
Evaluation of Shifts of Gene Transcription Levels of Unicellular Green Alga Chlamydomonas reinhardtii Due to UV-C Irradiation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgal Strain and Culturing Condition
2.2. Cell Sterilization with UV-C Irradiation
2.3. Evaluation of Lipid Composition and Quantity in Cells with Gas Chromatography
2.4. Measurement of Levels of Gene Translation
3. Results and Discussion
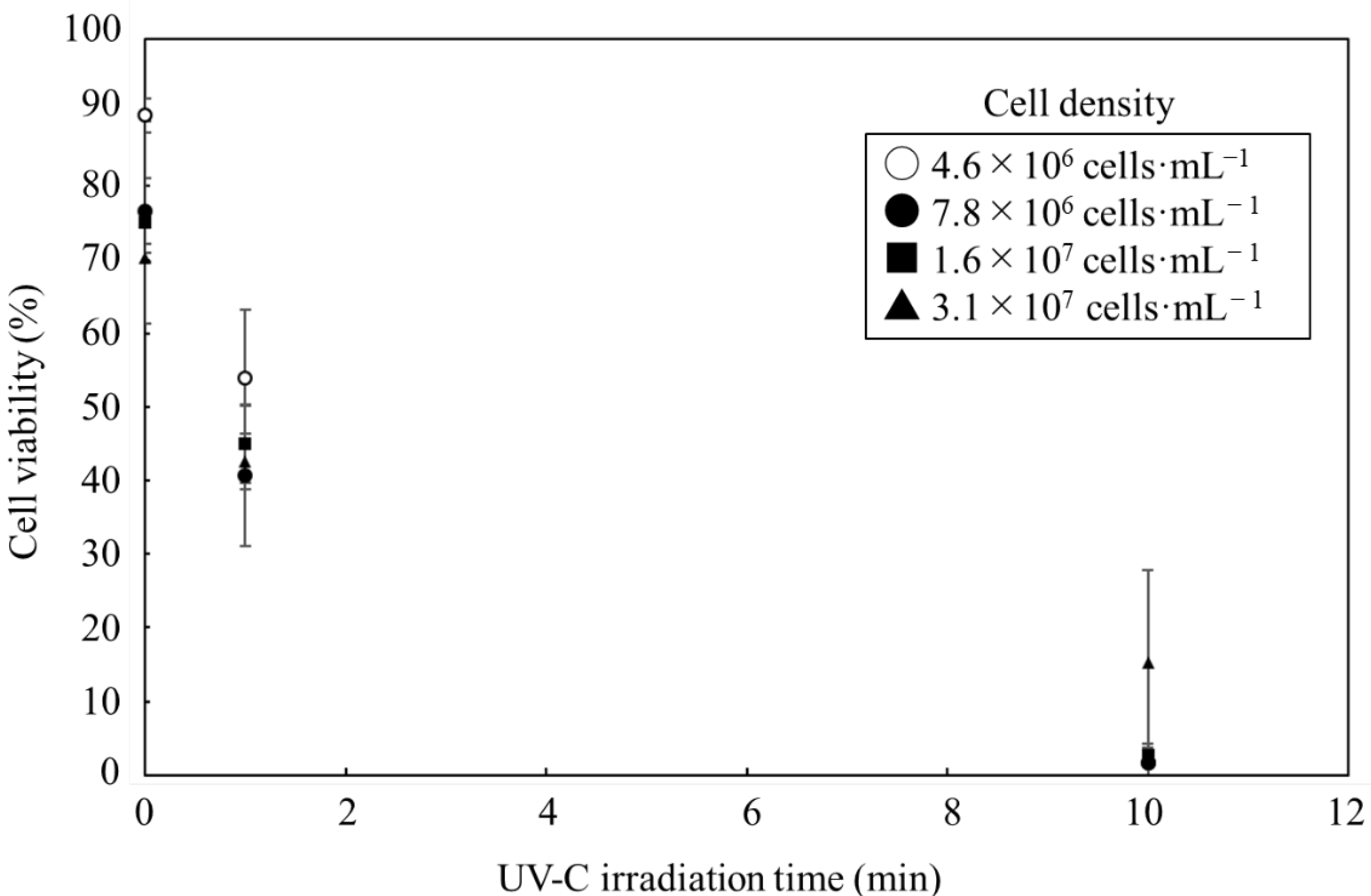
3.1. Effect of UV-C Irradiation on Cell Viability
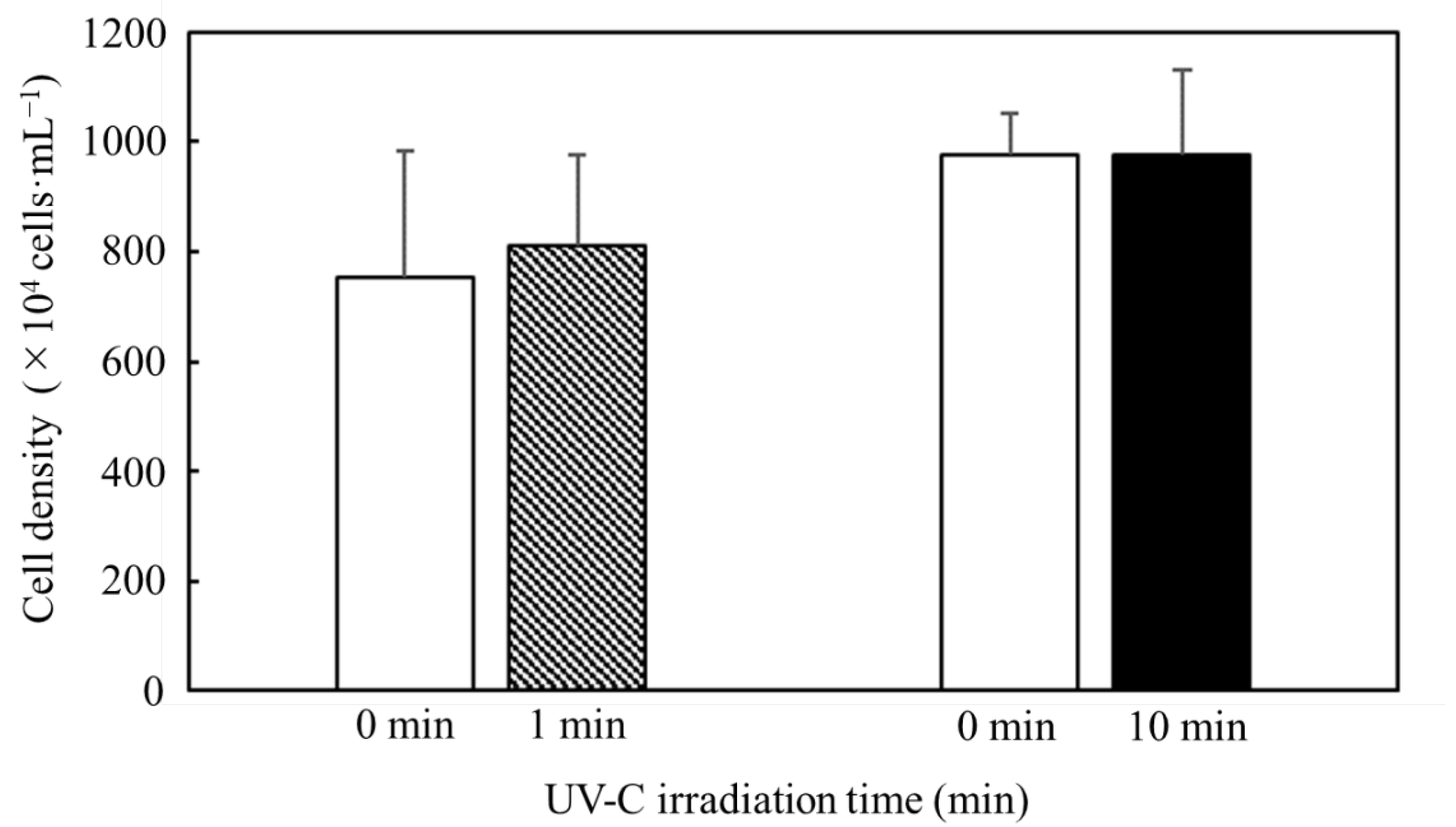
3.2. Effect of UV-C Irradiation on Cell Structure
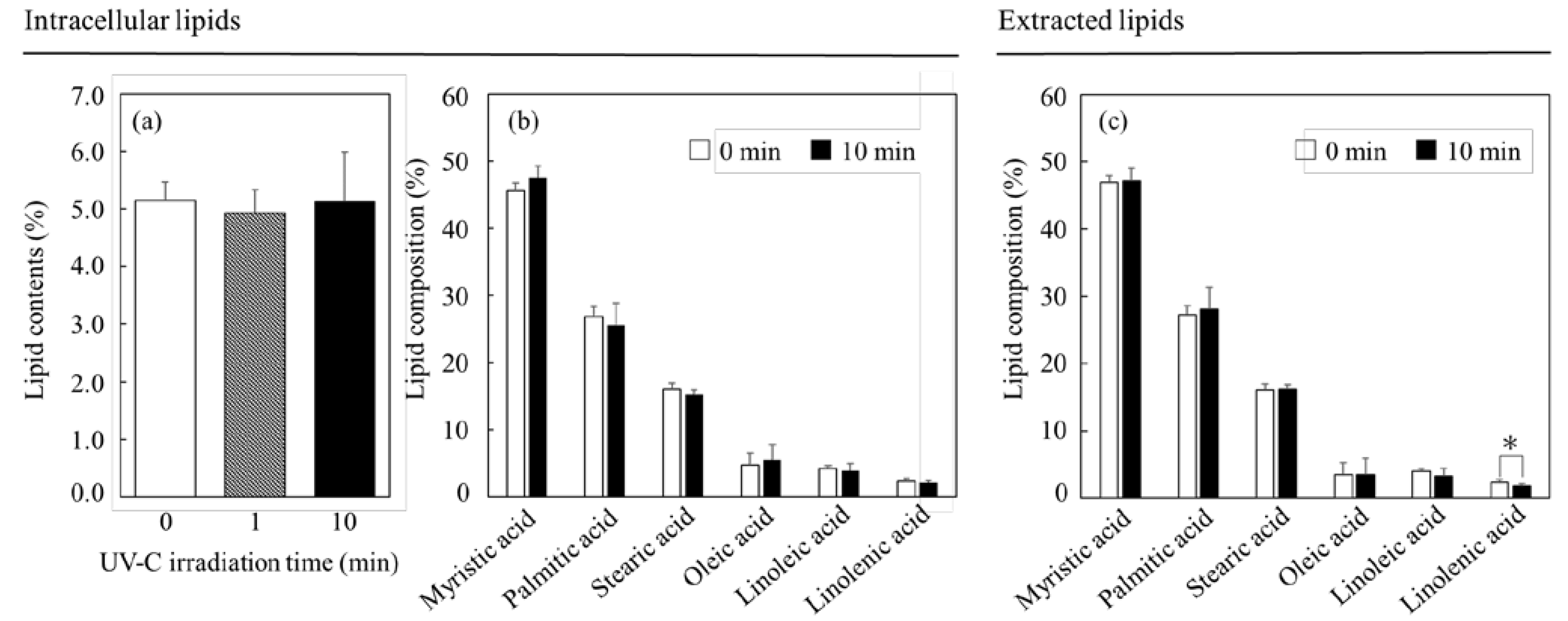
3.3. Effect of UV-C Irradiation on Lipids
3.4. Selection of a Housekeeping Gene for Quantitative PCR under UV-C Irradiation
3.5. Analysis of Transcription-Level under UV-C Irradiation
3.5.1. Effect of UV-C Irradiation on Transcription on Glycolysis
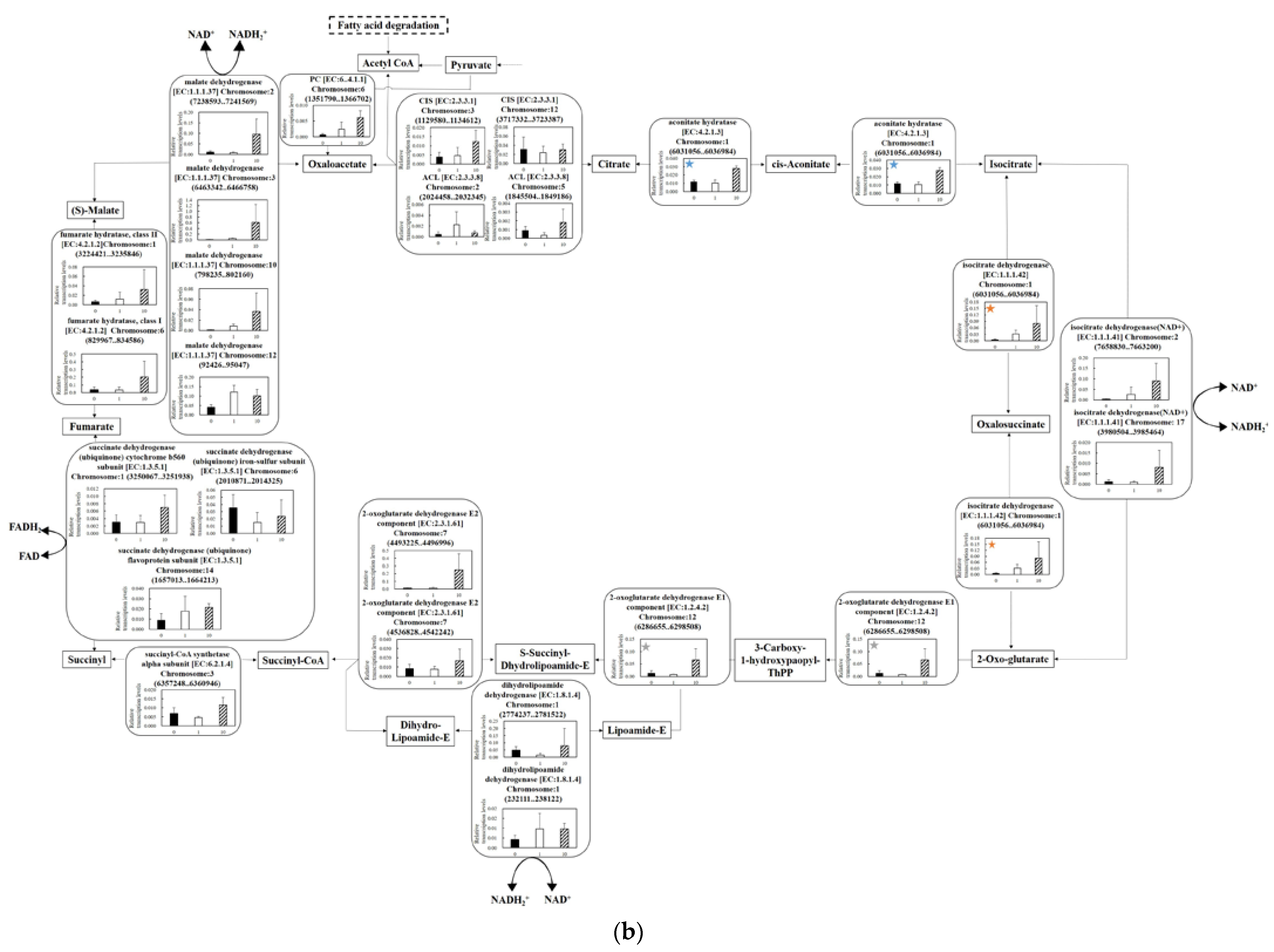
3.5.2. Effect of UV-C Irradiation on Transcription on the TCA Cycle
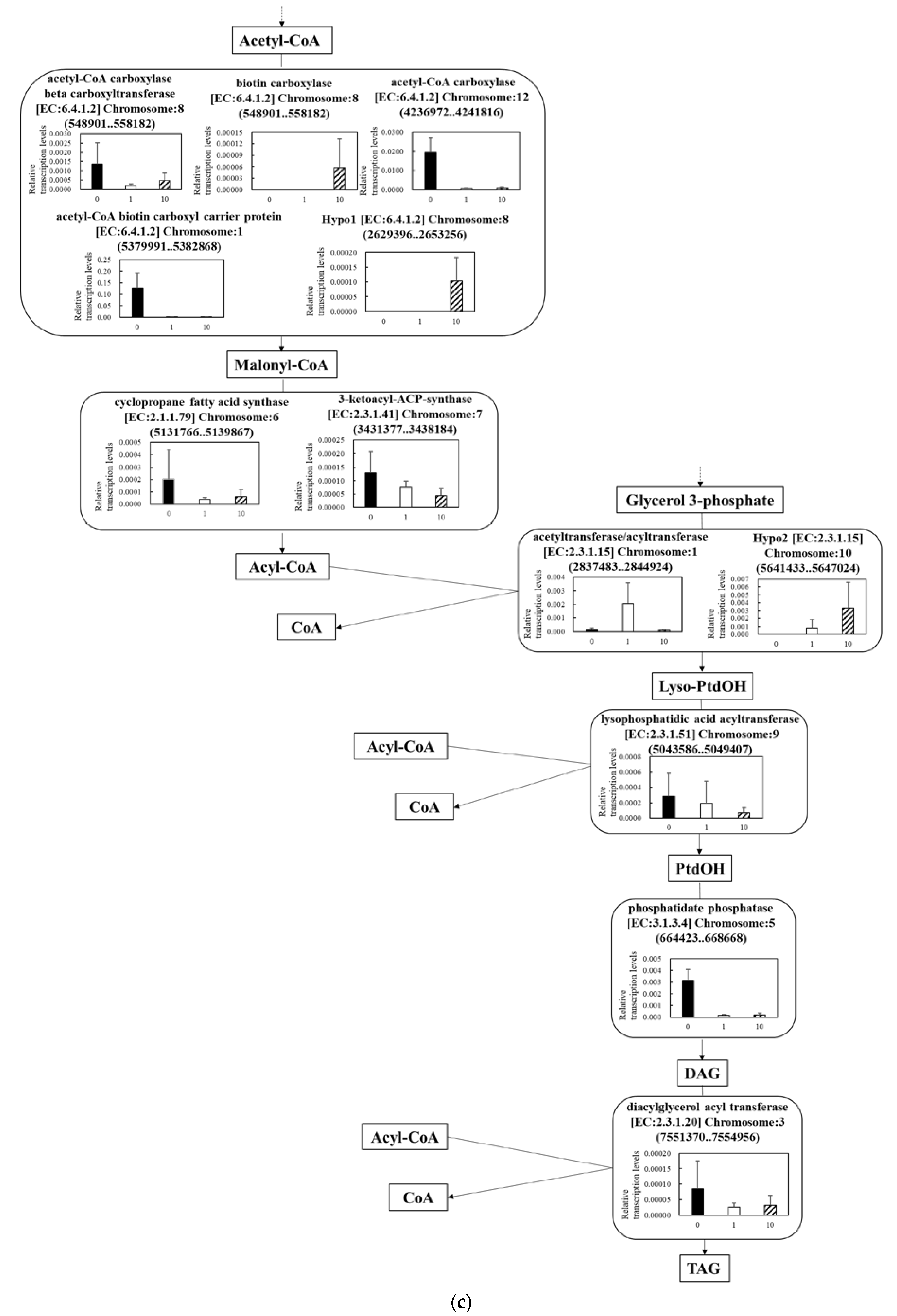
3.5.3. Effect of UV-C Irradiation on Transcription on Lipid Synthesis
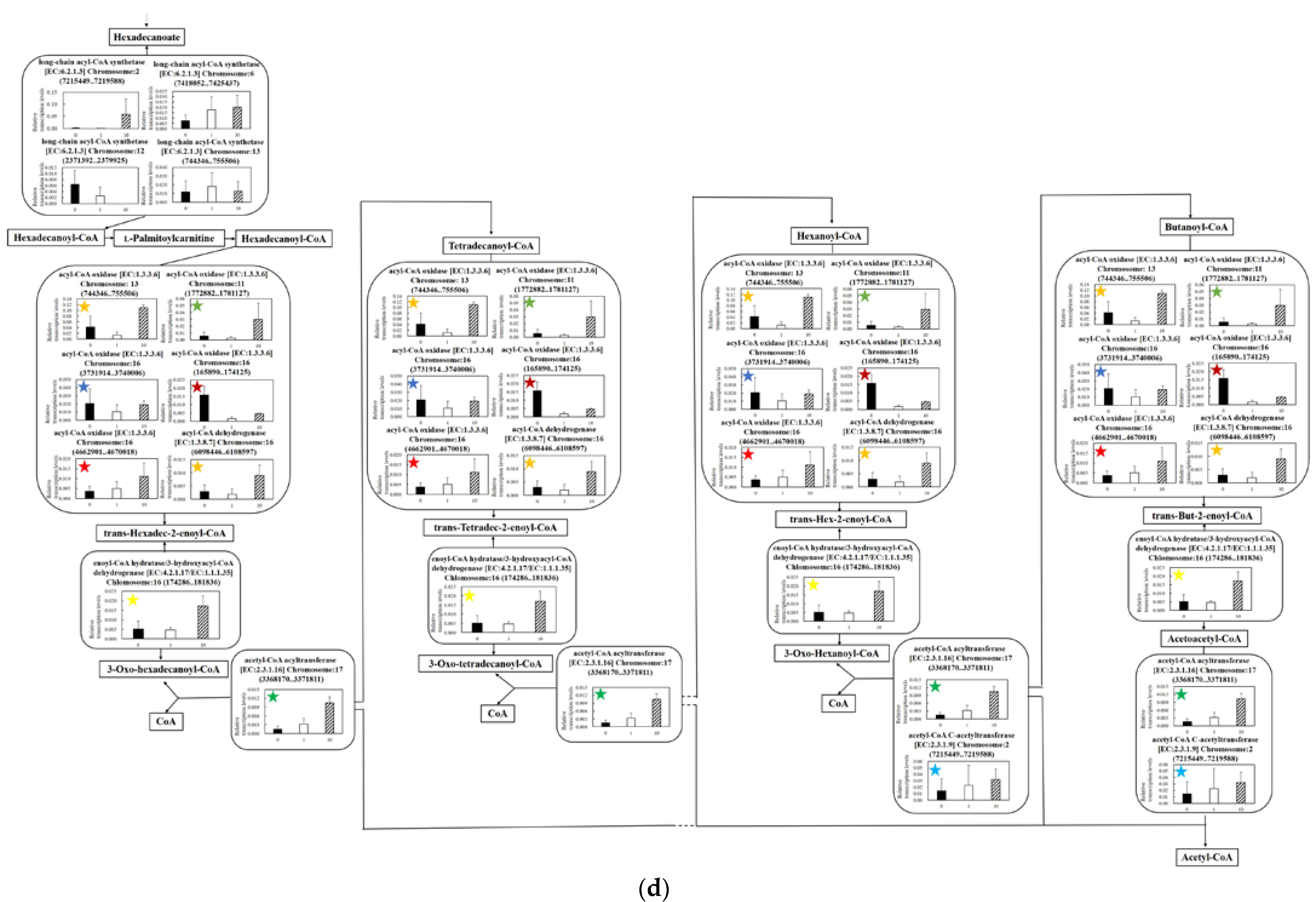
