Abstract
Entomopathogenic nematodes have been used in biological control for some time and are an alternative for the control of insect pests, but during their implementation, situations have arisen that can be improved. These vary with each species and include their production and storage. Oscheius myriophila, an entomopathogenic nematode (EPN), was monitored for its performance when produced in vivo, as well as its development using Galleria mellonella larvae, using the MC5-2014 strain isolated from soil samples in the municipality of Tepalcingo, Morelos, México. For a study with native strains of EPNs, a wide range of tests must be conducted because the required conditions can be very specific. In vivo production was quantified at initial infective juvenile (IJ) inocula of 50, 100 and 500, and we obtained the same production for the three inocula. The life cycle of the EPNs lasted 12 days, and two generations were observed in which adults were found at days 5 and 9. Both evaluations were performed at a temperature of 27 °C in G. mellonella larvae. In addition, the temperatures of 8, 12, 20 and 24 °C were evaluated for their storage, and we observed that the EPNs can be kept for at least 6 months, maintaining a survival rate of 58.67% and a good infective capacity at a temperature of 12 °C, remaining above 60%.
1. Introduction
Entomopathogenic nematodes (EPNs) are a promising alternative for the control of insect pests due to their wide range of hosts and active search method [1]. Although much information on EPNs has been generated since the 1980s, most of it focuses on the families Steinernematidae [2] and Heterorhabditidiae [3], which are associated with their symbiotic bacteria Xenorhabdus and Photorhabdus, respectively [4,5]. In addition to these families, species of EPNs belonging to the Rhabditidae family have been reported [6,7,8], and in turn, other species, for example, Serratia, have been reported to be symbiotic bacterial species [5,9].
Derived from EPN search and identification work, the Biological Control Laboratory of the Biotechnology Research Center of the Autonomous University of the State of Morelos (CEIB/UAEM) has EPNs that have not yet been reported in Mexico, one of which is O. myriophila [10], a species belonging to the Rhabditidae family.
For a study with native strains of EPNs, a wide range of tests must be conducted because the required conditions can be very specific; therefore, it is important to determine their development and the temperatures at which they are produced in greater volume and to preserve them for a longer period of time while maintaining their infectivity [11].
Carrying out propagations in vivo and determining the optimal conditions allowed us to perform an in-depth study. In vivo production is a simple process in which live host insects are used. This approach has various advantages, such as low start-up costs and little use of technology. This is an important production method because it allows us to obtain high-quality IJs. G. mellonella larvae are used as a substitute host [12], and are commercially produced in large quantities in several countries and susceptible to most nematode species [13].
In turn, G. mellonella is also used to observe the development of the EPN, learn more about it, and obtain information including its life cycle duration and the number of generations it could produce within the host [13,14,15]. In terms of the abovementioned families, the storage conditions for steinernematids involve temperatures that range from 8 to 15 °C, and they survive for 6 to 9 months; on the other hand, heterorhabdithids, when kept under the same conditions, survive for 3 to 4 months [16]. Therefore, knowing the requirements of each species is important in this field of study because they can vary considerably; currently, it is believed that it is best to use native species of EPNs in geographical areas where biocontrol is applied [17,18,19].
2. Materials and Methods
2.1. G. mellonella Production
G. mellonella was kept in the incubator at 30 ± 2 °C, and fed with a mixture that included rice flour, brewer’s yeast, wheat bran, glycerin and honey.
2.2. O. myriophila Strain MC5-2014 Propagation
At the bottom of 90 mm diameter × 15 mm depth Petri dishes, on a disk of medium-pore filter paper, 5 G. mellonella larvae were placed and 200 μL of suspension with EPNs was added for each larva; at the end of 5 days, the larvae were placed in white traps maintained at 27 °C with periods of daylight [20].
Once the EPNs reproduced and developed into new generations, they were deposited in 50 mL Corning culture flasks with 20 mL of sterile distilled water and kept refrigerated at 12 ± 1 °C to obtain the initial concentrated suspension [21].
2.3. Quantification of IJs
The method proposed by Woodring and Kaya [21] was used. For this, the initial suspension contained in the Corning flasks was homogenized. Then, 100 μL of suspension was obtained and deposited in a Petri dish, followed by counting the IJs with a stereoscopic microscope (Motic instruments, DM143, Hong Kong, China). This process was repeated five times, and the average counts were calculated to determine the inoculum of the EPN suspensions.
