
The Tubulin Superfamily in Apicomplexan Parasites

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Identification of Apicomplexan Tubulin Genes
2.2. Tubulin Protein Analysis
2.3. Tubulin Transcriptomics and Proteomics
3. Results
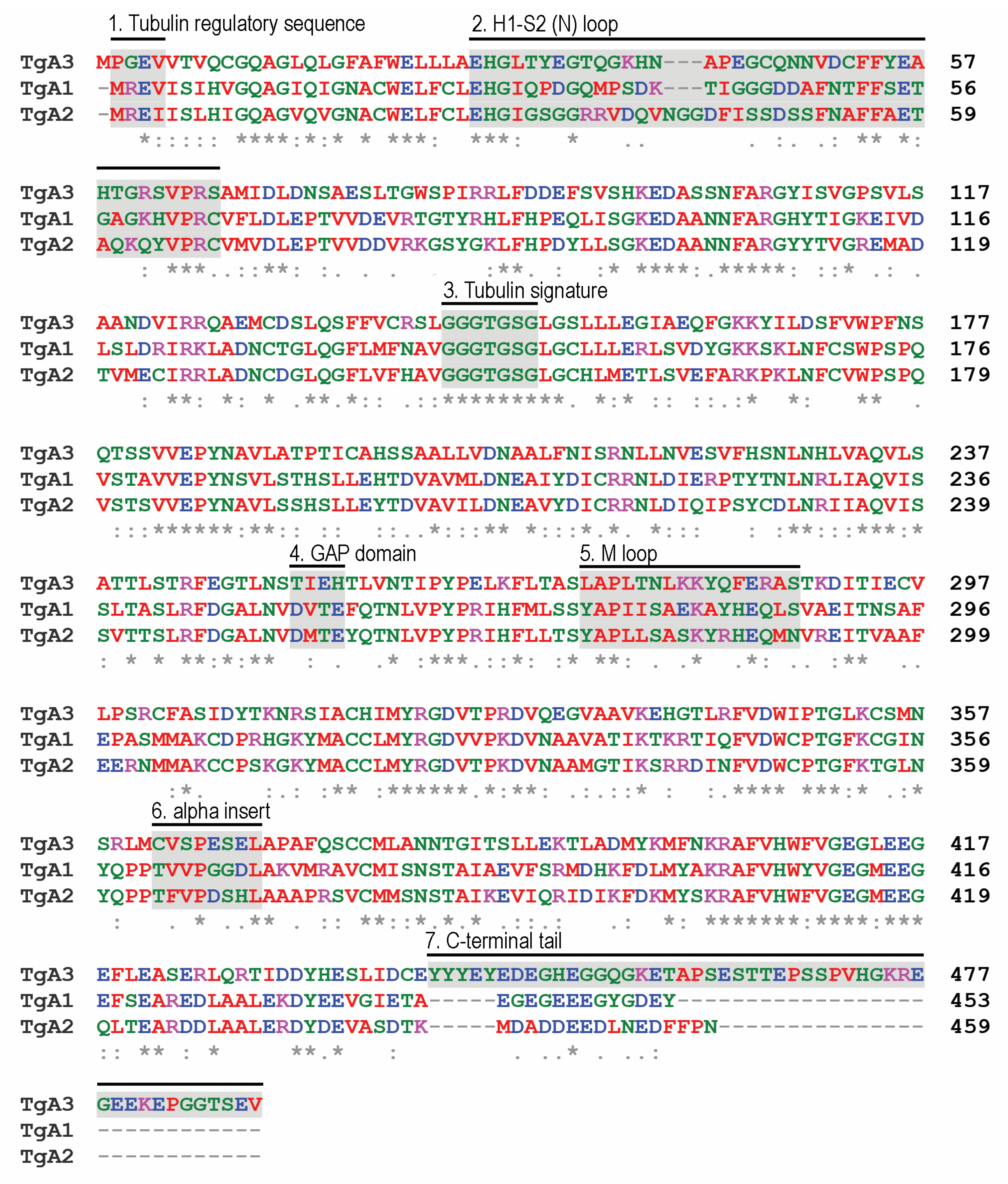
3.1. Apicomplexans Display Simple α-, β-, and γ-Isotype Repertoires

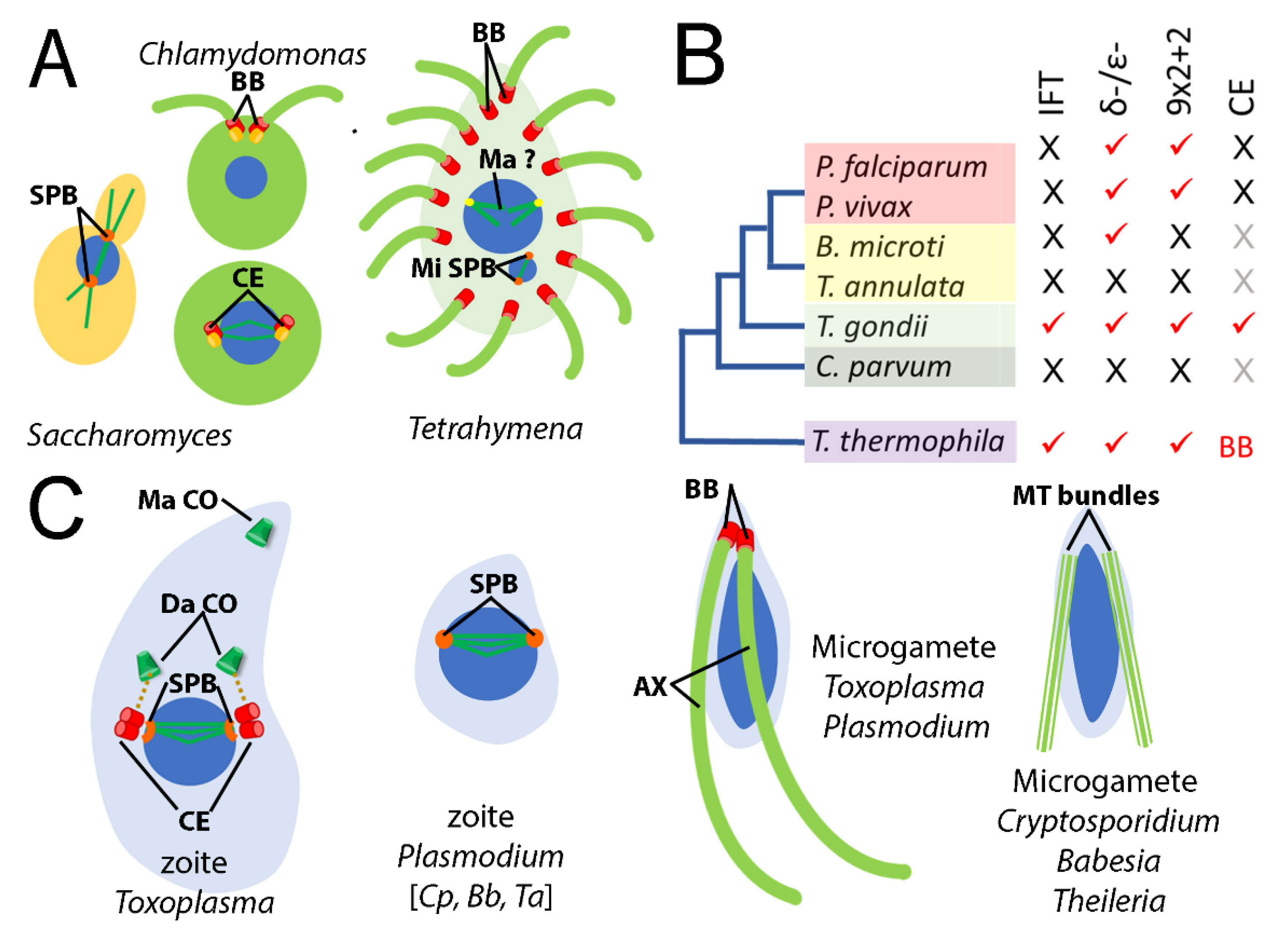
3.2. Not All Apicomplexans Have Genes for δ- and ε-Tubulins
3.3. Tubulin Expression Is Developmentally Regulated during the Parasite Life Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gudimchuk, N.B.; McIntosh, J.R. Regulation of microtubule dynamics, mechanics and function through the growing tip. Nat. Rev. Mol. Cell Biol. 2021, 22, 777–795. [Google Scholar] [CrossRef]
- Kristensson, M.A. The Game of Tubulins. Cells 2021, 10, 745. [Google Scholar] [CrossRef]
- Lopez-Fanarraga, M.; Avila, J.; Guasch, A.; Coll, M.; Zabala, J.C. Review: Postchaperonin tubulin folding cofactors and their role in microtubule dynamics. J. Struct. Biol. 2001, 135, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; Tranter, D.; Pardon, E.; Chi, G.; Kramer, H.; Happonen, L.; Knee, K.M.; Janz, J.M.; Steyaert, J.; Bulawa, C.; et al. Snapshots of actin and tubulin folding inside the TRiC chaperonin. Nat. Struct. Mol. Biol. 2022, 29, 420–429. [Google Scholar] [CrossRef]
- Findeisen, P.; Muhlhausen, S.; Dempewolf, S.; Hertzog, J.; Zietlow, A.; Carlomagno, T.; Kollmar, M. Six subgroups and extensive recent duplications characterize the evolution of the eukaryotic tubulin protein family. Genome Biol. Evol. 2014, 6, 2274–2288. [Google Scholar] [CrossRef] [Green Version]
- McKean, P.G.; Vaughan, S.; Gull, K. The extended tubulin superfamily. J. Cell Sci. 2001, 114, 2723–2733. [Google Scholar] [CrossRef]
- Dutcher, S.K. Long-lost relatives reappear: Identification of new members of the tubulin superfamily. Curr. Opin. Microbiol. 2003, 6, 634–640. [Google Scholar] [CrossRef]
- Marshall, W.F.; Rosenbaum, J.L. Tubulin superfamily: Giving birth to triplets. Curr. Biol. 2003, 13, R55–R56. [Google Scholar] [CrossRef] [Green Version]
- Zupa, E.; Liu, P.; Wurtz, M.; Schiebel, E.; Pfeffer, S. The structure of the γ-TuRC: A 25-years-old molecular puzzle. Curr. Opin. Struct. Biol. 2021, 66, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Kollman, J.M.; Merdes, A.; Mourey, L.; Agard, D.A. Microtubule nucleation by γ-tubulin complexes. Nat. Rev. Mol. Cell Biol. 2011, 12, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Fulton, C. The Amazing Evolutionary Complexity of Eukaryotic Tubulins: Lessons from Naegleria and the Multi-tubulin Hypothesis. Front. Cell Dev. Biol. 2022, 10, 867374. [Google Scholar] [CrossRef] [PubMed]
- Nsamba, E.T.; Gupta, M.L., Jr. Tubulin isotypes—Functional insights from model organisms. J. Cell Sci. 2022, 135, jcs259539. [Google Scholar] [CrossRef] [PubMed]
- Bera, A.; Gupta, M.L., Jr. Microtubules in Microorganisms: How Tubulin Isotypes Contribute to Diverse Cytoskeletal Functions. Front. Cell Dev. Biol. 2022, 10, 913809. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.G.; Borisy, G.G. Evolution of the multi-tubulin hypothesis. Bioessays 1997, 19, 451–454. [Google Scholar] [CrossRef] [PubMed]
- Ti, S.C.; Alushin, G.M.; Kapoor, T.M. Human β-Tubulin Isotypes Can Regulate Microtubule Protofilament Number and Stability. Dev. Cell 2018, 47, 175–190.e5. [Google Scholar] [CrossRef] [Green Version]