3.5.4. Effect of UV-C Irradiation on Transcription of Lipid Degradation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hasunuma, T.; Takaki, A.; Matsuda, M.; Kato, Y.; Vavricka, C.J.; Kondo, A. Single-stage astaxanthin production enhances the nonmevalonate pathway and photosynthetic central metabolism in Synechococcus sp. PCC 7002. ACS Synth. Biol. 2019, 8, 2701–2709. [Google Scholar] [CrossRef] [PubMed]
- Hitschler, L.; Nissen, S.L.; Kuntz, M.; Basen, M. Alcohol dehydrogenases adhE and adhB with broad substrate ranges are important enzymes for organic acid reduction in Thermoanaerobacter sp. strain X514. Biotechnol. Biofuels 2021, 14, 187. [Google Scholar] [CrossRef] [PubMed]
- Viktor, J.M.; Rose, H.S.; van Zyl, H.W.; Viljoen-Bloom, M. Raw starch conversion by Saccharomyces cerevisiae expressing aspergillus tubingensis amylases. Biotechnol. Biofuels 2013, 6, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Li, Y.; Wu, N.; Lan, Q.C. CO2 bio-mitigation using microalgae. Appl. Microbiol. Biotechnol. 2008, 79, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Ashour, M.; Alprol, A.E.; Alsaqufi, A.S. Aquatic plants and aquatic animals in the context of sustainability: Cultivation techniques, integration, and blue revolution. Sustainability 2022, 14, 3257. [Google Scholar] [CrossRef]
- Yoo, G.; Park, W.K.; Kim, C.W.; Choi, Y.E.; Yang, J.W. Direct lipid extraction from wet Chlamydomonas reinhardtii biomass using osmotic shock. Bioresour. Technol. 2012, 123, 717–722. [Google Scholar] [CrossRef]
