

The Genomic Characteristics of an Arthritis-Causing Salmonella pullorum


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. Growth Curves
2.3. Minimum Inhibitory Concentration (MIC) Assay
2.4. RNA Isolation and Quantitative Real-Time PCR (qPCR)
2.5. Extracting and Sequencing Bacterial Genome Library Construction
2.6. Gene Prediction
2.7. Gene Function Annotation
2.8. Genomic Variability Analysis of Arthritis-Causing S. pullorum
2.8.1. ANI Analysis
2.8.2. Constructing the Reference Sequence Index File
2.8.3. Sequence Alignment
2.8.4. Recalibration Base Quality Score (BQSR)
2.8.5. Quality Control and Filtering of Variant Sites
2.8.6. Phylogenetic Tree Construction
2.9. Enrichment of Differential Genes
2.9.1. Screening for Nonsynonymous Mutations
2.9.2. VCF Information Statistics
2.10. Statistical Analysis
3. Results
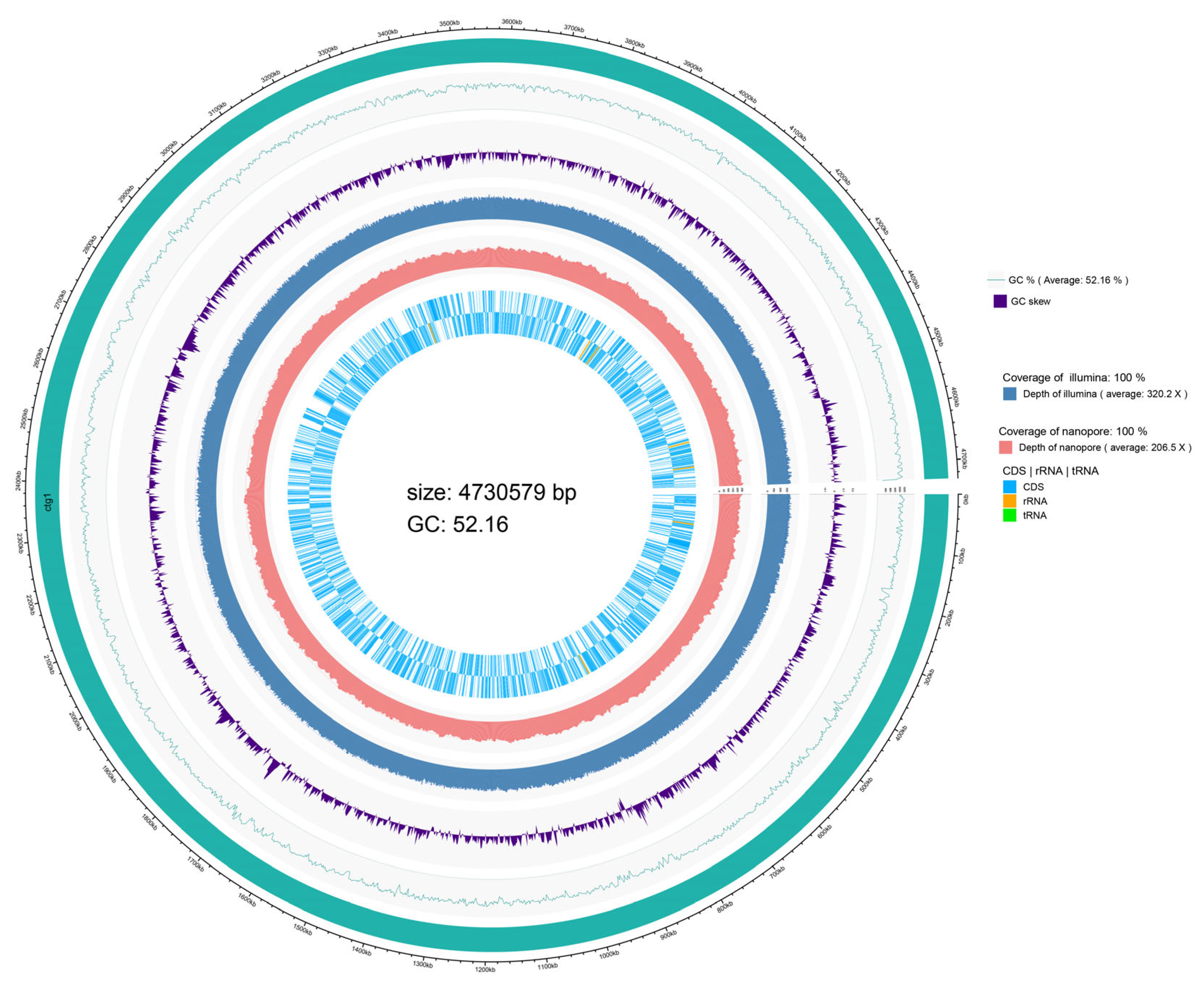
3.1. General Genomics Features of SP206-2
3.2. Genomic Variability Analysis of Arthritis-Causing S. pullorum
3.3. Gene Variations between Arthritis-Causing and Diarrhea-Causing S. pullorum
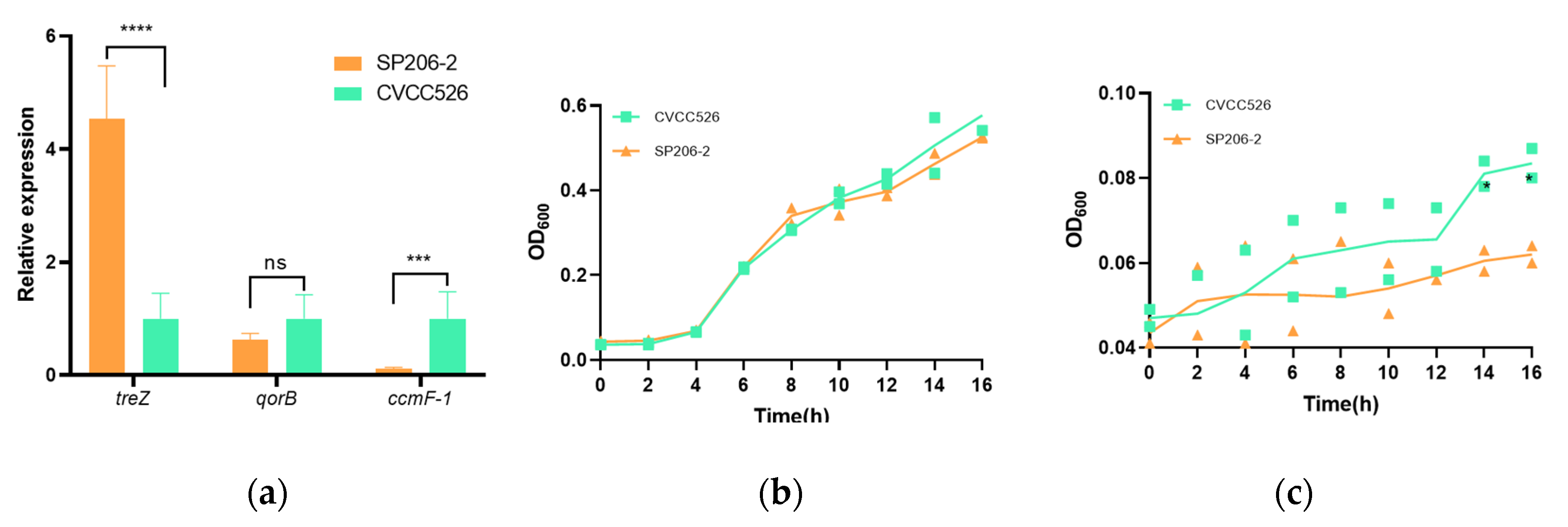
3.4. Relative Expression of Differentially Expressed Genes
3.5. Growth Differences between Arthritis-Causing and Diarrhea-Causing S. pullorum
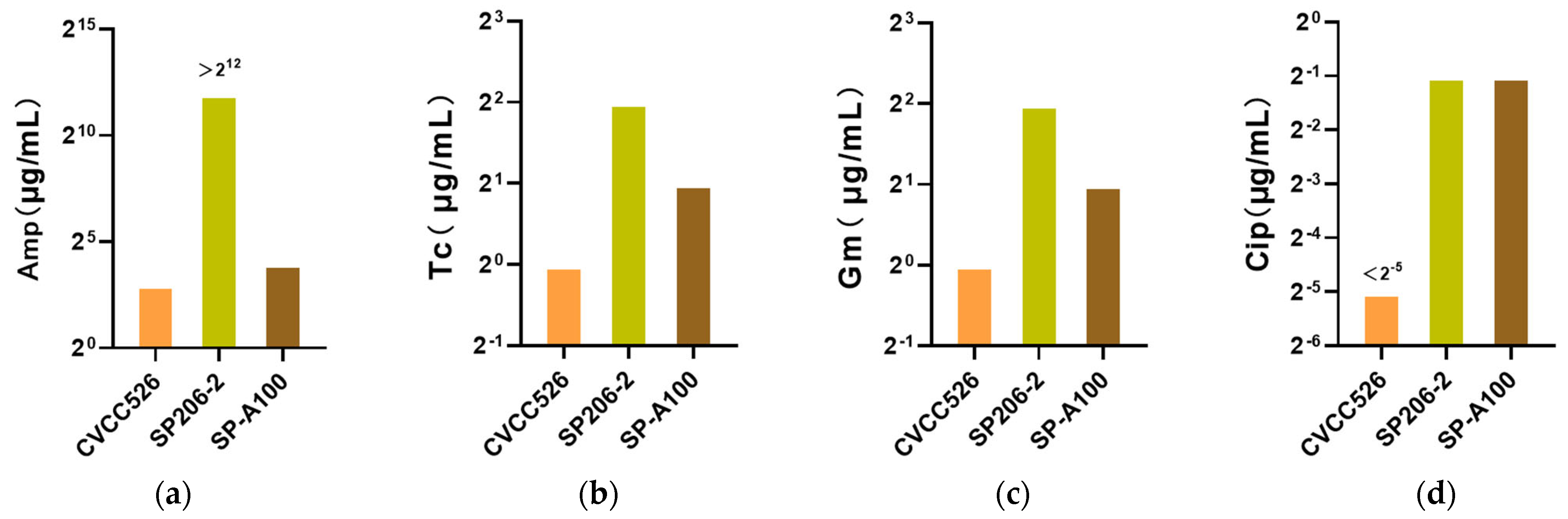
3.6. Drug-Resistant Phenotypic Differences of S. pullorum Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Ghany, W.A.A. Salmonellosis: A food borne zoonotic and public health disease in Egypt. J. Infect. Dev. Ctries. 2020, 14, 674–678. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; El-Mageed, T.A.A.; Soliman, S.M.; Khafaga, A.F.; Swelum, A.A.; Ahmed, A.E.; Alshammari, F.A.; El-Hack, M.E.A. The control of poultry salmonellosis using organic agents: An updated overview. Poult. Sci. 2022, 101, 101716. [Google Scholar] [CrossRef] [PubMed]
- Bumstead, N.; Barrow, P. Resistance to Salmonella gallinarum, S. pullorum, and S. enteritidis in inbred lines of chickens. Avian Dis. 1993, 37, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H.L. Fowl typhoid and pullorum disease. Rev. Sci. Technol. 2000, 19, 405–424. [Google Scholar] [CrossRef] [PubMed]
