

Effective Biocorrosive Control in Oil Industry Facilities: 16S rRNA Gene Metabarcoding for Monitoring Microbial Communities in Produced Water
 , , , , , ,
, , , , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling of Produced Water (PW)
2.2. Produced Water (PW)
2.3. SRB, APB, and GANB Enrichment Cultures Obtained from PW
2.4. DNA Extraction
2.5. 16S rRNA Gene Amplicon Metagenomic Sequencing
2.6. Bioinformatic Analyses
- If x = 0 and y = 0, then the conditional mean is 0;
- If x = 5 and y = 0, then the conditional mean is 5;
- If x = 0 and y = 8, then the conditional mean is 8;
- If x = 3 and y = 4, then the conditional mean is 3.5 (or 3, since the integer part is used);
- If x = 6 and y = 9, then the conditional mean is 7.5 (or 7, since the integer part is used).
2.7. Integrating Metabarcoding and of PW Samples and Enrichments
2.8. Correlations of the Composition of Microbial Communities to the Physicochemical Parameters of PW
3. Results
3.1. MPN Methods
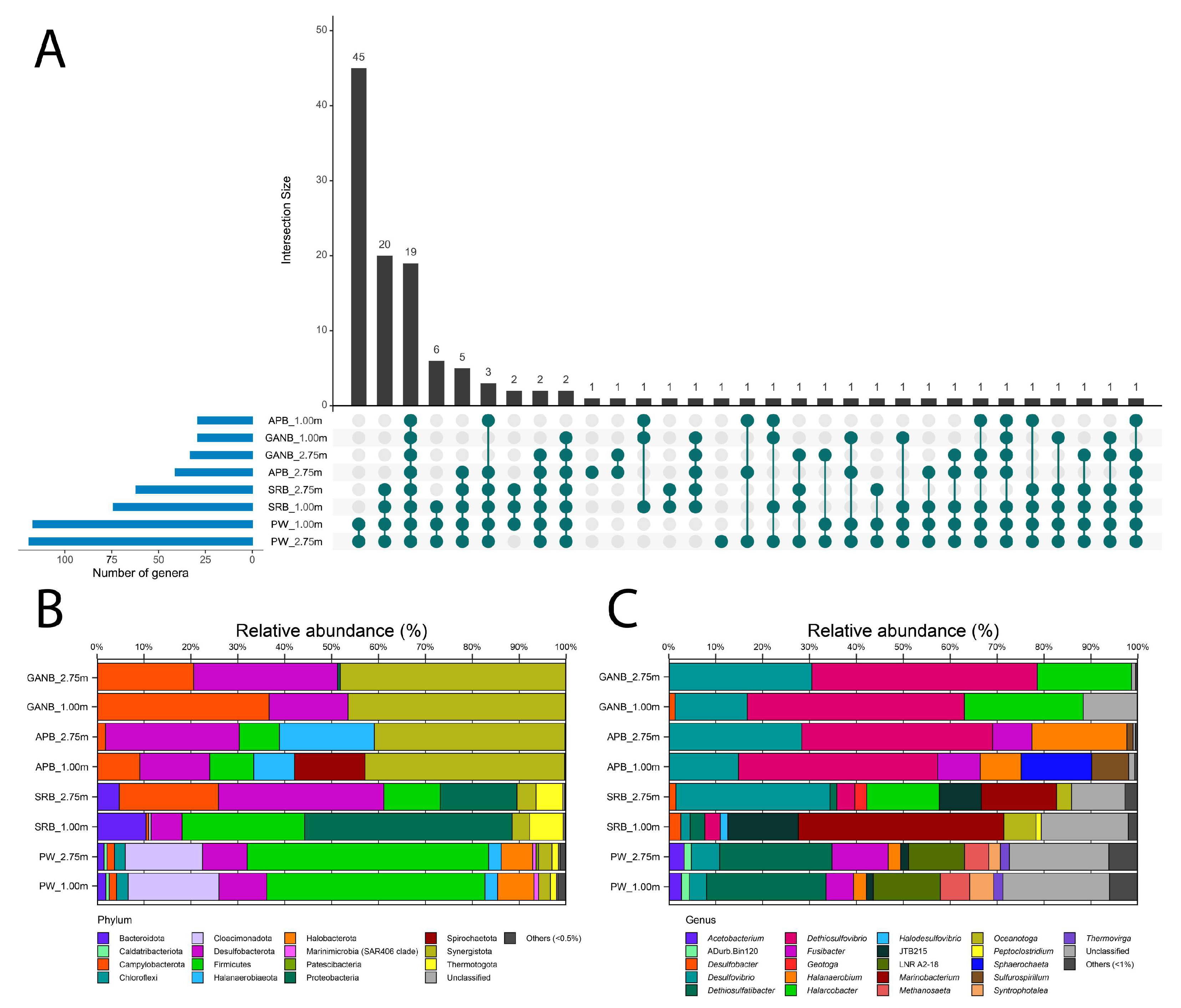
3.2. Shareability and Uniqueness Patterns and Relative Abundance of Phyla and Genera Revealed by Metabarcoding in PW and Enrichments