Using the formula of Woodring and Kaya [21], we obtained the required dilutions to use in each of the bioassays:
where:
A = Volume of the concentrated suspension to be diluted (mL),
B = Number of nematodes in 1 mL diluted suspension (IJs/mL),
C = Final volume in milliliters of the new dilution (mL),
D = Desired nematode inoculum in the new dilution (IJs/mL) [21].
2.4. Bioassays
In Vivo Production
G. mellonella larvae were used, which were previously weighed; their weights were between 0.2 and 0.3 g.
The G. mellonella larvae were placed on medium-pore filter paper disks at the bottom of the Petri dishes (90 diameter × 15 mm depth). Then, we added different inoculum numbers of initial IJs, i.e., 50, 100 and 500 IJs.
Our experimental unit comprised 5 Petri dishes with G. mellonella larvae, adding the different inocula of 50, 100 and 500 IJs; thus, there were 3 treatments with 5 repetitions each. We evaluated the production obtained in each experimental unit, for which modified white traps were mounted once the larvae were completely dead, and a single trap was used for each repetition. Five larvae were in each group. All the larvae died from day 6 and were kept at a temperature of 27 °C. Throughout the experiment, the counts were made from day 8 of the larvae infection.
Monitoring was performed every 48 h by collecting the water from the white traps and counting the IJs produced during that time using a stereoscopic microscope (Motic instruments, DM143, China) [21].
2.5. Development of O. myriophila
To observe the development of the EPNs and confirm the presence of 2 or 3 generations, 14 Petri dishes with dimensions of 60 diameter × 15 mm depth were used with filter paper at the bottom, which was previously moistened with 400 µL of sterile distilled water. To each Petri dish, 5 larvae of G. mellonella and 500 IJs for each larva were added, with a total of 2500 IJs per Petri dish. The temperature was maintained at 27 °C, and monitoring started 8 h after placing the IJs. The progress was checked at 12 h and then every 24 h for the duration of the cycle, taking 5 larvae every 24 h during the days evaluated, and the different stages of O. myriophila were observed.
2.6. Fixation of EPNs
Five G. mellonella larvae exposed to the EPNs were taken; they were passed through 5% chlorine and sterile distilled water and later dissected and analyzed. The EPNs were observed with a stereoscopic microscope (Motic instruments, DM143, China). Later, they were fixed with 4% saline formalin at boiling point and individually mounted on a slide; the EPNs from each stage were taken [22].
2.7. Cleared of EPNs
To observe the EPNs in detail, they were evaluated after they were fixed using heat. A nematode was placed on a slide and different concentrations of glycerin and water were added in the following order: 1/20, 1/15, 1/10, 1/5 and 1/1. The different concentrations were added to the slides, which were placed on a heating plate.
When these processes were complete, the EPNs were observed under an optical microscope (Leica DM500, St. Gallen, Switzerland), and each stage was analyzed by measuring their length and width using Leica Application Suite, version 3.4.0 [23].
2.8. Storage of IJs of O. myriophila
Corning culture flasks (50 mL) were used, allowing the EPNs to receive oxygen. For each treatment (at temperatures of 8, 12, 24 and 27 °C), there were 15 flasks with a capacity of 50 mL, with 3000 IJs contained in 40 m water. We performed 3 repetitions since they were independent experiments. Subsequently, quantification was performed every 15 days to determine the survival percentage at different temperatures; thus, the mortality percentage was determined [21,24].
2.9. IJ Pathogenicity
To perform the pathogenicity bioassays, 4 flasks stored at various temperatures (8, 12, 20 and 27 °C) were taken at random, and they were quantified to obtain the average number of live and dead IJs. Once these data were obtained, a sixth or seventh instar G. mellonella larva was placed individually in a 60 diameter × 15 mm depth Petri dish with filter paper at the bottom, and a suspension of 250 μL of sterile distilled water with 10 IJs was added to each Petri dish with its respective G. mellonella larva.
Each experimental unit was made up of 10 G. mellonella larvae, with four repetitions for each temperature, and mortality was monitored at 48, 72, 96 and 120 h at 27 °C. The larvae were considered dead when a flaccid appearance and change in color appeared. Likewise, the presence of EPNs was verified with a stereoscopic microscope (Motic instruments, DM143, China).