- Pamula, M.C.; Ti, S.-C.; Kapoor, T.M. The structured core of human β tubulin confers isotype-specific polymerization properties. J. Cell Biol. 2016, 213, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Hausrat, T.J.; Radwitz, J.; Lombino, F.L.; Breiden, P.; Kneussel, M. α- and β-tubulin isotypes are differentially expressed during brain development. Dev. Neurobiol. 2021, 81, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Turk, E.; Wills, A.A.; Kwon, T.; Sedzinski, J.; Wallingford, J.B.; Stearns, T. ζ-Tubulin Is a Member of a Conserved Tubulin Module and Is a Component of the Centriolar Basal Foot in Multiciliated Cells. Curr. Biol. 2015, 25, 2177–2183. [Google Scholar] [CrossRef] [Green Version]
- Stathatos, G.G.; Dunleavy, J.E.M.; Zenker, J.; O’Bryan, M.K. δ and ε tubulin in mammalian development. Trends Cell Biol. 2021, 31, 774–787. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Luo, Y.; Zhou, S.; An, L.; Wang, C.; Jin, Q.; Zhou, M.; Xu, J.R. Molecular evolution and functional divergence of tubulin superfamily in the fungal tree of life. Sci. Rep. 2014, 4, 6746. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.C.; Bettencourt-Dias, M.; Durand, B.; Megraw, T.L. Drosophila melanogaster as a model for basal body research. Cilia 2016, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodges, M.E.; Scheumann, N.; Wickstead, B.; Langdale, J.A.; Gull, K. Reconstructing the evolutionary history of the centriole from protein components. J. Cell Sci. 2010, 123, 1407–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, A.; Sankaralingam, P.; O’Connell, K.F.; Muller-Reichert, T. Revisiting Centrioles in Nematodes-Historic Findings and Current Topics. Cells 2018, 7, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Lancet Editors. Malaria in 2022: A year of opportunity. Lancet 2022, 399, 1573. [Google Scholar] [CrossRef]
- Vannier, E.; Gewurz, B.E.; Krause, P.J. Human babesiosis. Infect. Dis. Clin. North Am. 2008, 22, 469–488. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Christie, J.; Koster, L.; Du, A.; Yao, C. Emerging Human Babesiosis with “Ground Zero” in North America. Microorganisms 2021, 9, 440. [Google Scholar] [CrossRef]
- Ullah, R.; Shams, S.; Khan, M.A.; Ayaz, S.; ul Akbar, N.; Din, Q.u.; Khan, A.; Leon, R.; Zeb, J. Epidemiology and molecular characterization of Theileria annulata in cattle from central Khyber Pakhtunkhwa, Pakistan. PLoS ONE 2021, 16, e0249417. [Google Scholar] [CrossRef]
- Roy, S.; Bhandari, V.; Barman, M.; Kumar, P.; Bhanot, V.; Arora, J.S.; Singh, S.; Sharma, P. Population Genetic Analysis of the Theileria annulata Parasites Identified Limited Diversity and Multiplicity of Infection in the Vaccine from India. Front. Microbiol. 2020, 11, 579929. [Google Scholar] [CrossRef]
- Keeling, P.J. Reduction and compaction in the genome of the apicomplexan parasite Cryptosporidium parvum. Dev. Cell 2004, 6, 614–616. [Google Scholar] [CrossRef] [Green Version]
- Gerace, E.; Lo Presti, V.D.M.; Biondo, C. Cryptosporidium Infection: Epidemiology, Pathogenesis and Differential Diagnosis. Eur. J. Microbiol. Immunol. 2019, 9, 119–123. [Google Scholar] [CrossRef]
- Barta, J.R.; Schrenzel, M.D.; Carreno, R.; Rideout, B.A. The genus Atoxoplasma (Garnham 1950) as a junior objective synonym of the genus Isospora (Schneider 1881) species infecting birds and resurrection of Cystoisospora (Frenkel 1977) as the correct genus for Isospora species infecting mammals. J. Parasitol. 2005, 91, 726–727. [Google Scholar] [CrossRef]
- Dubey, J.P.; Almeria, S. Cystoisospora belli infections in humans: The past 100 years. Parasitology 2019, 146, 1490–1527. [Google Scholar] [CrossRef]
- Li, J.; Cui, Z.; Qi, M.; Zhang, L. Advances in Cyclosporiasis Diagnosis and Therapeutic Intervention. Front. Cell. Infect. Microbiol. 2020, 10, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Osorio, S.; Chaparro-Gutierrez, J.J.; Gomez-Osorio, L.M. Overview of Poultry Eimeria Life Cycle and Host-Parasite Interactions. Front. Vet. Sci. 2020, 7, 384. [Google Scholar] [CrossRef]