- Thomas, S.; Marshall-Thompson, J.A.; Kapoore, R.V.; Vaidyanathan, S.; Pandhal, J.; Fairclough, J.P.A. The effect of high-intensity ultraviolet light to elicit microalgal cell lysis and enhance lipid extraction. Metabolites 2018, 8, 65. [Google Scholar]
- Ono, T.; Yoshioka, M. Liquid Composition Dispersed with Biomass-Derived Component, Its Production Method and Product Produced from This Liquid Composition. JP Patent 20,062,891,64A, 26 October 2006. [Google Scholar]
- Zhu, Z.; Jiang, J.; Fa, Y. Overcoming the biological contamination in microalgae and cyanobacteria mass cultivations for photosynthetic biofuel production. Molecules 2020, 25, 5220. [Google Scholar] [CrossRef]
- Schmid, J.; Hoenes, K.; Rath, M.; Vatter, P.; Hessling, M. UV-C inactivation of legionella rubrilucens. GMS Hyg. Infect. Control. 2017, 12, Doc06. [Google Scholar]
- Rout, A.K.; Singh, H.; Patel, S.; Raghvan, V.; Gautam, S.; Minda, R.; Rao, B.J.; Chary, K.V.R. Structural characterization of a novel KH-domain containing plant chloroplast endonuclease. Sci. Rep. 2018, 8, 13750. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.; Li, Y.; Schenk, M.P. UV-C mediated lipid induction and settling, a step change towards economical microalgal biodiesel production. Green Chem. 2014, 16, 3539–3548. [Google Scholar] [CrossRef]