2.10. Statistical Analysis
The obtained results in in vivo production were transformed with the square root method and analyzed with the statistical package SAS 9.1; ANOVA was performed with its respective Tukey test. Standard deviations of the means were calculated in Microsoft Excel Office 365.
The obtained data in bioassays of storage were transformed using arcsine and analyzed with an analysis of variance (ANOVA) and a Tukey multiple comparison of means at 0.05 probability using the R Commander statistical package 2.7-1 version. Standard deviations of the means were calculated in Microsoft Excel Office 365.
3. Results
3.1. In Vivo Production
When evaluating the production at three different initial inocula, different quantities were obtained and no significant differences were observed (Table 1).

Table 1.
Number of IJs belonging to O. myriophila produced in G. mellonella larvae at different days with three different initial inocula of IJs at a temperature of 27 °C; the mean ± SD is shown. p > 0.05 (the same letters indicate no significant differences).
3.2. Development of O. myriophila
The development of the MC5-2014 strain belonging to the O. myriophila species was monitored at a temperature of 27 °C and an inoculum of 500 IJs per G. mellonella larva. From day 4, 100% mortality of the infected larvae was observed, and the following was discovered (Figure 1):
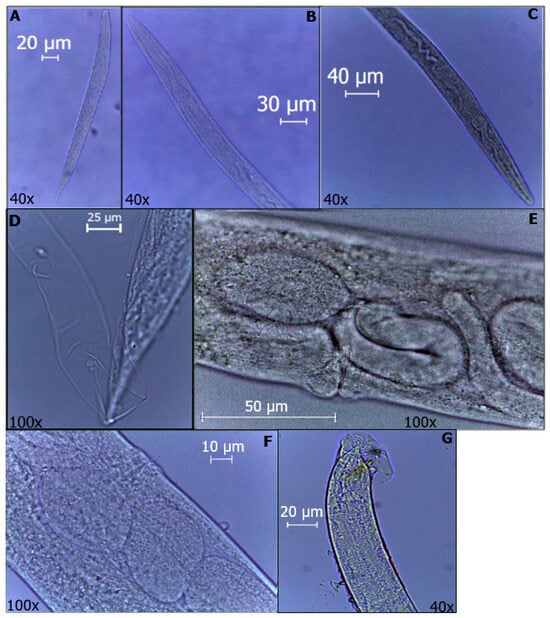
Figure 1.
Developmental stages of O. myriophila. (A) J1. (B) J2. (C) J3. (D) J4. (E) Endotokia matricida of female. (F) Female with eggs inside. (G) Male rays and spicules are visible.
At 8 h of observation, J3 were found to be, on average, 526 µm long and 38 µm wide; the same was observed at 12 h; however, it was not until 48 h that there was a greater quantity within the analyzed larvae. At 96 h, J3 and J4 were approximately 510 µm long and 37 µm wide. At 120 h, both J4 and adults could be observed, i.e., the first generation, where the females were, on average, 1090 µm long and 78 µm wide; the matricidal endotochy process could be observed inside. Likewise, at 144 h, gravid females were observed that were 1320 µm long and 90 µm wide in addition to some J1, which were observed without visible organs inside and were, on average, 256 µm in length and 22 µm in width. Subsequently, at 168 h, J1, J2 and J3 were observed, which were 250 µm long and 22 µm wide, 386 µm long and 28 µm wide, and 582 µm long and 43 µm wide for each juvenile analyzed, respectively.
Then, we observed mostly J4 and adults, and J4 showed average measurements of 615 µm in length and 40 µm in width. The females analyzed at 216 h were, on average, 906 µm in length and 65 µm in width, and matricidal endotochy was also observed in them. The males were approximately 850 µm in length and 56 µm in width.
To explain this in slightly more detail, we evaluated the measurements of different stages of the EPNs at different times using an optical microscope (Leica DM500, Switzerland) and the Leica Application Suite program, version 3.4.0. Their lengths and widths are shown in Table 2.
Table 2.
The results of the development of O. myriophila, where length and width measurements at the different stages of development were determined (mean ± SD) and the respective n in each case was observed (N/A = nonviable IJs).