- Blake, D.P.; Worthing, K.; Jenkins, M.C. Exploring Eimeria Genomes to Understand Population Biology: Recent Progress and Future Opportunities. Genes 2020, 11, 1103. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Exposito, D.; Ferre, I.; Ortega-Mora, L.M.; Alvarez-Garcia, G. Advances in the diagnosis of bovine besnoitiosis: Current options and applications for control. Int. J. Parasitol. 2017, 47, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.L.; Kruszon-Moran, D.; Elder, S.; Rivera, H.N.; Press, C.; Montoya, J.G.; McQuillan, G.M. Toxoplasma gondii Infection in the United States, 2011–2014. Am. J. Trop. Med. Hyg. 2018, 98, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Tran, J.Q.; Li, C.; Chyan, A.; Chung, L.; Morrissette, N.S. SPM1 stabilizes subpellicular microtubules in Toxoplasma gondii. Eukaryot. Cell 2012, 11, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; He, Y.; Benmerzouga, I.; Sullivan, W.J., Jr.; Morrissette, N.S.; Murray, J.M.; Hu, K. An ensemble of specifically targeted proteins stabilizes cortical microtubules in the human parasite Toxoplasma gondii. Mol. Biol. Cell 2016, 27, 549–571. [Google Scholar] [CrossRef] [Green Version]
- De Leon, J.C.; Scheumann, N.; Beatty, W.; Beck, J.R.; Tran, J.Q.; Yau, C.; Bradley, P.J.; Gull, K.; Wickstead, B.; Morrissette, N.S. A SAS-6-like protein suggests that the Toxoplasma conoid complex evolved from flagellar components. Eukaryot. Cell 2013, 12, 1009–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévêque, M.F.; Berry, L.; Besteiro, S. An evolutionarily conserved SSNA1/DIP13 homologue is a component of both basal and apical complexes of Toxoplasma gondii. Sci. Rep. 2016, 6, 27809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenal, K.; Dubremetz, J.F.; Lebrun, M.; Soldati-Favre, D. Gliding motility powers invasion and egress in Apicomplexa. Nat. Rev. Microbiol. 2017, 15, 645–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wall, R.J.; Roques, M.; Katris, N.J.; Koreny, L.; Stanway, R.R.; Brady, D.; Waller, R.F.; Tewari, R. SAS6-like protein in Plasmodium indicates that conoid-associated apical complex proteins persist in invasive stages within the mosquito vector. Sci. Rep. 2016, 6, 28604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosetti, N.; Dos Santos Pacheco, N.; Soldati-Favre, D.; Jacot, D. Three F-actin assembly centers regulate organelle inheritance, cell-cell communication and motility in Toxoplasma gondii. eLife 2019, 8, e42669. [Google Scholar] [CrossRef]
- Cruz-Bustos, T.; Feix, A.S.; Ruttkowski, B.; Joachim, A. Sexual Development in Non-Human Parasitic Apicomplexa: Just Biology or Targets for Control? Animals 2021, 11, 2891. [Google Scholar] [CrossRef]
- Bechtsi, D.P.; Waters, A.P. Genomics and epigenetics of sexual commitment in Plasmodium. Int. J. Parasitol. 2017, 47, 425–434. [Google Scholar] [CrossRef]
- Sinden, R.E. Targeting the Parasite to Suppress Malaria Transmission. Adv. Parasitol. 2017, 97, 147–185. [Google Scholar]
- Marques, S.R.; Ramakrishnan, C.; Carzaniga, R.; Blagborough, A.M.; Delves, M.J.; Talman, A.M.; Sinden, R.E. An essential role of the basal body protein SAS-6 in Plasmodium male gamete development and malaria transmission. Cell. Microbiol. 2015, 17, 191–206. [Google Scholar] [CrossRef] [Green Version]
