. [Google Scholar] [CrossRef]
- Gesualdo, L.; Di Leo, V.; Coppo, R. The mucosal immune system and IgA nephropathy. Semin. Immunopathol. 2021, 43, 657–668. [Google Scholar] [CrossRef]
- Chen, K.; Magri, G.; Grasset, E.K.; Cerutti, A. Rethinking mucosal antibody responses: IgM, IgG and IgD join IgA. Nat. Rev. Immunol. 2020, 20, 427–441. [Google Scholar] [CrossRef]
- Abdelsattar, M.M.; Vargas-Bello-Pérez, E.; Zhang, N. Age-related changes in blood biochemical composition of Hu sheep. Ital. J. Anim. Sci. 2022, 21, 1297–1306. [Google Scholar] [CrossRef]
- Tothova, C.; Link, R.; Kyzekova, P.; Nagy, O. Serum protein electrophoretic pattern in piglets during the early postnatal period. Sci. Rep. 2021, 11, 17539. [Google Scholar] [CrossRef]
- Zhu, H.L.; Zhao, X.W.; Chen, S.; Tan, W.; Han, R.W.; Qi, Y.X.; Huang, D.W.; Yang, Y.X. Evaluation of colostrum bioactive protein transfer and blood metabolic traits in neonatal lambs in the first 24 hours of life. J. Dairy. Sci. 2021, 104, 1164–1174. [Google Scholar] [CrossRef]
- Yi, H.; Yang, G.; Xiong, Y.; Wen, X.; Wang, Z.; Yang, X.; Gao, K.; Wang, L.; Jiang, Z. Effects of Lactobacillus reuteri LR1 on the growth performance, intestinal morphology and intestinal barrier function in weaned pigs. J. Anim. Sci. 2018, 96, 2342–2351. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, Y.; Wen, X.; Wang, L.; Jiang, Z.; Zheng, C. Effects of low-molecular-weight chitosan on the growth performance, intestinal morphology, barrier function, cytokine expression and antioxidant system of weaned piglets. BMC Vet. Res. 2018, 14, 215. [Google Scholar] [CrossRef]
- Wang, M.; Huang, H.; Liu, S.; Zhuang, Y.; Yang, H.; Li, Y.; Chen, S.; Wang, L.; Yin, L.; Yao, Y. Tannic acid modulates intestinal barrier functions associated with intestinal morphology, antioxidative activity, and intestinal tight junction in a diquat-induced mouse model. RSC Adv. 2019, 9, 31988–31998. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lee, S.H.; Park, Y.K.; Kwon, I.K.; Chae, B.J. Effects of dietary supplementation of antimicrobial peptide-A3 on growth performance, nutrient digestibility, intestinal and fecal microflora and intestinal morphology in weanling pigs. Anim. Feed. Sci. Technol. 2012, 177, 98–107. [Google Scholar] [CrossRef]
- Lam, S.; Bai, X.; Shkoporov, A.N.; Park, H.; Wu, X.; Lan, P.; Zuo, T. Roles of the gut virome and mycobiome in faecal microbiota transplantation. Lancet. Gastroenterol. Hepatol. 2022, 7, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.A.; Baffy, N. Modulation of the gut microbiota: A focus on treatments for irritable bowel syndrome. Postgrad. Med. 2017, 129, 872–888. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Moal, L.L.; Servin, A.L. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: Mucins, antimicrobial peptides, and microbiota. Clin. Microbiol. Rev. 2006, 19, 315–337. [Google Scholar] [CrossRef]
- Gao, J.; Li, Y.; Wan, Y.; Hu, T.; Liu, L.; Yang, S.; Gong, Z.; Zeng, Q.; Wei, Y.; Yang, W.; et al. A Novel Postbiotic from Lactobacillus rhamnosus GG With a Beneficial Effect on Intestinal Barrier Function. Front. Microbiol. 2019, 10, 477. [Google Scholar] [CrossRef]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Lyons, A.; O’Mahony, D.; O’Brien, F.; MacSharry, J.; Sheil, B.; Ceddia, M.; Russell, W.M.; Forsythe, P.; Bienenstock, J.; Kiely, B.; et al. Bacterial strain-specific induction of Foxp3+ T regulatory cells is protective in murine allergy models. Clin. Exp. Allergy 2010, 40, 811–819. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Englyst, H.N. Starch utilization by the human large intestinal microflora. J. Appl. Bacteriol. 1986, 60, 195–201. [Google Scholar] [CrossRef]