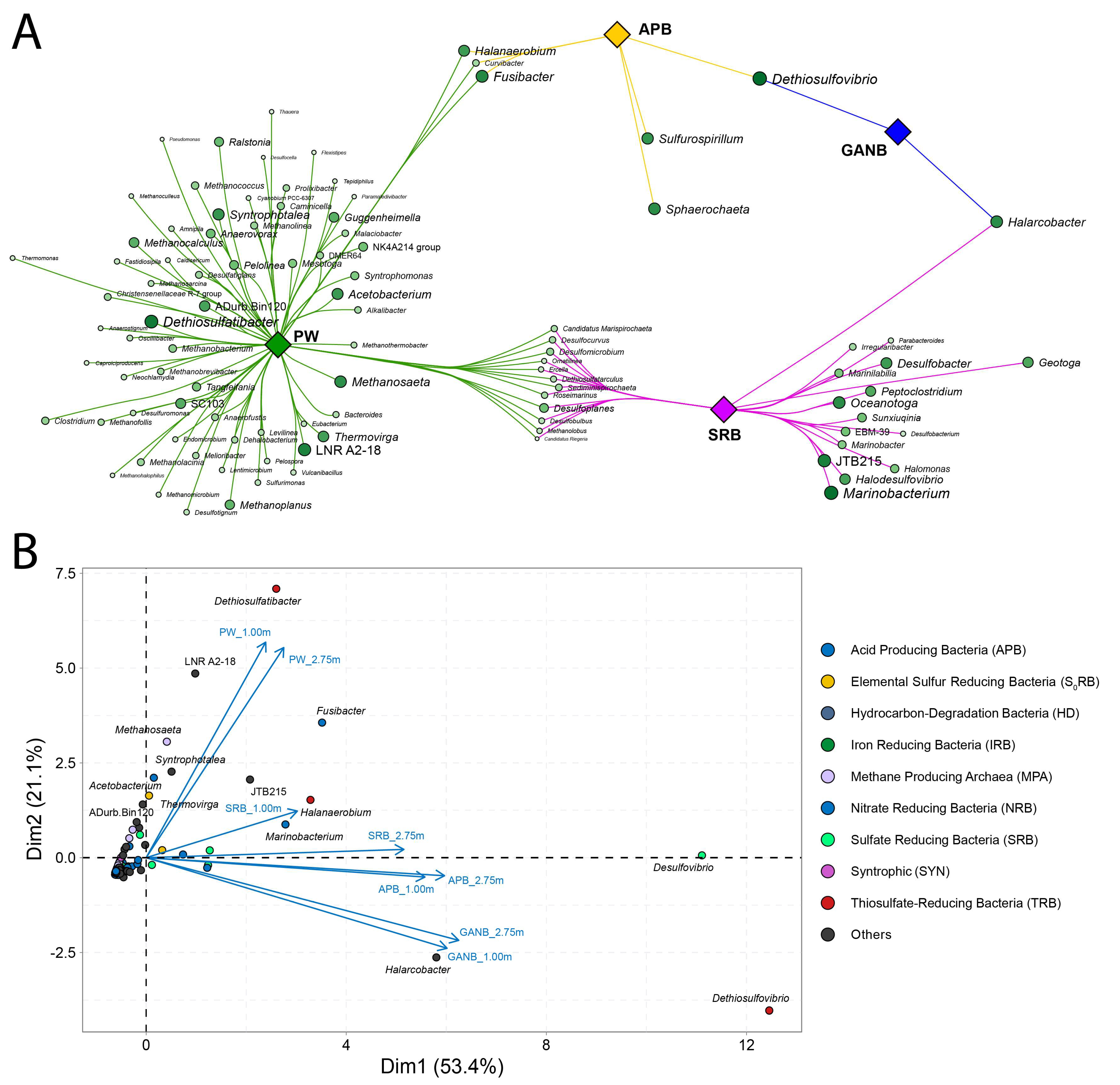
3.3. Microbial Diversity of PW and Enrichments
3.3.1. Statistical Evaluation of Patterns among Microbial Communities Directly Retrieved from PW and Enrichment Cultures
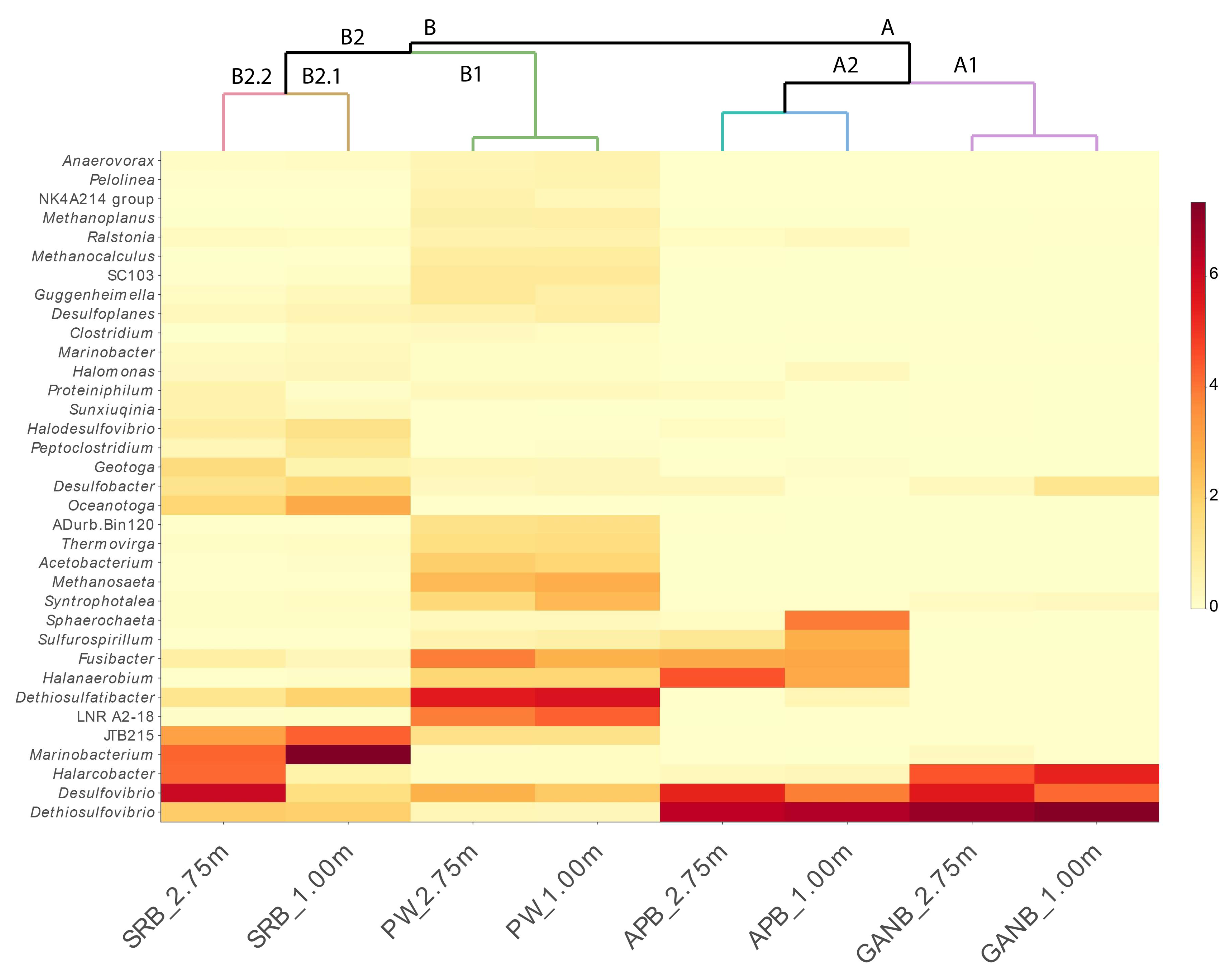
3.3.2. Cluster Analysis in Q and R Modes
3.4. Physicochemical Characterization
3.4.1. Physicochemical Features of PW
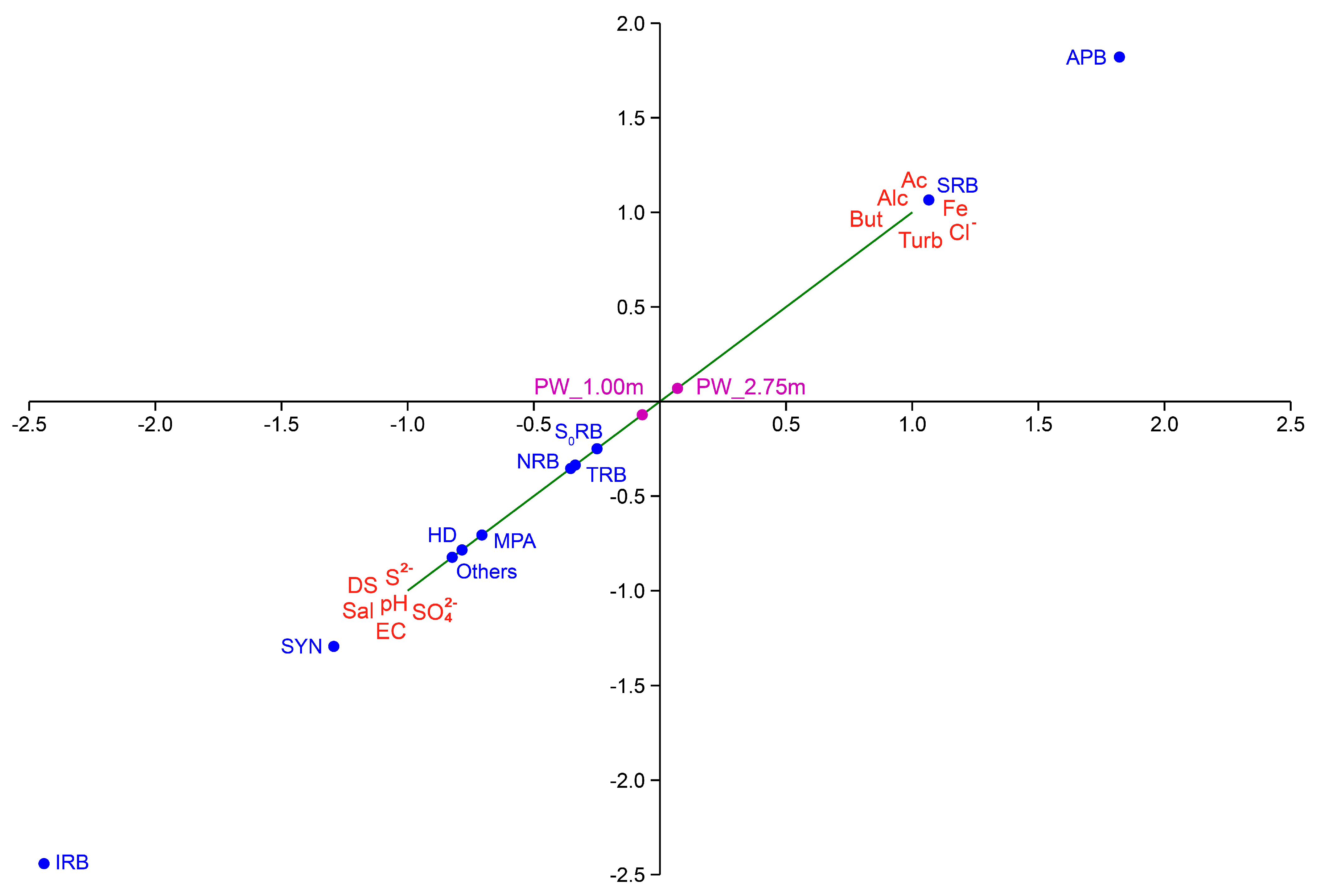
3.4.2. Associations between the Physicochemical Features and the Microorganisms Present in PW Environmental Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajasekar, A.; Anandkumar, B.; Maruthamuthu, S.; Ting, Y.P.; Rahman, P.K.S.M. Characterization of Corrosive Bacterial Consortia Isolated from Petroleum-Product-Transporting Pipelines. Appl. Microbiol. Biotechnol. 2010, 85, 1175–1188. [Google Scholar] [CrossRef]
- Hashemi, S.J.; Bak, N.; Khan, F.; Hawboldt, K.; Lefsrud, L.; Wolodko, J. Bibliometric Analysis of Microbiologically In Fl Uenced Corrosion (MIC) of Oil and Gas Engineering Systems. Corrosion 2018, 9312, 468–486. [Google Scholar] [CrossRef] [PubMed]