- Morrissette, N. Targeting Toxoplasma tubules: Tubulin, microtubules and associated proteins in a human pathogen. Eukaryot. Cell 2015, 14, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Bell, A. Microtubule inhibitors as potential antimalarial agents. Parasitol. Today 1998, 14, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The eukaryotic pathogen, vector and host bioinformatics resource center. Nucleic Acids Res. 2021, 50, D898–D911. [Google Scholar] [CrossRef] [PubMed]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukes, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coordinators, N.R. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal omega. Curr. Protoc. Bioinform. 2014, 48, 3–13. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Lasonder, E.; Rijpma, S.R.; van Schaijk, B.C.; Hoeijmakers, W.A.; Kensche, P.R.; Gresnigt, M.S.; Italiaander, A.; Vos, M.W.; Woestenenk, R.; Bousema, T.; et al. Integrated transcriptomic and proteomic analyses of P. falciparum gametocytes: Molecular insight into sex-specific processes and translational repression. Nucleic Acids Res. 2016, 44, 6087–6101. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Diaz, E.; Yerbanga, R.S.; Lefevre, T.; Cohuet, A.; Rowley, M.J.; Ouedraogo, J.B.; Corces, V.G. Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in Anopheles gambiae. Sci. Rep. 2017, 7, 40655. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Barragan, M.J.; Lemieux, J.; Quinones, M.; Williamson, K.C.; Molina-Cruz, A.; Cui, K.; Barillas-Mury, C.; Zhao, K.; Su, X.Z. Directional gene expression and antisense transcripts in sexual and asexual stages of Plasmodium falciparum. BMC Genom. 2011, 12, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanghi, G.; Vembar, S.S.; Baumgarten, S.; Ding, S.; Guizetti, J.; Bryant, J.M.; Mattei, D.; Jensen, A.T.R.; Renia, L.; Goh, Y.S.; et al. A Specific PfEMP1 Is Expressed in P. falciparum Sporozoites and Plays a Role in Hepatocyte Infection. Cell Rep. 2018, 22, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, C.; Maier, S.; Walker, R.A.; Rehrauer, H.; Joekel, D.E.; Winiger, R.R.; Basso, W.U.; Grigg, M.E.; Hehl, A.B.; Deplazes, P.; et al. An experimental genetically attenuated live vaccine to prevent transmission of Toxoplasma gondii by cats. Sci. Rep. 2019, 9, 1474. [Google Scholar] [CrossRef]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef] [Green Version]
- Kemphues, K.J.; Raff, R.A.; Kaufman, T.C.; Raff, E.C. Mutation in a structural gene for a β-tubulin specific to testis in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1979, 76, 3991–3995. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, D.J.; Fujioka, H.; Fried, M.; Keister, D.B.; Aikawa, M.; Kaslow, D.C. α-tubulin II is a male-specific protein in Plasmodium falciparum. Mol. Biochem. Parasitol. 1992, 56, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Fennell, B.J.; Al-shatr, Z.A.; Bell, A. Isotype expression, post-translational modification and stage-dependent production of tubulins in erythrocytic Plasmodium falciparum. Int. J. Parasitol. 2008, 38, 527–539. [Google Scholar] [CrossRef]
- Schwank, S.; Sutherland, C.J.; Drakeley, C.J. Promiscuous expression of α-tubulin II in maturing male and female Plasmodium falciparum gametocytes. PLoS ONE 2010, 5, e14470. [Google Scholar] [CrossRef] [Green Version]
- Kooij, T.W.; Franke-Fayard, B.; Renz, J.; Kroeze, H.; van Dooren, M.W.; Ramesar, J.; Augustijn, K.D.; Janse, C.J.; Waters, A.P. Plasmodium berghei α-tubulin II: A role in both male gamete formation and asexual blood stages. Mol. Biochem. Parasitol. 2005, 144, 16–26. [Google Scholar] [CrossRef]
- Lin, Z.; Gasic, I.; Chandrasekaran, V.; Peters, N.; Shao, S.; Mitchison, T.J.; Hegde, R.S. TTC5 mediates autoregulation of tubulin via mRNA degradation. Science 2020, 367, 100–104. [Google Scholar] [CrossRef]