3.3. Storage IJs of O. myriophila
When analyzing the IJs stored for 15 days, a significant difference was observed. The IJs that were kept at 8 °C did not survive and exhibited a mortality rate of 100%; on the other hand, the IJs at temperatures of 12, 20 and 27 °C remained the same (p > 0.05), with survival rates of 93.33%, 88% and 85%, respectively.
No significant differences were observed in terms of survival, but at up to 120 days of analysis, survival decreased to 66.67% at 12 °C, 42.67% at 20 °C and 37.33% at 27 °C.
Likewise, at 150 days of storage, the IJs corresponding to temperatures of 20 and 27 °C decreased their survival percentage to 0%, unlike those preserved at 12 °C that maintained a survival rate of 58.67% at 180 days; this is illustrated in Figure 2.
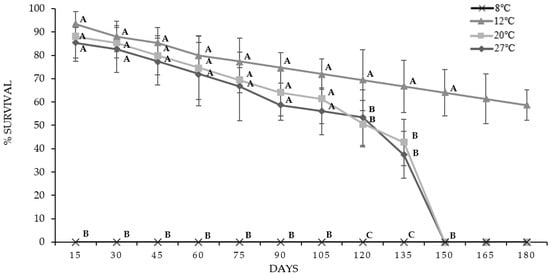
Figure 2.
Survival percentages of IJs from O. myriophila analyzed for 180 days at four different temperatures; the mean ± SD is indicated. The same letters indicate that there are no significant differences (p < 0.05).
The IJs at temperatures of 20 and 27 °C were kept for four and a half months; on the other hand, those kept at 12 °C maintained their survival for more than six months. Of note, the IJs stored at 8 °C had 100% mortality at the first evaluation, which was at day 15.
Pathogenicity of IJs
Pathogenicity was monitored after 30 days; there were no significant differences at the beginning (p > 0.05) because the infectivity percentages at this time were 97.5% at 12 and 20 °C and over 95% at 27 °C.
A significant difference in the evaluation could be observed at up to 90 days, and it was observed that the pathogenicity at 12 and 20 °C was significantly the same; on the other hand, a significant difference was observed between 12 and 27 °C. For the evaluation at 120 days, the pathogenicity was reduced to 20 and 27 °C and remained at 55% and 45%, respectively. Finally, at 180 days, the IJs preserved at 12 °C exhibited both survival and pathogenicity, which decreased at the same time (Table 3).
Table 3.
Percentage rates of pathogenicity of IJs on 10 G. mellonella larvae evaluated over 180 days at different temperatures (n = 10). The values are shown as the mean ± SD. The same letters indicate that there are no significant differences (p < 0.05). (N/A = nonviable IJs).
4. Discussion
In vivo propagation is used to produce EPNs on a laboratory scale and use them in various investigations, as well as to generate the necessary material for field tests [25]. White traps are needed to carry out this method, with good results when obtaining IJs.
According to Sáenz and Luque [26], the first days of emergence are those in which the greatest number of IJs are obtained, at up to 90% of the total production, and in our case, it was between days 6 and 14. The days with the highest production did not present significant differences, but this behavior was observed in the three inocula analyzed.
Likewise, other authors such as Realpe et al. [27] have carried out production studies with G. mellonella larvae using Steinernema colombiense and Heterorhabditis bacteriophora, and they obtained quantities of 86,250 and 78,750 IJs, respectively. Although these are higher figures than those obtained in the present study, averages of 165,000 IJs/larvae have also been recorded for different species, ranging between 30,000 and 240,000 IJs [28], thus approaching the reported yields.
Likewise, Boff and collaborators [29] mentioned that the decrease in IJs in in vivo propagation may be due to different factors, for example, age, size and even the susceptibility of the host insect used, in addition to different conditions, such as the environmental conditions and the inoculum used.
To complement the results obtained in in vivo propagation, it is also important to understand the development of the EPN over time by observing it, with the help of G. mellonella in this case. By observing the development of the EPN, it is known that the life cycle of the steinernematids is on average between 7 and 10 days and that of the heterorhabdithids is between 12 and 15 days [30], and they have between 1 and 2 generations throughout their development, which depend mainly on the availability of resources. Sáenz and Luque [26] monitored Steinernema feltiae, with an average cycle of 8 days. Additionally, the size of the adults was smaller in the second generation, which coincides with species of families other than the one used in this study [31].