- Ochoa-Alfaro, A.E.; Gaytán-Luna, D.E.; González-Ortega, O.; Zavala-Arias, K.G.; Paz-Maldonado, L.M.T.; Rocha-Uribe, A.; Soria-Guerra, R.E. pH effects on the lipid and fatty acids accumulation in Chlamydomonas reinhardtii. Biotechnol. Prog. 2019, 35, e2891. [Google Scholar] [CrossRef]
- Hounslow, E.; Evans, C.A.; Pandhal, J.; Sydney, T.; Couto, N.; Pham, T.K.; Gilmour, D.J.; Wright, P.C. Quantitative proteomic comparison of salt stress in Chlamydomonas reinhardtii and the snow alga Chlamydomonas nivalis reveals mechanisms for salt-triggered fatty acid accumulation via reallocation of carbon resources. Biotechnol. Biofuels 2021, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.H.; Nakanishi, A.; Ye, X.; Chang, J.S.; Chen, C.Y.; Hasunuma, T.; Kondo, A. Dynamic metabolic profiling of the marine microalga Chlamydomonas sp. JSC4 and enhancing its oil production by optimizing light intensity. Biotechnol. Biofuels 2015, 8, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moharikar, S.; D’Souza, J.S.; Rao, B.J. A homologue of the defender against the apoptotic death gene (dad1) in UV-exposed Chlamydomonas cells is downregulated with the onset of programmed cell death. J. Biosci. 2007, 32, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Moharikar, S.; D’Souza, J.S.; Kulkarni, A.B.; Rao, B.J. Apoptotic-like cell death pathway is induced in unicellular chlorophyte Chalmydomonas reinhardtii cells following UV irradiation. J. Phycology 2006, 42, 423–433. [Google Scholar] [CrossRef]
