
Isolation and Evaluation of Streptomyces melanogenes YBS22 with Potential Application for Biocontrol of Rice Blast Disease

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Phenotypic Characteristics of the Strain YBS22
2.2. Library Construction and Genome Sequencing
2.3. Genome Assembly and Annotation
2.4. Taxonomy of the Strain YBS22
2.5. Antifungal Activity of the Strain YBS22 and Its Active Compounds
2.6. Extraction, Purification, and Identification of the Antifungal Active Compounds
2.7. Conidial Germination Inhibition Assays
2.8. Statistical Analysis
3. Results
3.1. Identification and Phenotypic Characteristics of the Strain YBS22
3.2. Genomic Characteristics of YBS22
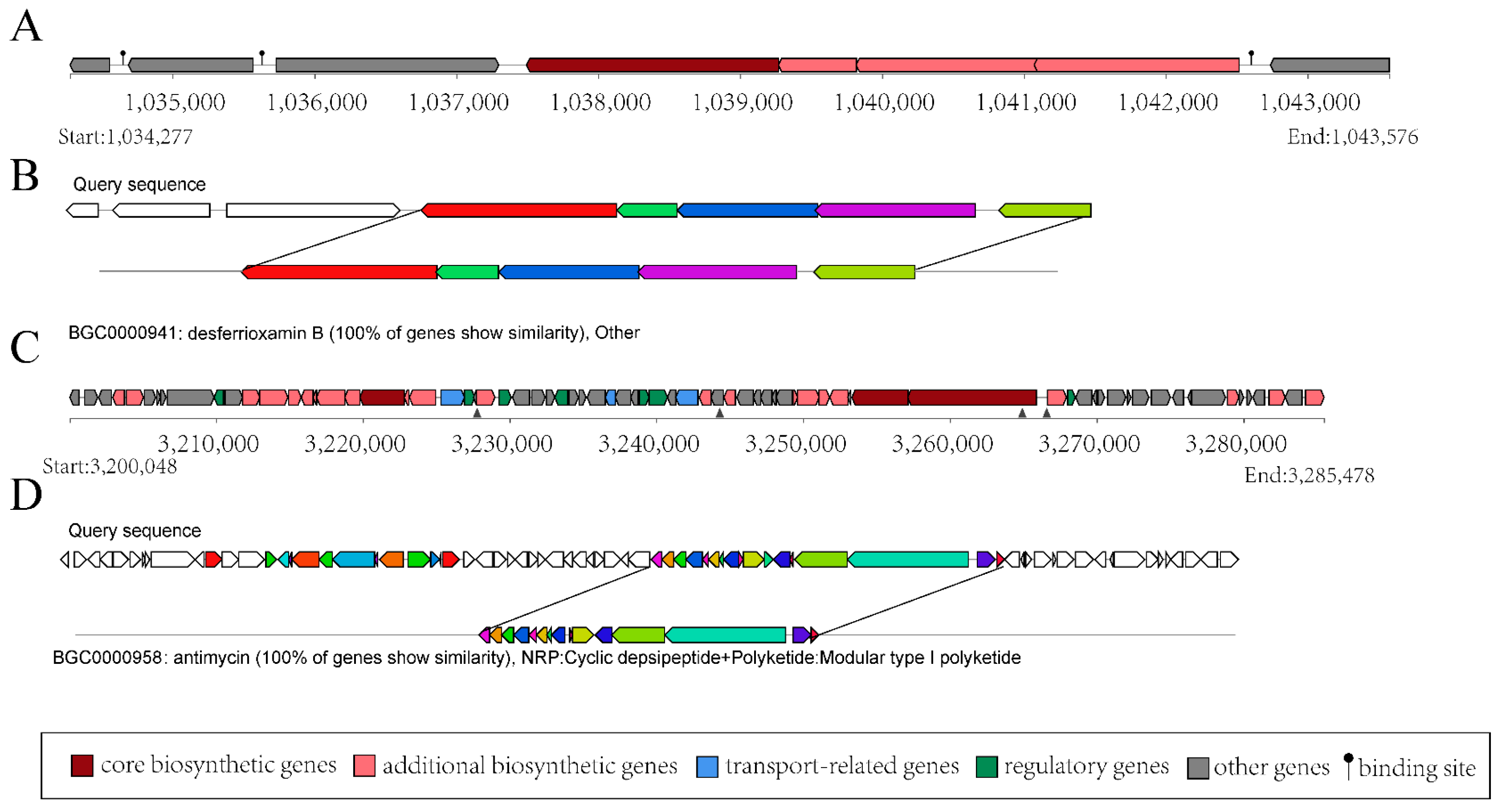
3.3. Genes/Gene Clusters for Antibiotic Synthesis
3.4. Biocontrol Potential of the Strain YBS22
3.5. Preliminary Characterization of the Active Metabolites of YBS22
3.6. Inhibitory Activity of Compound 1 on Spore Germination and Development of M. oryzae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jayachandra, Y.A.; Kontro, M.H.; Ganachari, S.V. Actinobacteria: Ecology, Diversity, Classification and Extensive Applications. Rhizosphere Biology (RHBIO); Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Nawani, N.; Aigle, B.; Mandal, A.; Bodas, M.; Ghorbel, S.; Prakash, D. Actinomycetes: Role in Biotechnology and Medicine. BioMed Res. Int. 2013, 2013, 687190. [Google Scholar] [CrossRef]
- Chaurasia, A.; Meena, B.R.; Tripathi, A.N.; Pandey, K.K.; Rai, A.B.; Singh, B. Actinomycetes: An unexplored microorganisms for plant growth promotion and biocontrol in vegetable crops. World J. Microbiol. Biotechnol. 2018, 34, 132. [Google Scholar] [CrossRef] [PubMed]
- Le, K.D.; Yu, N.H.; Park, A.R.; Park, D.-J.; Kim, C.-J.; Kim, J.-C. Streptomyces sp. AN090126 as a Biocontrol Agent against Bacterial and Fungal Plant Diseases. Microorganisms 2022, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gan, M.; Dong, B.; Zhang, T.; Li, Y.; Zhang, Y.; Fan, X.; Wu, Y.; Bai, S.; Chen, M.; et al. 4862f, a new inhibitor of Hiv-1 protease, from the culture of Streptomyces I03a-04862. Molecules 2012, 18, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Karthik, N.; Binod, P.; Pandey, A. Purification and characterisation of an acidic and antifungal chitinase produced by a Streptomyces sp. Bioresour. Technol. 2015, 188, 195–201. [Google Scholar] [CrossRef]
- Ventorino, V.; Ionata, E.; Birolo, L.; Montella, S.; Marcolongo, L.; de Chiaro, A.; Espresso, F.; Faraco, V.; Pepe, O. Lignocellulose-Adapted Endo-Cellulase Producing Streptomyces Strains for Bioconversion of Cellulose-Based Materials. Front. Microbiol. 2016, 7, 2061. [Google Scholar] [CrossRef]
- Kubiak, K.; Wrzosek, M.; Przemieniecki, S.; Damszel, M.; Sierota, Z. Bacteria inhabiting wood of roots and stumps in forest and arable soils. For. Sci. 2018, 86, 319–342. [Google Scholar]
- Jones, S.E.; Elliot, M.A. Streptomyces Exploration: Competition, Volatile Communication and New Bacterial Behaviours. Trends Microbiol. 2017, 25, 522–531. [Google Scholar] [CrossRef]