- Barrow, P.A.; Neto, O.C.F. Pullorum disease and fowl typhoid—New thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef]
- Schat, K.A.; Nagaraja, K.V.; Saif, Y.M. Pullorum Disease: Evolution of the Eradication Strategy. Avian Dis. 2021, 65, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Li, Z.; Zhou, X.; Huang, C.; Hu, Y.; Geng, S.; Chen, X.; Li, Q.; Pan, Z.; Jiao, X. Induction of arthritis in chickens by infection with novel virulent Salmonella Pullorum strains. Vet. Microbiol. 2019, 228, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Kang, X.; Zhou, K.; Yue, M. A global dataset for prevalence of Salmonella Gallinarum between 1945 and 2021. Sci. Data 2022, 9, 495. [Google Scholar] [CrossRef]
- Braga, J.F.V.; Chanteloup, N.K.; Trotereau, A.; Baucheron, S.; Guabiraba, R.; Ecco, R.; Schouler, C. Diversity of Escherichia coli strains involved in vertebral osteomyelitis and arthritis in broilers in Brazil. BMC Vet. Res. 2016, 12, 140. [Google Scholar] [CrossRef]
- Mosleh, N.; Shomali, T.; Namazi, F.; Marzban, M.; Mohammadi, M.; Boroojeni, A.M. Comparative evaluation of therapeutic efficacy of sulfadiazine-trimethoprim, oxytetracycline, enrofloxacin and florfenicol on Staphylococcus aureus-induced arthritis in broilers. Br. Poult. Sci. 2016, 57, 179–184. [Google Scholar] [CrossRef]
- Liu, R.; Xu, B.; Zhang, J.; Sun, H.; Liu, C.; Lu, F.; Pan, Q.; Zhang, X. Mycoplasma synoviae induces serum amyloid A upregulation and promotes chicken synovial fibroblast cell proliferation. Microb. Pathog. 2021, 154, 104829. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, T.; Wang, W.; Xie, Q.; Wan, Z.; Qin, A.; Ye, J.; Shao, H. Isolation and Molecular Characteristics of a Novel Recombinant Avian Orthoreovirus from Chickens in China. Front. Vet. Sci. 2021, 8, 771755. [Google Scholar] [CrossRef]
- Landman, W.J.; Veldman, K.T.; Mevius, D.J.; van Eck, J.H. Investigations of Enterococcus faecalis-induced bacteraemia in brown layer pullets through different inoculation routes in relation to the production of arthritis. Avian Pathol. 2003, 32, 463–471. [Google Scholar] [CrossRef]