To complement the results of the propagation and development of the nematode, its storage evaluation is also important. This is a strong point within the investigation, since there is little information about it, especially when we talk about specific species, such as the one addressed in this investigation. Understanding EPNs in detail is essential, in addition to knowing whether or not the evaluated conditions affect their pathogenicity. According to Kaya and Stock [16], the optimum storage temperatures for steinernematids is 4 to 15 °C for 6 to 9 months, and the heterorhabdithids under similar conditions can only be kept for 3 to 4 months; they also mention that inocula between 1000 and 2000 are ideal for storage and that most species are active at a temperature of 25 ± 2 °C.
In addition, there is information that extreme climates are detrimental to most EPN species, that is, temperatures greater than 30 °C or less than 9 °C, which vary depending on the geographical region where they are found [32].
Each species will withstand different temperatures since species belonging to the same family can persist at different temperatures; for example, Steinernema glaseri can withstand temperatures between 10 and 35 °C because it is a subtropical species. On the other hand, the Steinernema carpocapsae species persists best at temperatures between 5 and 15 °C [32].
Additionally, ecological studies of EPNs have shown that not only species isolates but also isolates within a species of EPNs differ in host range, infectivity, environmental stress tolerance, reproduction and host-finding ability [33,34,35,36,37,38,39].
The species we worked with was isolated from an area where the temperature ranges between 13 and 34 °C throughout the year, and it could not withstand the temperature of 8 °C at which it was evaluated and in which null survival was observed from the beginning of the storage experiment.
Therefore, temperature significantly affects storage because the survival and therefore also the infectivity of the IJs depend on it [40,41].
Author Contributions
Conceptualization, V.M.H.-V.; methodology, V.M.H.-V., G.P.-C., J.A.R.-T. and I.A.-S.; validation, T.M.G.-A. and T.G.R.-O.; formal analysis, I.A.-S. and T.M.G.-A.; data curation, T.M.G.-A.; writing—original draft preparation, T.M.G.-A. and V.M.H.-V.; writing—review and editing, T.M.G.-A., V.M.H.-V. and G.P.-C.; funding acquisition, V.M.H.-V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All data analyzed in this study are included in this article.
Acknowledgments
The authors acknowledge that Conacyt/México provided a scholarship for doctoral studies to T.M.G.A.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shapiro-Ilan, D.; Gaugler, R. Production technology for entomopathogenic nematodes and their bacterial symbionts. J. Ind. Microbiol. Biotechnol. 2002, 28, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Travassos, L. Sobre O Genera Oxystomatium. Boletin Biol. 1927, 5, 20–21. [Google Scholar]
- Poinar, G. Description and biology of a new insect parasitic Rhabditoid, Heterorhabditis bacteriophora N. Gen., N. Sp. (Rhabditida; Heterorhabditidae N. Fam.). Nematologica 1975, 21, 463–470. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Hazir, S.; Glazer, I. Basic and applied research: Entomopathogenic nematodes. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 91–105. [Google Scholar] [CrossRef]
- Picoaga, A.; Abelleira, A.; Mansilla, J.; Areeiro, E. The Entomopathogenic Nematodes and Their Application for Biological Control of Insect Pests. Vitic. Enol. Prof. 2007, 113, 42–46. [Google Scholar]
- Zhang, C.; Liu, J.; Xu, M.; Sun, J.; Yang, S.; An, X.; Gao, G.; Lin, M.; Lai, R.; He, Z.; et al. Heterorhabditidoides chongmingensis gen. nov., sp. nov. (Rhabditida: Rhabditidae), a novel menber of the entomopathogenic nematodes. J. Invertebr. Pathol. 2008, 98, 153–168. [Google Scholar] [CrossRef]