- LeBlanc, N. Bacteria in the genus Streptomyces are effective biological control agents for management of fungal plant pathogens: A meta-analysis. BioControl 2021, 67, 111–121. [Google Scholar] [CrossRef]
- Mun, B.-G.; Lee, W.-H.; Kang, S.-M.; Lee, S.-U.; Lee, S.-M.; Lee, D.Y.; Shahid, M.; Yun, B.-W.; Lee, I.-J. Streptomyces sp. LH 4 promotes plant growth and resistance against Sclerotinia sclerotiorum in cucumber via modulation of enzymatic and defense pathways. Plant Soil 2020, 448, 87–103. [Google Scholar] [CrossRef]
- Li, X.; Lang, D.; Wang, J.; Zhang, W.; Zhang, X. Plant-beneficial Streptomyces dioscori SF1 potential biocontrol and plant growth promotion in saline soil within the arid and semi-arid areas. Environ. Sci. Pollut. Res. 2023, 30, 70194–70212. [Google Scholar] [CrossRef]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Pooja, K.; Katoch, A. Past, present and future of rice blast management. Plant Sci. Today 2014, 1, 165–173. [Google Scholar] [CrossRef]
- Hollomon, W.D. (Ed.) Fungicide resistance: 40 years on and still a major problem. In Fungicide Resistance in Plant Pathogens: Principles and a Guide to Practical Management; Springer: Tokyo, Japan, 2015. [Google Scholar]
- Fonseca, M.I.; Shimizu, E.; Zapata, P.D.; Villalba, L.L. Copper inducing effect on laccase production of white rot fungi native from Misiones (Argentina). Enzym. Microb. Technol. 2010, 46, 534–539. [Google Scholar] [CrossRef]
- Shi, L.; Nwet, T.T.; Ge, B.; Zhao, W.; Liu, B.; Cui, H.; Zhang, K. Antifungal and plant growth-promoting activities of Streptomyces roseoflavus strain NKZ-259. Biol. Control 2018, 125, 57–64. [Google Scholar] [CrossRef]
- Qi, D.; Zou, L.; Zhou, D.; Chen, Y.; Gao, Z.; Feng, R.; Zhang, M.; Li, K.; Xie, J.; Wang, W. Taxonomy and broad-spectrum antifungal activity of Streptomyces sp. Sca3-4 isolated from rhizosphere soil of opuntia stricta. Front. Microbiol. 2019, 10, 1390. [Google Scholar] [CrossRef] [PubMed]
- Law, J.W.F.; Ser, H.L.; Khan, T.M.; Chuah, L.H.; Pusparajah, P.; Chan, K.G.; Goh, B.H.; Lee, L.H. The potential of Streptomyces as biocontrol agents against the rice blast fungus, Magnaporthe oryzae (Pyricularia oryzae). Front. Microbiol. 2017, 8, 3. [Google Scholar] [CrossRef]
- Xu, T.; Li, Y.; Zeng, X.; Yang, X.; Yang, Y.; Yuan, S.; Hu, X.; Zeng, J.; Wang, Z.; Liu, Q.; et al. Isolation and evaluation of endophytic Streptomyces endus OsiSh-2 with potential application for biocontrol of rice blast disease. J. Sci. Food Agric. 2017, 97, 1149–1157. [Google Scholar] [CrossRef] [PubMed]
- Awla, H.K.; Kadir, J.; Othman, R.; Rashid, T.S.; Hamid, S.; Wong, M.Y. Plant growth-promoting abilities and biocontrol efficacy of Streptomyces sp. UPMRS4 against Pyricularia oryzae. Biol. Control 2017, 112, 55–63. [Google Scholar] [CrossRef]