- Scott, K.P.; Martin, J.C.; Duncan, S.H.; Flint, H.J. Prebiotic stimulation of human colonic butyrate-producing bacteria and bifidobacteria, in vitro. FEMS Microbiol. Ecol. 2014, 87, 30–40. [Google Scholar] [CrossRef]
- Willemsen, L.E.M.; Koetsier, M.A.; Deventer, S.J.H.V.; Tol, E.A.F.V. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E1 and E2 production by intestinal myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, S.; Toh, H.; Taylor, T.D.; Ohno, H.; Hattori, M. Acetate-producing bifidobacteria protect the host from enteropathogenic infection via carbohydrate transporters. Gut Microbes 2012, 3, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Arvans, D.L.; Vavricka, S.R.; Ren, H.; Musch, M.W.; Kang, L.; Rocha, F.G.; Lucioni, A.; Turner, J.R.; Alverdy, J.; Chang, E.B. Luminal bacterial flora determines physiological expression of intestinal epithelial cytoprotective heat shock proteins 25 and 72. Am. J. Physiol.-Gastrointest. Liver Physiol. 2005, 288, G696–G704. [Google Scholar] [CrossRef]
- Duca, F.A.; Sakar, Y.; Covasa, M. The modulatory role of high fat feeding on gastrointestinal signals in obesity. J. Nutr. Biochem. 2013, 24, 1663–1677. [Google Scholar] [CrossRef]
- Cani, P.D.; Possemiers, S.; Van de Wiele, T.; Guiot, Y.; Everard, A.; Rottier, O.; Geurts, L.; Naslain, D.; Neyrinck, A.; Lambert, D.M.; et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut 2009, 58, 1091–1103. [Google Scholar] [CrossRef]
- O’Mahony, L.; Feeney, M.; O’Halloran, S.; Murphy, L.; Kiely, B.; Fitzgibbon, J.; Lee, G.; O’Sullivan, G.; Shanahan, F.; Collins, J.K. Probiotic impact on microbial flora, inflammation and tumour development in IL-10 knockout mice. Aliment. Pharmacol. Ther. 2001, 15, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Cano, P.G.; Santacruz, A.; Trejo, F.M.; Sanz, Y. Bifidobacterium CECT 7765 improves metabolic and immunological alterations associated with obesity in high-fat diet-fed mice. Obesity 2013, 21, 2310–2321. [Google Scholar] [CrossRef] [PubMed]
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tome, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Tang, S.; Xie, J.; Fang, W.; Wen, X.; Yin, C.; Meng, Q.; Zhong, R.; Chen, L.; Zhang, H. Chronic heat stress induces the disorder of gut transport and immune function associated with endoplasmic reticulum stress in growing pigs. Anim. Nutr. 2022, 11, 228–241. [Google Scholar] [CrossRef]
- Han, K.-I.; Kim, J.-S.; Lee, K.C.; Eom, M.K.; Suh, M.K.; Kim, H.S.; Park, S.-H.; Lee, J.H.; Kang, S.W.; Park, J.-E.; et al. Senegalimassilia faecalis sp. nov., an anaerobic actinobacterium isolated from human faeces, and emended description of the genus Senegalimassilia. Int. J. Syst. Evol. Microbiol. 2020, 70, 1684–1690. [Google Scholar] [CrossRef]
- Jin, M.; Li, D.; Ji, R.; Liu, W.; Xu, X.; Li, Y. Changes in intestinal microflora in digestive tract diseases during pregnancy. Arch. Gynecol. Obstet. 2020, 301, 243–249. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Wang, H.; Ma, Y.; Zhao, X.; Zhang, X.; Yang, H.; Qian, J.; Li, J. Saccharomyces boulardii alleviates ulcerative colitis carcinogenesis in mice by reducing TNF-α and IL-6 levels and functions and by rebalancing intestinal microbiota. BMC Microbiol. 2019, 19, 246. [Google Scholar] [CrossRef] [PubMed]
- Xing, S.; Chen, S.; Zhao, Y.; Luo, Y.; Yu, B.; He, J.; Huang, Z.; Zheng, P.; Mao, X.; Luo, J.; et al. Effects of High Ambient Temperature on Small Intestinal Morphology and Colonic Microbiota in Weaned Piglets. Animals 2022, 12, 1743. [Google Scholar] [CrossRef] [PubMed]
- Geng, S.; Yang, L.; Cheng, F.; Zhang, Z.; Li, J.; Liu, W.; Li, Y.; Chen, Y.; Bao, Y.; Chen, L.; et al. Gut Microbiota Are Associated With Psychological Stress-Induced Defections in Intestinal and Blood-Brain Barriers. Front. Microbiol. 2019, 10, 3067. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Content |