- Conlette, O.C.; Emmanuel, N.E.; Olukayode, A.O. Factors That Influence Methanogenic Activities in a Low Sulfate Oil-Producing Facility. Pet. Sci. Technol. 2018, 36, 688–695. [Google Scholar] [CrossRef]
- Li, Y.; Xu, D.; Chen, C.; Li, X.; Jia, R.; Zhang, D.; Sand, W.; Wang, F.; Gu, T. Anaerobic Microbiologically Influenced Corrosion Mechanisms Interpreted Using Bioenergetics and Bioelectrochemistry: A Review. J. Mater. Sci. Technol. 2018, 34, 1713–1718. [Google Scholar] [CrossRef]
- Qu, Q.; Wang, L.; Li, L.; He, Y.; Yang, M.; Ding, Z. Effect of the Fungus, Aspergillus niger, on the Corrosion Behaviour of AZ31B Magnesium Alloy in Artificial Seawater. Corros. Sci. 2015, 98, 249–259. [Google Scholar] [CrossRef]
- Kato, S. Microbial Extracellular Electron Transfer and Its Relevance to Iron Corrosion. Microb. Biotechnol. 2016, 9, 141–148. [Google Scholar] [CrossRef]
- Birkeland, N.K.; Schönheit, P.; Poghosyan, L.; Fiebig, A.; Klenk, H.P. Complete Genome Sequence Analysis of Archaeoglobus fulgidus Strain 7324 (DSM 8774), a Hyperthermophilic Archaeal Sulfate Reducer from a North Sea Oil Field. Stand. Genomic Sci. 2017, 12, 79. [Google Scholar] [CrossRef]