- Anders, K.R.; Botstein, D. Dominant-lethal α-tubulin mutants defective in microtubule depolymerization in yeast. Mol. Biol. Cell 2001, 12, 3973–3986. [Google Scholar] [CrossRef] [PubMed]
- Salomaki, E.D.; Terpis, K.X.; Rueckert, S.; Kotyk, M.; Varadinova, Z.K.; Cepicka, I.; Lane, C.E.; Kolisko, M. Gregarine single-cell transcriptomics reveals differential mitochondrial remodeling and adaptation in apicomplexans. BMC Biol. 2021, 19, 77. [Google Scholar] [CrossRef]
- Janouškovec, J.; Paskerova, G.G.; Miroliubova, T.S.; Mikhailov, K.V.; Birley, T.; Aleoshin, V.V.; Simdyanov, T.G. Apicomplexan-like parasites are polyphyletic and widely but selectively dependent on cryptic plastid organelles. eLife 2019, 8, e49662. [Google Scholar] [CrossRef] [PubMed]
- Templeton, T.J.; Pain, A. Diversity of extracellular proteins during the transition from the ‘proto-apicomplexan’ alveolates to the apicomplexan obligate parasites. Parasitology 2016, 143, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.H.; Wares, J.P.; Kissinger, J.C. The Apicomplexan whole-genome phylogeny: An analysis of incongruence among gene trees. Mol. Biol. Evol. 2008, 25, 2689–2698. [Google Scholar] [CrossRef] [PubMed]
- Aylett, C.H.S.; Duggin, I.G. The Tubulin Superfamily in Archaea. Sub-Cell. Biochem. 2017, 84, 393–417. [Google Scholar]
- Hufnagel, L.A. The Cilioprotist Cytoskeleton, a Model for Understanding How Cell Architecture and Pattern Are Specified: Recent Discoveries from Ciliates and Comparable Model Systems. Methods Mol. Biol. 2022, 2364, 251–295. [Google Scholar]
- Bayless, B.A.; Galati, D.F.; Pearson, C.G. Tetrahymena basal bodies. Cilia 2016, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Morrissette, N.S.; Sibley, L.D. Disruption of microtubules uncouples budding and nuclear division in Toxoplasma gondii. J. Cell Sci. 2002, 115, 1017–1025. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Kingdom Chromista and its eight phyla: A new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences. Protoplasma 2018, 255, 297–357. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Rzhetsky, A. Evolutionary relationships of eukaryotic kingdoms. J. Mol. Evol. 1996, 42, 183–193. [Google Scholar] [CrossRef]
- Gajadhar, A.A.; Marquardt, W.C.; Hall, R.; Gunderson, J.; Ariztia-Carmona, E.V.; Sogin, M.L. Ribosomal RNA sequences of Sarcocystis muris, Theileria annulata and Crypthecodinium cohnii reveal evolutionary relationships among apicomplexans, dinoflagellates and ciliates. Mol. Biochem. Parasitol. 1991, 45, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T. Axoneme Structure from Motile Cilia. Cold Spring Harb. Perspect. Biol. 2017, 9, a028076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, M.; Van Houten, J. Using Paramecium as a Model for Ciliopathies. Genes 2021, 12, 1493. [Google Scholar] [CrossRef]
- Ryu, H.; Lee, H.; Lee, J.; Noh, H.; Shin, M.; Kumar, V.; Hong, S.; Kim, J.; Park, S. The molecular dynamics of subdistal appendages in multi-ciliated cells. Nat. Commun. 2021, 12, 612. [Google Scholar] [CrossRef] [PubMed]
- Chong, W.M.; Wang, W.-J.; Lo, C.-H.; Chiu, T.-Y.; Chang, T.-J.; Liu, Y.-P.; Tanos, B.; Mazo, G.; Tsou, M.-F.B.; Jane, W.-N.; et al. Super-resolution microscopy reveals coupling between mammalian centriole subdistal appendages and distal appendages. eLife 2020, 9, e53580. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, L.B.; Schroder, J.M.; Satir, P.; Christensen, S.T. The ciliary cytoskeleton. Compr. Physiol. 2012, 2, 779–803. [Google Scholar]
- Nechipurenko, I.; Sengupta, P. The rise and fall of basal bodies in the nematode Caenorhabditis elegans. Cilia 2017, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Smith, H. Nematode Sperm Motility. In WormBook; Chalfie, M., Ed.; 2006; Available online: http://www.wormbook.org (accessed on 1 January 2023).