- Borst, L.B.; Suyemoto, M.M.; Keelara, S.; Dunningan, S.E.; Guy, J.S.; Barnes, H.J. A chicken embryo lethality assay for pathogenic Enterococcus cecorum. Avian Dis. 2014, 58, 244–248. [Google Scholar] [CrossRef]
- Alpigiani, I.; Abrahantes, J.C.; Michel, V.; Huneau-Salaun, A.; Chemaly, M.; Keeling, L.J.; Gervelmeyer, A.; Bacci, C.; Brindani, F.; Bonardi, S.; et al. Associations between animal welfare indicators and Campylobacter spp. in broiler chickens under commercial settings: A case study. Prev. Vet. Med. 2017, 147, 186–193. [Google Scholar] [CrossRef]
- Sexton, T.Y.; Geornaras, I.; Belk, K.E.; Bunning, M.; Martin, J.N. Salmonella Contamination in Broiler Synovial Fluid: Are We Missing a Potential Reservoir? J. Food Prot. 2018, 81, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Kang, M.S.; An, B.K.; Song, E.A.; Kwon, J.H.; Kwon, Y.K. Occurrence of purulent arthritis broilers vertically infected with Salmonella enterica serovar Enteritidis in Korea. Poult. Sci. 2010, 89, 2116–2122. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Johnston, R.N.; Liu, G.R.; Liu, S.L. Genomic comparison between Salmonella Gallinarum and Pullorum: Differential pseudogene formation under common host restriction. PLoS ONE 2013, 8, e59427. [Google Scholar] [CrossRef]
- Li, P.; Zhang, M.; Hao, G.; Sun, S. Research Note: Hypervirulent arthritis-causing Salmonella Pullorum isolated from Chinese native chicken breeds significantly decreased growth performance of chicks. Poult. Sci. 2022, 101, 101575. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests, 13th ed.; CLSI Supplement M02; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Laslett, D.; Canback, B. ARAGORN, a program to detect tRNA genes and tmRNA genes in nucleotide sequences. Nucleic Acids Res. 2004, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rodland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, D.L.; Eddy, S.R. Fast filtering for RNA homology search. Bioinformatics 2011, 27, 3102–3109. [Google Scholar] [CrossRef] [PubMed]