- Kayani, B.J.; Weaver, D.T.; Gopalakrishnan, V.; King, E.S.; Dolson, E.; Krishnan, N.; Pelesko, J.; Scott, M.J.; Hitomi, M.; Cadnum, J.L.; et al. UV-C tower for point-of-care decontamination of filtering facepiece respirators. Am. J. Infect. Control 2021, 49, 424–429. [Google Scholar] [CrossRef]
- She, R.C.; Chen, D.; Pak, P.; Armani, D.K.; Schubert, A.; Armani, A.M. Lightweight UV-C disinfection system. Biomed. Opt. Express 2020, 11, 4326–4332. [Google Scholar] [CrossRef]
- Colina, F.; Carbó, M.; Meijón, M.; Cañal, M.J.; Valledor, L. Low UV-C stress modulates Chlamydomonas reinhardtii biomass composition and oxidative stress response through proteomic and metabolomic changes involving novel signalers and effectors. Biotechnol. Biofuels 2020, 13, 110. [Google Scholar] [CrossRef]
- Ho, S.H.; Kondo, A.; Hasunuma, T.; Chang, J.S. Engineering strategies for improving the CO2 fixation and carbohydrate productivity of Scenedesmus obliquus CNW-N used for bioethanol fermentation. Bioresour. Technol. 2013, 143, 163–171. [Google Scholar] [CrossRef]
- Luz, D.S.; Silva, D.G.; Souza, M.M.; Giroldo, D.; Martins, C.M.G. Efficiency of neutral red, evans blue and MTT to assess viability of the freshwater microalgae Desmodesmus communis and Pediastrum boryanum. Psychol. Res. 2016, 64, 56–60. [Google Scholar]