- Medina, E.M.; Buchler, N.E. Chytrid fungi. Curr. Biol. 2020, 30, R516–R520. [Google Scholar] [CrossRef]
- Leander, B.S.; Keeling, P.J. Morphostasis in alveolate evolution. Trends Ecol. Evol. 2003, 18, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Francia, M.E.; Dubremetz, J.F.; Morrissette, N.S. Basal body structure and composition in the apicomplexans Toxoplasma and Plasmodium. Cilia 2015, 5, 3. [Google Scholar] [CrossRef] [Green Version]
- Zeeshan, M.; Brady, D.; Markus, R.; Vaughan, S.; Ferguson, D.; Holder, A.A.; Tewari, R. Plasmodium SAS4: Basal body component of male cell which is dispensable for parasite transmission. Life Sci. Alliance 2022, 5. [Google Scholar] [CrossRef]
- Fritz-Laylin, L.K.; Fulton, C. Naegleria: A classic model for de novo basal body assembly. Cilia 2016, 5, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizukami, I.; Gall, J. Centriole replication. II. Sperm formation in the fern, Marsilea and the cycad, Zamia. J. Cell Biol. 1966, 29, 97–111. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.A.; Ferguson, D.J.; Miller, C.M.; Smith, N.C. Sex and Eimeria: A molecular perspective. Parasitology 2013, 140, 1701–1717. [Google Scholar] [CrossRef] [PubMed]
- Sinden, R.E.; Talman, A.; Marques, S.R.; Wass, M.N.; Sternberg, M.J. The flagellum in malarial parasites. Curr. Opin. Microbiol. 2010, 13, 491–500. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Heydorn, A.O. Electron microscopical study on gamogony of Sarcocystis suihominis in human tissue cultures. Z Parasitenkd 1979, 58, 97–113. [Google Scholar] [CrossRef]
- Ferguson, D.J. Toxoplasma gondii and sex: Essential or optional extra? Trends Parasitol. 2002, 18, 355–359. [Google Scholar] [CrossRef]
- Fernando, M.A. Fine structural changes associated with microgametogenesis of Eimeria acervulina in chickens. Z. Parasitenkd. 1973, 43, 33–42. [Google Scholar] [CrossRef]
- Dutcher, S.K.; Trabuco, E.C. The UNI3 gene is required for assembly of basal bodies of Chlamydomonas and encodes δ-tubulin, a new member of the tubulin superfamily. Mol. Biol. Cell 1998, 9, 1293–1308. [Google Scholar] [CrossRef] [PubMed]
- Dutcher, S.K.; Morrissette, N.S.; Preble, A.M.; Rackley, C.; Stanga, J. ε-tubulin is an essential component of the centriole. Mol. Biol. Cell 2002, 13, 3859–3869. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, J.L.; Lechtreck, K.F. Chlamydomonas Basal Bodies as Flagella Organizing Centers. Cells 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Garnham, P.C.; Bird, R.G.; Baker, J.R. Electron microscope studies of motile stages of malaria parasites. V. Exflagellation in Plasmodium, Hepatocystis and Leucocytozoon. Trans. R. Soc. Trop. Med. Hyg. 1967, 61, 58–68. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Weber, G.; Schein, E.; Büscher, G. Elektronenmikroskopische Untersuchung an Entwicklungsstadien von Theileria annulata (Dschunkowsky, Luhs, 1904) im Darm und in der Hämolymphe von Hyalomma anatolicum excavatum (Koch, 1844). Z. Parasitenkd. 1975, 48, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Rudzinska, M.A.; Spielman, A.; Lewengrub, S.; Trager, W.; Piesman, J. Sexuality in piroplasms as revealed by electron microscopy in Babesia microti. Proc. Natl. Acad. Sci. USA 1983, 80, 2966–2970. [Google Scholar] [CrossRef] [Green Version]
- Rudzinska, M.A.; Spielman, A.; Lewengrub, S.; Piesman, J.; Karakashian, S. Penetration of the peritrophic membrane of the tick by Babesia microti. Cell Tissue Res. 1982, 221, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Weber, G.; Friedhoff, K.T. Preliminary observations on the ultrastructure of supposed sexual stages of Babesia bigemina (Piroplasmea). Z. Parasitenkd. 1977, 53, 83–92. [Google Scholar] [CrossRef]