- Chen, X.; Hu, L.-F.; Huang, X.-S.; Zhao, L.-X.; Miao, C.-P.; Chen, Y.-W.; Xu, L.-H.; Han, L.; Li, Y.-Q. Isolation and Characterization of New Phenazine Metabolites with Antifungal Activity against Root-Rot Pathogens of Panax notoginseng from Streptomyces. J. Agric. Food Chem. 2019, 67, 11403–11407. [Google Scholar] [CrossRef] [PubMed]
- Ondrejíčková, P.; Šturdíková, M.; Hushegyi, A.; Švajdlenka, E.; Markošová, K.; Čertík, M. Endophytic Streptomyces sp. Ac35, a producer of bioactive isoflavone aglycones and antimycins. J. Ind. Microbiol. Biotechnol. 2016, 43, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Fu, S.N.; Bao, Y.X.; Yang, Y.; Shen, H.F.; Lin, B.R.; Zhou, G.X. Kitamycin C, a new antimycin-type antibiotic from Streptomyces antibioticus strain 200-09. Nat. Prod. Res. 2017, 31, 1819–1824. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Xu, Y.; McConnell, O.; Liu, L.; Li, Y.; Qi, S.; Huang, X.; Qian, P. Two Antimycin a Analogues from Marine-Derived Actinomycete Streptomyces lusitanus. Mar. Drugs 2012, 10, 668–676. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Liu, H.Y.; Chen, J.; Yuan, L.J.; Sun, W.; Zhang, L.X.; Zhang, Y.Q.; Yu, L.Y.; Li, W.J. Diversity of culturable actinobacteria from Qinghai-Tibet Plateau, China. Antonie Van Leeuwenhoek 2010, 98, 213–223. [Google Scholar] [CrossRef]
- Kunova, A.; Bonaldi, M.; Saracchi, M.; Pizzatti, C.; Chen, X.; Cortesi, P. Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 2016, 16, 272. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Besemer, J.; Borodovsky, M. Genemark: Web software for gene finding in prokaryotes, eukaryotes and viruses. Nucleic Acids Res. 2005, 33 (Suppl. S2), W451–W454. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef]
- Medema, M.H.; Blin, K.; Cimermancic, P.; De Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The Silva ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16s rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 2, 346–351. [Google Scholar] [CrossRef]
- Wang, H.; Wen, K.; Zhao, X.; Wang, X.; Li, A.; Hong, H. The inhibitory activity of endophytic Bacillus sp. strain CHM1 against plant pathogenic fungi and its plant growth-promoting effect. Crop Prot. 2009, 28, 634–639. [Google Scholar] [CrossRef]
- Ando, S.; Tanabe, S.; Akimoto-Tomiyama, C.; Nishizawa, Y.; Minami, E. The Supernatant of a Conidial Suspension of Magnaporthe oryzae Contains a Factor that Promotes the Infection of Rice Plants. J. Phytopathol. 2009, 157, 420–426. [Google Scholar] [CrossRef]
- Seo, Y.-W.; Cho, K.-W.; Rho, J.-R.; Mo, S.-J.; Shin, J.-H. A new analog of antimycin from Streptomyces sp. M03033. J. Microbiol. Biotechnol. 2001, 11, 663–667. [Google Scholar]
- Shaikh, A.; Mullowney, M.; O’Hainmhire, E.; Colunga-Hernandez, D.; Burdette, J.; Murphy, B. A New Antimycin Analog from Marine-Derived Streptomyces sp. Planta Med. 2013, 79, PK1. [Google Scholar] [CrossRef]
- Arai, T.; Mikami, Y. Chromogenicity of Streptomyces. Appl. Environ. Microbiol. 1972, 23, 402–406. [Google Scholar] [CrossRef]
- Gulyas, F. Study of the formation of melanoid pigments by some actinomycetes and microscopic fungi. Symp. Biol. Hung. 1972, 11, 153–158. [Google Scholar]