- Masuda, A.; Murakam, K. Research on the development and the application of high-density mass production cultivation of microalgae. Ecol. Eng. 2012, 24, 57–59. [Google Scholar]
- Martysiak-Żurowska, D.; Puta, M.; Kotarska, J.; Cybula, K.; Malinowska-Pańczyk, E.; Kołodziejska, I. The effect of UV-C irradiation on lipids and selected biologically active compounds in human milk. Int. Dairy J. 2017, 66, 42–48. [Google Scholar] [CrossRef]
- Frankel, E.N. Lipid Oxidation; Woodhead Publishing: Philadelphia, PA, USA, 2012; pp. 51–64. [Google Scholar]
- Lemaire, C.; Wollman, F.A. The chloroplast ATP synthase in Chlamydomonas reinhardtii. J. Biol. Chem. 1989, 264, 10228–10234. [Google Scholar] [CrossRef]
- Wu, S.; Xu, L.; Huang, R.; Wang, Q. Improved biohydrogen production with an expression of codon-optimized hemH and lba genes in the chloroplast of Chlamydomonas reinhardtii. Bioresour. Technol. 2011, 102, 2610–2616. [Google Scholar] [CrossRef]
- Tsuchido, T.; Sakamoto, J. Stress responses in bacterial cells and development of their monitoring methods by using mutated green fluorescent proteins. Eng. Technol. 2011, 18, 75–80. [Google Scholar]