- Jia, R.; Unsal, T.; Xu, D.; Lekbach, Y.; Gu, T. Microbiologically in Fl Uenced Corrosion and Current Mitigation Strategies: A State of the Art Review. Int. Biodeterior. Biodegradation 2019, 137, 42–58. [Google Scholar] [CrossRef]
- Liu, J.; Wu, J.; Lin, J.; Zhao, J.; Xu, T.; Yang, Q.; Zhao, J.; Zhao, Z.; Song, X. Changes in the Microbial Community Diversity of Oil Exploitation. Genes 2019, 10, 556. [Google Scholar] [CrossRef] [PubMed]
- Dutra, J.; Gomes, R.; Jasper, G.; García, Y.; Xiomara, D.; Cardoso, M.S.; Waldow, V.; Groposo, C. Corrosion-Influencing Microorganisms in Petroliferous Regions on a Global Scale: Systematic Review, Analysis, and Scientific Synthesis of 16S Amplicon Metagenomic Studies. PeerJ 2023, 11, e14642. [Google Scholar] [CrossRef]
- Pfennig, N.; Biebl, H. Desulfuromonas acetoxidans Gen. Nov. and Sp. Nov., a New Anaerobic, Sulfur-Reducing, Acetate-Oxidizing Bacterium. Arch. Microbiol 1976, 110, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Gnansounou, E. Microbial Dynamics in Petroleum Oilfields and Their Relationship with Physiological Properties of Petroleum Oil Reservoirs. Bioresour. Technol. 2017, 245, 1258–1265. [Google Scholar] [CrossRef] [PubMed]