- Wigley, P.; Hulme, S.D.; Powers, C.; Beal, R.K.; Berchieri, A., Jr.; Smith, A.; Barrow, P. Infection of the reproductive tract and eggs with Salmonella enterica serovar pullorum in the chicken is associated with suppression of cellular immunity at sexual maturity. Infect. Immun. 2005, 73, 2986–2990. [Google Scholar] [CrossRef] [PubMed]
- Berchieri, A., Jr.; Murphy, C.K.; Marston, K.; Barrow, P.A. Observations on the persistence and vertical transmission of Salmonella enterica serovars Pullorum and Gallinarum in chickens: Effect of bacterial and host genetic background. Avian Pathol. 2001, 30, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, Z.; Qiang, B.; Xu, Y.; Chen, X.; Li, Q.; Jiao, X. Loss and Gain in the Evolution of the Salmonella enterica Serovar Gallinarum Biovar Pullorum Genome. mSphere 2019, 4, e00627-18. [Google Scholar] [CrossRef] [PubMed]
- Ley, T.J.; Mardis, E.R.; Ding, L.; Fulton, B.; McLellan, M.D.; Chen, K.; Dooling, D.; Dunford-Shore, B.H.; McGrath, S.; Hickenbotham, M.; et al. DNA sequencing of a cytogenetically normal acute myeloid leukaemia genome. Nature 2008, 456, 66–72. [Google Scholar] [CrossRef]
- Lupkowska, A.; Monem, S.; Debski, J.; Stojowska-Swedrzynska, K.; Kuczynska-Wisnik, D.; Laskowska, E. Protein aggregation and glycation in Escherichia coli exposed to desiccation-rehydration stress. Microbiol. Res. 2023, 270, 127335. [Google Scholar] [CrossRef]
- Zhang, B.; Yao, H.; Qi, H.; Zhang, X.L. Trehalose and alginate oligosaccharides increase the stability of muscle proteins in frozen shrimp (Litopenaeus vannamei). Food Funct. 2020, 11, 1270–1278. [Google Scholar] [CrossRef]
- Kim, I.K.; Yim, H.S.; Kim, M.K.; Kim, D.W.; Kim, Y.M.; Cha, S.S.; Kang, S.O. Crystal structure of a new type of NADPH-dependent quinone oxidoreductase (QOR2) from Escherichia coli. J. Mol. Biol. 2008, 379, 372–384. [Google Scholar] [CrossRef]