- Park, J.J.; Wang, H.; Gargouri, M.; Deshpande, R.R.; Skepper, J.N.; Holguin, F.O.; Juergens, M.T.; Shachar-Hill, Y.; Hicks, L.M.; Gang, D.R. The response of Chlamydomonas reinhardtii to nitrogen deprivation: A systems biology analysis. Plant J. 2014, 81, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Johnsona, X.; Alric, J. Central carbon metabolism and electron transport in Chlamydomonas reinhardtii: Metabolic constraints for carbon partitioning between oil and starch. Eukaryot. Cell 2013, 12, 776–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCartney, R.G.; Rice, J.E.; Sanderson, S.J.; Bunik, V.; Lindsay, H.; Lindsay, J.G. Subunit interactions in the mammalian alpha-ketoglutarate dehydrogenase complex. Evidence for direct association of the alpha-ketoglutarate dehydrogenase and dihydrolipoamide dehydrogenase components. J. Biol. Chem. 1998, 273, 24158–24164. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Reyes, I.; Diebold, L.P.; Kong, H.; Schieber, M.; Huang, H.; Hensley, C.T.; Mehta, M.M.; Wang, T.; Santos, J.H.; Woychik, R.; et al. TCA cycle and mitochondrial membrane potential are necessary for diverse biological functions. Mol. Cell 2016, 61, 199–209. [Google Scholar] [CrossRef] [Green Version]
- Soupene, E.; Kuypers, F.A. Mammalian long-chain acyl-CoA synthetases. Exp. Biol. Med. 2008, 233, 507–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.J.; Le Dall, M.T.; Wach, Y.; Laroche, C.; Belin, J.M.; Gaillardin, C.; Nicaud, J.M. Evaluation of acyl coenzyme A oxidase (Aox) isozyme function in the n-alkane-assimilating yeast yarrowia lipolytica. J. Bacteriol. 