- Coelho, R.R.R.; Linhares, L.F. Melanogenic actinomycetes (Streptomyces spp.) from Brazilian soils. Biol. Fertil. Soils 1993, 15, 220–224. [Google Scholar] [CrossRef]
- Muangham, S.; Pathom-Aree, W.; Duangmal, K. Melanogenic actinomycetes from rhizosphere soil—Antagonistic activity against Xanthomonas oryzae and plant-growth-promoting traits. Can. J. Microbiol. 2015, 61, 164–170. [Google Scholar] [CrossRef]
- Liu, C.; Sheng, J.; Chen, L.; Zheng, Y.; Lee, D.Y.W.; Yang, Y.; Xu, M.; Shen, L. Biocontrol Activity of Bacillus subtilis Isolated from Agaricus bisporus Mushroom Compost against Pathogenic Fungi. J. Agric. Food Chem. 2015, 63, 6009–6018. [Google Scholar] [CrossRef]
- Tourova, T.P. Copy Number of Ribosomal Operons in Prokaryotes and Its Effect on Phylogenetic Analyses. Microbiology 2003, 72, 389–402. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Intragenomic heterogeneity between multiple 16S ribosomal RNA operons in sequenced bacterial genomes. FEMS Microbiol. Lett. 2003, 228, 45–49. [Google Scholar] [CrossRef]
- Engene, N.; Gerwick, W.H. Intra-genomic 16s rrna gene heterogeneity in cyanobacterial genomes. Fottea 2011, 11, 17–24. [Google Scholar] [CrossRef]
- Guo, Y.; Zheng, W.; Rong, X.; Huang, Y. A multilocus phylogeny of the Streptomyces griseus 16S rRNA gene clade: Use of multilocus sequence analysis for streptomycete systematics. Int. J. Syst. Evol. Microbiol. 2008, 58, 149–159. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yuan, J.; E, Y.; Raza, W.; Shen, Q.; Huang, Q. Effects of volatile organic compounds from Streptomyces albulus NJZJSA2 on growth of two fungal pathogens. J. Basic Microbiol. 2015, 55, 1104–1117. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Suwannarach, N.; Lumyong, S.; Ito, S.-I.; Matsui, K.; Sunpapao, A. Streptomyces angustmyceticus Nr8-2 as a Potential microorganism for the biological control of leaf spots of Brassica rapa subsp. pekinensis caused by Colletotrichum sp. and Curvularia lunata. Biol. Control 2019, 138, 104046. [Google Scholar] [CrossRef]
- Nguyen, X.H.; Naing, K.W.; Lee, Y.S.; Kim, Y.H.; Moon, J.H.; Kim, K.Y. Antagonism of antifungal metabolites from Streptomyces griseus H7602 against Phytophthora capsici. J. Basic Microbiol. 2014, 55, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Selim, M.S.M.; Abdelhamid, S.A.; Mohamed, S.S. Secondary metabolites and biodiversity of actinomycetes. J. Genet. Eng. Biotechnol. 2021, 19, 72. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Dangi, P.; Choudhary, M. Actinomycetes: Source, identification, and their applications. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 801–832. [Google Scholar]
- Hussain, A.A.; Mostafa, S.A.; Ghazal, S.A.; Ibrahim, S.Y. Studies on antifungal antibiotic and bioinsecticidal activities of some actinomycete isolates. Afr. J. Mycol. Biotechnol. 2002, 10, 63–80. [Google Scholar]