- Abebe, E.; Jumba, M.; Bonner, K.; Gray, V.; Morris, K.; Thomas, W. An entomopathogenic Caenorhabditis briggsae. J. Exp. Biol. 2010, 213, 3223–3229. [Google Scholar] [CrossRef]
- Torrini, G.; Mazza, G.; Carletti, B.; Benvenuti, C.; Roversi, P.; Fanelli, E.; Luca, F.; Troccoli, A.; Tarasco, E.O. onirici sp. n. (Nematoda: Rhabditidae): A new entomopathogenic nematode from an Italian cave. Zootaxa 2015, 3937, 533–548. [Google Scholar] [CrossRef]
- Seenivasan, N.; Sivakumar, M. Screening for environmental stress-tolerant entomopathogenic nematodes virulent against cotton bollworms. Phytoparasitica 2014, 42, 165–177. [Google Scholar] [CrossRef]
- Castro-Ortega, I.; Caspeta-Mandujano, J.; Suárez-Rodríguez, R.; Peña-Chora, G.; Ramírez-Trujillo, J.; Cruz-Pérez, K.; Sosa, I.; Hernández-Velázquez, V. Oscheius myriophila (Nematoda: Rhabditida) isolated in sugar cane soils in Mexico with potential to be used as entomopathogenic nematode. J. Nematol. 2020, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, S.; Xu, M.; Sun, J.; Liu, H.; Liu, J.; Liu, H.; Kan, F.; Sun, J.; Lai, R.; et al. Serratia nematodiphila sp. nov., associated symbiotically with the entomopathogenic nematode Heterorhabditidoides chongmingensis (Rhabditida: Rhabditidae). Int. J. Syst. Evol. Microbiol. 2009, 59, 1603–1608. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Gaugler, R.; Lewis, E. In vivo production of entomopathogenic nematodes. Int. J. Nematol. 2004, 14, 13–18. [Google Scholar]
- Sáenz, A. Importancia de los nemátodos entomopatógenos para el control biológico de plagas en palma de aceite. Palmas 2005, 26, 41–57. [Google Scholar]
- Nguyen, K.; Buss, E. Steinernema phyllophagae n. sp (Rhabditida: Steinernematidae), a new entomopathogenic nematode from Florida, USA. Nematology 2011, 13, 425–442. [Google Scholar] [CrossRef]
- Pilz, C.; Wegensteiner, R.; Keller, S. Natural occurrence of insect pathogenic fungi and insect parasitic nematodes in Diabrotica virgifera virgifera populations. Biocontrol 2008, 53, 353–359. [Google Scholar] [CrossRef]
- Kaya, H.; Stock, S. Techniques in insect nematology. In Manual of Techniques in Insect Pathology; Academic Press: Cambridge, MA, USA, 1997; pp. 281–324. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Escuer, M.; Labrador, S.; Robertson, L.; Barrios, L.; Gutiérrez, C. Distribution of the entomopathogenic nematodes from La Rioja (Northern Spain). J. Invertebr. Pathol. 2007, 95, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Koppenhöfer, A.; Kaya, H. Ecological characterization of Steinernema rarum. J. Invertebr. Pathol. 1999, 73, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Vinciguerra, M.; Clausi, M. Biological control of chestnut insect pests by means of entomopathogenic nematodes. Adv. Hortic. Sci. 2006, 20, 40–44. [Google Scholar]
- Castillo, C.; Gallegos, P.; Oña, M. Manejo de Nemátodos Entomopatógenos en Laboratorio; Instituto Nacional Autónomo de Investigaciones Agropecuarias (INIAP): Quito, Ecuador; Estación Experimental Santa Catalina (EESC): Quito, Ecuador; Departamento Nacional de Protección Vegetal (DNPV): Quito, Ecuador; Panamericana Sur de Quito: Quito, Ecuador, 2011; pp. 48–49. [Google Scholar]
- Woodring, J.; Kaya, H. Steinernematid and Heterorhabditid Nematodes: A Handbook of Techniques; Agricultural Experiment Station Fayetteville: Fayetteville, AR, USA, 1988; Volume 31, pp. 1—130, 249. [Google Scholar]
- Maldonado, G. Manual de Prácticas de Parasitología Con Énfasis en Helmintos Parásitos de Peces de Agua Dulce y Otros Animales Silvestres de México; Universidad Nacional Autónoma de México: México City, Mexico, 2009; pp. 15–18. Available online: http://www.ibiologia.unam.mx (accessed on 9 November 2021).