- Tandel, J.; English, E.D.; Sateriale, A.; Gullicksrud, J.A.; Beiting, D.P.; Sullivan, M.C.; Pinkston, B.; Striepen, B. Life cycle progression and sexual development of the apicomplexan parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.M.; Franke-Fayard, B.; Mair, G.R.; Lasonder, E.; Janse, C.J.; Mann, M.; Waters, A.P. Proteome analysis of separated male and female gametocytes reveals novel sex-specific Plasmodium biology. Cell 2005, 121, 675–687. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Chen, Z.; Wang, Z.; Shrestha, S.; Li, X.; Li, R.; Cui, L. Sex-Specific Biology of the Human Malaria Parasite Revealed from the Proteomes of Mature Male and Female Gametocytes. Mol. Cell. Proteom. 2017, 16, 537–551. [Google Scholar] [CrossRef] [Green Version]
- Tomasina, R.; Francia, M.E. The Structural and Molecular Underpinnings of Gametogenesis in Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2020, 10, 608291. [Google Scholar] [CrossRef]
- Straschil, U.; Talman, A.M.; Ferguson, D.J.P.; Bunting, K.A.; Xu, Z.; Bailes, E.; Sinden, R.E.; Holder, A.A.; Smith, E.F.; Coates, J.C.; et al. The Armadillo Repeat Protein PF16 Is Essential for Flagellar Structure and Function in Plasmodium Male Gametes. PLoS ONE 2010, 5, e12901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olliaro, P.L.; Taylor, W.R. Antimalarial compounds: From bench to bedside. J. Exp. Biol. 2003, 206, 3753–3759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, V.R.; Curran, M.P. Nitazoxanide: A review of its use in the treatment of gastrointestinal infections. Drugs 2007, 67, 1947–1967. [Google Scholar] [CrossRef]
- Leport, C.; Raffi, F.; Matheron, S.; Katlama, C.; Regnier, B.; Saimot, A.G.; Marche, C.; Vedrenne, C.; Vilde, J.L. Treatment of central nervous system toxoplasmosis with pyrimethamine/sulfadiazine combination in 35 patients with the acquired immunodeficiency syndrome. Efficacy of long-term continuous therapy. Am. J. Med. 1988, 84, 94–100. [Google Scholar] [CrossRef]
- Pawlowic, M.C.; Somepalli, M.; Sateriale, A.; Herbert, G.T.; Gibson, A.R.; Cuny, G.D.; Hedstrom, L.; Striepen, B. Genetic ablation of purine salvage in Cryptosporidium parvum reveals nucleotide uptake from the host cell. Proc. Natl. Acad. Sci. USA 2019, 116, 21160–21165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saldarriaga, J.F.; McEwan, M.; Fast, N.M.; Taylor, F.; Keeling, P.J. Multiple protein phylogenies show that Oxyrrhis marina and Perkinsus marinus are early branches of the dinoflagellate lineage. Int. J. Syst. Evol. Microbiol. 2003, 53, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Fast, N.M.; Xue, L.; Bingham, S.; Keeling, P.J. Re-examining alveolate evolution using multiple protein molecular phylogenies. J. Eukaryot. Microbiol. 2002, 49, 30–37. [Google Scholar] [CrossRef]
- Keeling, P.J.; Doolittle, W.F. α-tubulin from early-diverging eukaryotic lineages and the evolution of the tubulin family. Mol. Biol. Evol. 1996, 13, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Gaillard, N.; Sharma, A.; Abbaali, I.; Liu, T.; Shilliday, F.; Cook, A.D.; Ehrhard, V.; Bangera, M.; Roberts, A.J.; Moores, C.A.; et al. Inhibiting parasite proliferation using a rationally designed anti-tubulin agent. EMBO Mol. Med. 2021, 13, e13818. [Google Scholar] [CrossRef]
- Ullah, I.; Gahalawat, S.; Booshehri, L.M.; Niederstrasser, H.; Majumdar, S.; Leija, C.; Bradford, J.M.; Hu, B.; Ready, J.M.; Wetzel, D.M. An Antiparasitic Compound from the Medicines for Malaria Venture Pathogen Box Promotes Leishmania Tubulin Polymerization. ACS Infect. Dis. 2020, 6, 2057–2072. [Google Scholar] [CrossRef] [PubMed]