- Xu, D. Microbiologically Influenced Corrosion (MIC) Mechanisms and Mitigation; Ohio University: Athens, OH, USA, 2013. [Google Scholar]
- Liang, R.; Grizzle, R.S.; Duncan, K.E.; McInerney, M.J.; Suflita, J.M. Roles of Thermophilic Thiosulfate-Reducing Bacteria and Methanogenic Archaea in the Biocorrosion of Oil Pipelines. Front. Microbiol. 2014, 5, 89. [Google Scholar] [CrossRef] [PubMed]
- Little, B.J.; Lee, J.S. Microbiologically Influenced Corrosion, 1st ed.; Pesce, W.J., Wiley, P.B., Eds.; Wiley-Interscience: Hoboken, NJ, USA, 2007; ISBN 9781626239777. [Google Scholar]
- Roche, M. Corrosion Management: A Key Issue In Pipeline Integrity. In Proceedings of the International Petroleum Tecnology Conference, Dubai, United Arab Emirates, 4–6 December 2007; pp. 1–11. [Google Scholar]
- Lele, P.; Syed, A.H.; Riordon, J.; Mosavat, N.; Guerrero, A.; Fadaei, H.; Sinton, D. Deformation of Microdroplets in Crude Oil for Rapid Screening of Enhanced Oil Recovery Additives Pushan. J. Pet. Sci. Eng. 2018, 165, 298–304. [Google Scholar] [CrossRef]
- Mnif, S.; Bru-adan, V.; Godon, J.; Sayadi, S.; Chamkha, M. Characterization of the Microbial Diversity in Production Waters of Mesothermic and Geothermic Tunisian Oil Fi Elds. J. Basic Microbiol. 2013, 53, 45–61. [Google Scholar] [CrossRef]