- Ascenzi, P.; Coletta, M.; Wilson, M.T.; Fiorucci, L.; Marino, M.; Polticelli, F.; Sinibaldi, F.; Santucci, R. Cardiolipin-cytochrome c complex: Switching cytochrome c from an electron-transfer shuttle to a myoglobin- and a peroxidase-like heme-protein. IUBMB Life 2015, 67, 98–109. [Google Scholar] [CrossRef]
- Verbrugghe, E.; Van Parys, A.; Haesendonck, R.; Leyman, B.; Boyen, F.; Haesebrouck, F.; Pasmans, F. Subtherapeutic tetracycline concentrations aggravate Salmonella Typhimuriuminfection by increasing bacterial virulence. J. Antimicrob. Chemother. 2016, 71, 2158–2166. [Google Scholar] [CrossRef]
- Dawan, J.; Kim, S.; Ahn, J. Assessment of phenotypic heterogeneity in Salmonella Typhimurium preadapted to ciprofloxacin and tetracycline. FEMS Microbiol. Lett. 2023, 370, fnad100. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Fragment Size (bp) | Annotation |
|---|---|---|---|
| treZ-L treZ-R | cgatccggcttcccgcgcccccagccacgctcgccgccaa | 259 | Trehalose hydrolase |
| qorB-L qorB-R | agcgcgcattgcggaggcgagcaccgcgcagcaccgcaac | 278 | NAD(P)H dehydrogenase |
| ccmF-1-L ccmF-1-R | gcgggcgcgggagccgtatcggcgcgcccaccgaaagcct | 129 | Cytochrome C synthetase |
| 16S rRNA-L 16S rRNA-R | cagggggccgccttcgccacagcgcacgcaggcggtctgt | 168 | Reference genome |
| Type | Number | Total Length (bp) | Average Length (bp) |
|---|---|---|---|
| Gene | 5007 | 4,306,080 | 860 |
| CDS | 4729 | 4,240,824 | 897 |
| tRNA | 75 | 5907 | 79 |
| 23S rRNA | 7 | 20,909 | 2987 |
| 16S rRNA | 7 | 10,710 | 1530 |
| 5S rRNA | 8 | 920 | 115 |
| tmRNA | 1 | 363 | 363 |
| Misc RNA | 180 | 26,447 | 147 |
| Pesudogene | 1426 | 391,080 | 274.25 |
| Gene | Mutation Type | Mutation Site | Amino Acid Mutation | Encoded Protein |
|---|---|---|---|---|
| mdtH | Disruptive inframe deletion | 656~673-bit deletion GCTACTATATGCTGGCGG | Gly219~Ala224 deletion | Norfloxacin resistant protein MdtH |
| tagO | Conservative inframe deletion | 469~507 -bit deletion CACGCCATGACGCCGGACAGCATCGACGTGACGCTGACC | His157~Thr169 deletion | Type VI secretory system associated proteins TagO |
| rbsC | Disruptive inframe deletion | 264~308-bit deletion TTCAATCGTCGGCGTTGAAGTGAATGCGCTGGTCGCGGTTGCCGC | Ser89~Ala103 deletion | Ribose transporter RbsC |