- Powell, L.C.; Adams, J.Y.; Quoraishi, S.; Py, C.; Oger, A.; Gazze, S.A.; Francis, L.W.; von Ruhland, C.; Owens, D.; Rye, P.D.; et al. Alginate oligosaccharides enhance the antifungal activity of nystatin against candidal biofilms. Cell. Infect. Microbiol. 2023, 13, 1122340. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Chaudhary, S.; Sareen, D. Non-ribosomal peptide synthetases: Identifying the cryptic gene clusters and decoding the natural product. J. Biosci. 2017, 42, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Pang, F.; Solanki, M.K.; Wang, Z. Streptomyces can be an excellent plant growth manager. World J. Microbiol. Biotechnol. 2022, 38, 193. [Google Scholar] [CrossRef]
- Hosotani, N.; Kumagai, K.; Nakagawa, H.; Shimatani, T.; Saji, I. Antimycins A10~A16, Seven New Antimycin Antibiotics Produced by Streptomyces spp. SPA-10191 and SPA-8893. J. Antibiot. 2005, 58, 460–467. [Google Scholar] [CrossRef]
- Yao, C.F.; Schiebel, M.; Helmke, E.; Anke, H.; Laatsch, H. Prefluostatin and new urauchimycin derivatives produced by streptomycete isolates. J. Biosci. 2006, 61, 320–325. [Google Scholar] [CrossRef]
- Balkovec, J.M.; Black, R.M.; Abruzzo, G.K.; Bartizal, K.; Dreikorn, S.; Karl, N. Pneumocandin antifungal lipopeptides. The phenolic hydroxyl is required for 1,3-β-glucan synthesis inhibition. Bioorg. Med. Chem. Lett. 1993, 3, 2039–2042. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | 4 h | 8 h | ||
|---|---|---|---|---|
| Germination Rate (%) | Germ Tube Length (μm) | Germination Rate (%) | Germ Tube Length (μm) | |
| Mock | 90.00 | 51.5 ± 6.5 a | 100.00 | 107.8 ± 8.8 a |
| 50 μg/mL | 0.00 | - | 0.00 | - |
| 25 μg/mL | 9.76 | 6.8 ± 2.2 b | 68.42 | 47.2 ± 12.2 c |
| 5 μg/mL | 70.59 | 42.7 ± 4.7 c | 100.00 | 73.8 ± 8.8 d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, L.; Wang, F.; Liu, C.; Guan, Z.; Wang, M.; Zhong, R.; Xi, H.; Zhao, Y.; Wen, C. Isolation and Evaluation of Streptomyces melanogenes YBS22 with Potential Application for Biocontrol of Rice Blast Disease. Microorganisms 2023, 11, 2988. https://doi.org/10.3390/microorganisms11122988
Song L, Wang F, Liu C, Guan Z, Wang M, Zhong R, Xi H, Zhao Y, Wen C. Isolation and Evaluation of Streptomyces melanogenes YBS22 with Potential Application for Biocontrol of Rice Blast Disease. Microorganisms. 2023; 11(12):2988. https://doi.org/10.3390/microorganisms11122988
Chicago/Turabian StyleSong, Luyang, Fei Wang, Chuang Liu, Zhengzhe Guan, Mengjiao Wang, Rongrong Zhong, Huijun Xi, Ying Zhao, and Caiyi Wen. 2023. "Isolation and Evaluation of Streptomyces melanogenes YBS22 with Potential Application for Biocontrol of Rice Blast Disease" Microorganisms 11, no. 12: 2988. https://doi.org/10.3390/microorganisms11122988
APA StyleSong, L., Wang, F., Liu, C., Guan, Z., Wang, M., Zhong, R., Xi, H., Zhao, Y., & Wen, C. (2023). Isolation and Evaluation of Streptomyces melanogenes YBS22 with Potential Application for Biocontrol of Rice Blast Disease. Microorganisms, 11(12), 2988. https://doi.org/10.3390/microorganisms11122988