- Caspeta-Mandujano, J. Nemátodos Parásitos de Peces de Agua Dulce de México: Clave de Identificación, Descripción y Distribución de Las Especies; AGT Editor: Los Angeles, CA, USA, 2010; pp. 3–6. [Google Scholar]
- Acevedo, J.; Cavalcanti, R.; Andaló, V.; Mendonça, L. Efecto de temperatura, concentración y tiempo de almacenamiento en la supervivencia de nemátodos entomopatógenos. Rev. Colomb. Entomol. 2006, 32, 24–30. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Gaugler, R.; Tedders, L.; Brown, I.; Lewis, E. Optimization of inoculation for in vivo production of entomopathogenic nematodes. J. Nematol. 2002, 34, 343–350. [Google Scholar]
- Saenz, A.; Luque, J. Cultivation in vivo and method of storage for infective juveniles of Steinernema feltiae (Rhabdita: Stenernematidae). Agron. Colomb. 2000, 17, 37–42. [Google Scholar]
- Realpe, A.; Bustillo, P.; López, N. Optimización de la cría de Galleria mellonella (L.) para la producción de nematodos entomopatógenos parásitos de la broca del café. Cenicafé 2007, 58, 142–157. [Google Scholar]
- Lindegreen, J.; Valero, K.; Mackey, B. Simple in vivo production and storage methods for Steinernema carpocapsae infective juveniles. J. Nematol. 1993, 25, 193–197. [Google Scholar]
- Boff, M.; Wiegers, G.; Smits, P. Effect of storage time and temperature on infectivity, reproduction and development of Heterorhabditis megidis in Galleria mellonella. Nematology 2000, 2, 635–644. [Google Scholar] [CrossRef]
- Sáenz, A.; Mejia-Torres, M.C. Ecological characterisation of the Colombian entomopathogenic nematode Heterorhabditis sp. SL0708. Braz. J. Biol. 2013, 73, 239–243. [Google Scholar] [CrossRef]
- Çimen, H.; Lee, M.; Hatting, J.; Hazir, S.; Stock, S. Steinernema innovationi n. sp. (Panagrolaimomorpha: Steinernematidae), a new entomopathogenic nematode species from South Africa. J. Helminthol. 2015, 89, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H. Soil Ecology. In Entomopathogenic Nematodes in Biological Control; CRC Press: Boca Raton, FL, USA, 1990; Volume 93. [Google Scholar]
- Gaugler, R.; McGuire, T.; Campbell, J. Genetic variability among strains of the entomopathogenic nematode Steinernema feltiae. J. Nematol. 1989, 21, 247–253. [Google Scholar]
- Somasekhar, N.; Grewal, P.; Klein, M. Genetic variability in stress tolerance and fitness among natural populations of Steinernema carpocapsae. Biol. Control 2002, 23, 303–310. [Google Scholar] [CrossRef]
- Hominick, W. Entomopathogenic rhabditid nematodes and pest-control. Parasitol. Today 1990, 6, 148–152. [Google Scholar] [CrossRef]
- Rosa, J.; Simoes, N. Evaluation of twenty-eight strains of Heterorhabditis bacteriophora isolated in Azores for biocontrol of the armyworm, Pseudaletia unipuncta (Lepidoptera: Noctuidae). Biol. Control 2004, 29, 409–417. [Google Scholar] [CrossRef]
- Morton, A.; Garcia-del-Pino, F. Ecological characterization of entomopathogenic nematodes isolated in stone fruit orchard soils of Mediterranean areas. J. Invertebr. Pathol. 2009, 102, 203–213. [Google Scholar] [CrossRef]
- Glazer, I. Survival biology. In Entomopathogenic Nematology; Gaugler, R., Ed.; CABI: Wallingford, UK, 2002; pp. 169–187. [Google Scholar] [CrossRef]
- Rolston, A.; Meade, C.; Boyle, S.; Kakouli-Duarte, T.; Downes, M. Intraspecific variation among isolates of the entomopathogenic nematode Steinernema feltiae from Bull Island, Ireland. Nematology 2009, 11, 439–451. [Google Scholar]
- Hazir, S.; Stock, S.; Kaya, H.; Koppenhofer, A.; Keskin, N. Developmental temperature effects on five geographic isolates of the entomopathogenic nematode Steinernema feltiae (Nematoda: Steinernematidae). J. Invertebr. Pathol. 2001, 77, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Grewal, P.; Selvan, S.; Gaugler, R. Thermal adaptation of entomopathogenic nematodes -niche breadth for infection, establishment, and reproduction. J. Therm. Biol. 1994, 19, 245–253. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).