| sptP | Frameshift variant & stop gained | 369~370 sequence deletion TGTTGCGCCTGAAAAATTTTCGTCAAAAGTATTAACCTGGCTTGGAAAAATGCCGTTATTTAAAAACACTG | Lys124 termination of translation | Secretory effector protein SptP |
| thrL | Disruptive inframe deletion | 33~47-bit deletion CATCACCATTACCAC | Ile12~Thr16 deletion | Threonine operon precursor peptide |
| guaB | Conservative inframe deletion | 1447~1449-bit deletion AAC | Asn483 deletion | Inosine monophosphate dehydrogenase |
| hpaI | Frameshift variant | 362~378-bit deletion TGCTGGCGCGGGCATCG | Val121 termination of translation | P-hydroxyphenylacetic aldolase |
| ccmF-1 | Disruptive inframe insertion | 1039~1040sequence deletion GTTTGGGCGGCGGAAAGTGCGAAACGCTCATTCTGGCGACGC | Thr346~Leu347 deletion | Cytochrome C synthetase CcmF subunit |
| qorB | Frameshift variant | 648~649-bit deletion AA | Ser217 termination of translation | Quinone oxidoreductase |
| treZ | Frameshift variant | 3~4-bit deletion TT | Val2 termination of translation | mahosyhrehalose hydrolase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Z.; Huang, J.; Li, P.; Song, M.; Liu, B.; Tang, W.; Sun, S. The Genomic Characteristics of an Arthritis-Causing Salmonella pullorum. Microorganisms 2023, 11, 2986. https://doi.org/10.3390/microorganisms11122986
Lu Z, Huang J, Li P, Song M, Liu B, Tang W, Sun S. The Genomic Characteristics of an Arthritis-Causing Salmonella pullorum. Microorganisms. 2023; 11(12):2986. https://doi.org/10.3390/microorganisms11122986
Chicago/Turabian StyleLu, Zhiyuan, Jiaqi Huang, Peiyong Li, Mengze Song, Ben Liu, Wenli Tang, and Shuhong Sun. 2023. "The Genomic Characteristics of an Arthritis-Causing Salmonella pullorum" Microorganisms 11, no. 12: 2986. https://doi.org/10.3390/microorganisms11122986
APA StyleLu, Z., Huang, J., Li, P., Song, M., Liu, B., Tang, W., & Sun, S. (2023). The Genomic Characteristics of an Arthritis-Causing Salmonella pullorum. Microorganisms, 11(12), 2986. https://doi.org/10.3390/microorganisms11122986