1999, 181, 5140–5148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative stress is a mediator for increased lipid accumulation in a newly isolated Dunaliella salina strain. PLoS ONE 2014, 9, e91957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghuge, S.A.; Kadam, U.S.; Hong, J.C. Selenoprotein: Potential player in redox regulation in Chlamydomonas reinhardtii. Antioxidants 2022, 11, 1630. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakanishi, A.; Ozawa, N.; Watanabe, M. Evaluation of Shifts of Gene Transcription Levels of Unicellular Green Alga Chlamydomonas reinhardtii Due to UV-C Irradiation. Microorganisms 2023, 11, 633. https://doi.org/10.3390/microorganisms11030633
Nakanishi A, Ozawa N, Watanabe M. Evaluation of Shifts of Gene Transcription Levels of Unicellular Green Alga Chlamydomonas reinhardtii Due to UV-C Irradiation. Microorganisms. 2023; 11(3):633. https://doi.org/10.3390/microorganisms11030633
Chicago/Turabian StyleNakanishi, Akihito, Nanami Ozawa, and Masahiko Watanabe. 2023. "Evaluation of Shifts of Gene Transcription Levels of Unicellular Green Alga Chlamydomonas reinhardtii Due to UV-C Irradiation" Microorganisms 11, no. 3: 633. https://doi.org/10.3390/microorganisms11030633