|---|---|
| Ingredients | (% of feeding basis) |
| Corn | 40.0 |
| Soybean meal | 20.0 |
| Whey powder | 20.0 |
| Alfalfa hay | 16.0 |
| Premix (1) | 4.0 |
| Total | 100.0 |
| Nutrient levels (2) | (% dry matter) |
| NEL/(MJ/kg) | 11.59 |
| CP | 17.15 |
| EE | 3.87 |
| Ash | 9.10 |
| NDF | 20.61 |
| ADF | 11.29 |
| Ca | 1.45 |
| P | 0.57 |
| Items | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| ER | EW | AP | |||
| BW, kg | |||||
| 1 d | 3.32 b | 3.59 ab | 3.81 a | 0.08 | 0.045 |
| 21 d | 6.76 | 6.85 | 7.43 | 0.18 | 0.275 |
| 28 d | 8.13 | 9.03 | 8.44 | 0.22 | 0.262 |
| 35 d | 9.53 | 9.49 | 9.24 | 0.24 | 0.870 |
| ADG, g/d | |||||
| Pre-weaning | 171.83 | 194.25 | 165.48 | 7.24 | 0.673 |
| Post-weaning | 199.21 a | 65.24 b | 113.89 b | 13.73 | <0.001 |
| DMI (1), g/d | |||||
| Pre-weaning | 34.44 | 23.41 | 46.43 | 6.52 | 0.407 |
| Post-weaning | 130.08 | 128.89 | 164.92 | 15.20 | 0.612 |
| Feed Efficiency (2),% | |||||
| Pre-weaning | 6.65 b | 11.73 a | 6.24 b | 0.72 | 0.002 |
| Post-weaning | 1.65 a | 0.43 b | 0.73 b | 0.12 | <0.001 |
| Items | Treatment | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| ER | EW | AP | T | D | T × D | ||
| IgG, g/L | |||||||
| 21 d | 16.28 b | 17.56 a | 17.43 a | 0.18 | 0.001 | ||
| 28 d | 16.72 | 17.00 | 16.70 | 0.12 | 0.529 | ||
| 31 d | 18.25 a | 16.85 b | 16.78 b | 0.21 | 0.001 | ||
| 35 d | 15.99 | 16.39 | 16.68 | 0.20 | 0.394 | ||
| Overall | 16.81 | 16.95 | 16.89 | 0.10 | 0.726 | <0.001 | <0.001 |
| IgA, g/L | |||||||
| 21 d | 0.69 a | 0.62 c | 0.65 b | 0.01 | <0.001 | ||
| 28 d | 0.70 a | 0.63 b | 0.64 b | 0.01 | 0.001 | ||
| 31 d | 0.55 b | 0.68 a | 0.66 a | 0.01 | <0.001 | ||
| 35 d | 0.73 | 0.66 | 0.70 | 0.01 | 0.172 | ||
| Overall | 0.67 | 0.65 | 0.66 | 0.01 | 0.145 | <0.001 | <0.001 |
| IgM, g/L | |||||||
| 21 d | 1.02 a | 0.92 b | 0.94 b | 0.01 | 0.001 | ||
| 28 d | 1.00 a | 0.93 b | 0.96 ab | 0.01 | 0.041 | ||
| 31 d | 0.79 b | 1.01 a | 0.97 a | 0.02 | <0.001 | ||
| 35 d | 1.07 | 0.97 | 1.02 | 0.02 | 0.209 | ||
| Overall | 0.97 | 0.96 | 0.97 | 0.01 | 0.598 | <0.001 | <0.001 |
| Items | Groups | SEM | p | ||
|---|---|---|---|---|---|
| ER | EW | AP | |||
| Chao1 | 470.31 | 408.99 | 441.47 | 27.26 | 0.682 |
| Observed_species | 333.82 | 319.98 | 340.48 | 23.02 | 0.941 |
| PD_whole_tree | 30.59 | 30.79 | 31.53 | 1.96 | 0.981 |
| Shannon | 4.69 | 4.54 | 4.74 | 0.15 | 0.854 |
| Simpson | 0.89 | 0.89 | 0.91 | 0.13 | 0.850 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Han, L.; Liu, J.; Kang, L.; Zhao, L.; Cui, K. Yeast Peptides Improve the Intestinal Barrier Function and Alleviate Weaning Stress by Changing the Intestinal Microflora Structure of Weaned Lambs. Microorganisms 2023, 11, 2472. https://doi.org/10.3390/microorganisms11102472
Li Y, Han L, Liu J, Kang L, Zhao L, Cui K. Yeast Peptides Improve the Intestinal Barrier Function and Alleviate Weaning Stress by Changing the Intestinal Microflora Structure of Weaned Lambs. Microorganisms. 2023; 11(10):2472. https://doi.org/10.3390/microorganisms11102472
Chicago/Turabian StyleLi, Yanjun, Lulu Han, Jie Liu, Lingyun Kang, Ling Zhao, and Kai Cui. 2023. "Yeast Peptides Improve the Intestinal Barrier Function and Alleviate Weaning Stress by Changing the Intestinal Microflora Structure of Weaned Lambs" Microorganisms 11, no. 10: 2472. https://doi.org/10.3390/microorganisms11102472
APA StyleLi, Y., Han, L., Liu, J., Kang, L., Zhao, L., & Cui, K. (2023). Yeast Peptides Improve the Intestinal Barrier Function and Alleviate Weaning Stress by Changing the Intestinal Microflora Structure of Weaned Lambs. Microorganisms, 11(10), 2472. https://doi.org/10.3390/microorganisms11102472