- Parker, A.L.; Teo, W.S.; McCarroll, J.A.; Kavallaris, M. An Emerging Role for Tubulin Isotypes in Modulating Cancer Biology and Chemotherapy Resistance. Int. J. Mol. Sci. 2017, 18, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrissette, N.S.; Mitra, A.; Sept, D.; Sibley, L.D. Dinitroanilines bind α-tubulin to disrupt microtubules. Mol. Biol. Cell 2004, 15, 1960–1968. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | α-Tubulin | β-Tubulin | γ-Tubulin |
|---|---|---|---|
| Babesia microti | BmR1_04g09455 Ch IV BMR1_02g00915 1 Ch II | BMR1_01G01045 7 Ch I | BmR1_04g07605 10 Ch IV |
| Besnoitia besnoitia | BESB_078340 Ch VII BESB_039600 2 Ch II BESB_009760 Ch I BESB_009930 Ch I | BESB_049820 Ch III BESB_055160 Ch IV BESB_080330 Ch VII | BESB_059730 11 Ch V |
| Cryptosporidium parvum | cgd4_2860 Ch 4 | cgd6_4760 Ch 6 | cgd7_1980 12 Ch 7 |
| Cyclospora cayetanensis | cyc_07446 NA cyc_01940 3 NA | cyc_02710 NA cyc_00127 NA | cyc_08632 13 NA |
| Cystoisospora suis | CSUI_008696 NA CSUI_001393 NA CSUI_005294 4 NA | CSUI_006267 NA CSUI_009771 NA CSUI_006169 NA | CSUI_002962 NA |
| Eimeria tenella | ETH2_1417400 Ch 14 ETH2_0609600 5 Ch 6 | ETH2_0401900 Ch 4 | ETH2_1576800 14 Ch 15 |
| Hammondia hammondia | HHA_316400 NA HHA_231770 NA HHA_231400 NA | HHA_266960 NA HHA_221620 NA HHA_212240 NA | HHA_226870 NA |
| Neospora caninum | NCLIV_058890 Ch XI NCLIV_031800 6 Ch VIII NCLIV_031660 Ch VIII | NCLIV_039100 Ch IX NCLIV_005150 Ch II NCLIV_049140 Ch X | NCLIV_046130 Ch X |
| Plasmodium falciparum | PF3D7_0903700 Ch 9 PF3D7_0422300 Ch 4 | PF3D7_1008700 Ch 10 | PF3D7_0803700 Ch 8 |
| Plasmodium vivax | PVP01_0702100 Ch 7 PVP01_0530800 Ch 5 | PVP01_0808400 Ch 8 | PVP01_0116500 Ch 1 |
| Sarcocystis neurona | SN3_01900485 NA | SN3_00103320 NA SN3_02800250 8 NA | SN3_01300465 15 NA |
| Theileria annulata | TA08335 Ch 4 TA21240 Ch 1 | TA13315 9 Ch 2 | TA03470 16 Ch 3 |
| Toxoplasma gondii | TGME49_316400 Ch XI TGME49_231770 Ch VIII TGME49_231400 Ch VIII | TGME49_266960 Ch IX TGME49_221620 Ch II TGME49_212240 Ch X | TGME49_226870 Ch X |
| Species | δ-Tubulin | ε-Tubulin |
|---|---|---|
| Babesia microti | BMR1_02g03305 Ch II | BMR1_03g04130 Ch III |
| Besnoitia besnoitia | BESB_022320 Ch XII | BESB_029970 Ch XIII |
| Cryptosporidium parvum | Not identified | Not identified |
| Cyclospora cayetanensis | cyc_04504 1 NA | cyc_05034 NA |
| Cystoisospora suis | CSUI_009859 2 NA | CSUI_001319 NA |
| Eimeria tenella | ETH2_1040000 Ch 10 | ETH2_0530700 Ch 5 |
| Hammondia hammondia | HHA_207600 NA | HHA_275870 NA |
| Neospora caninum | NCLIV_002570 3 Ch Ib | NCLIV_007070 Ch III |
| Plasmodium falciparum | PF3D7_0933800 4 Ch 9 | PF3D7_1475700 Ch 14 |
| Plasmodium vivax | PVP01_0732500 5 Ch 7 | PVP01_1270600 Ch 12 |
| Sarcocystis neurona | SN3_00102395 6 NA | SN3_01100650 7 NA |
| Theileria annulata | Not identified | Not identified |
| Toxoplasma gondii | TGME49_207600 Ch Ib | TGME49_275870 8 Ch III |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrissette, N.; Abbaali, I.; Ramakrishnan, C.; Hehl, A.B. The Tubulin Superfamily in Apicomplexan Parasites. Microorganisms 2023, 11, 706. https://doi.org/10.3390/microorganisms11030706
Morrissette N, Abbaali I, Ramakrishnan C, Hehl AB. The Tubulin Superfamily in Apicomplexan Parasites. Microorganisms. 2023; 11(3):706. https://doi.org/10.3390/microorganisms11030706
Chicago/Turabian StyleMorrissette, Naomi, Izra Abbaali, Chandra Ramakrishnan, and Adrian B. Hehl. 2023. "The Tubulin Superfamily in Apicomplexan Parasites" Microorganisms 11, no. 3: 706. https://doi.org/10.3390/microorganisms11030706