- Bedoya, K.; Niño, J.; Acero, J.; Cabarcas, F.; Alzate, J.F. Assessment of the Microbial Community and Biocide Resistance Profile in Production and Injection Waters from an Andean Oil Reservoir in Colombia. Int. Biodeterior. Biodegrad. 2021, 157, 105137. [Google Scholar] [CrossRef]
- Thomas, J.E. Processamento Primário de Fluidos. In Fundamentos do Petróleo; Petrobras, Ed.; Academia: San Francisco, CA, USA, 2001; pp. 255–624. [Google Scholar]
- Machuca, L.; Salgar-Chaparro, S. Complementary DNA/RNA-Based Profiling: Characterization of Corrosive Microbial Communities and Their Functional Profiles in an Oil Production Facility. Front. Microbiol. 2019, 10, 2587. [Google Scholar] [CrossRef]
- Fatah, M.C.; Ismail, M.C.; Wahjoedi, B.A. Effects of Sulphide Ion on Corrosion Behaviour of X52 Steel in Simulated Solution Containing Metabolic Products Species: A Study Pertaining to Microbiologically Influenced Corrosion (MIC). Corros. Eng. Sci. Technol. 2013, 48, 211–220. [Google Scholar] [CrossRef]
- Lv, M.; Du, M. A Review: Microbiologically Influenced Corrosion and the Effect of Cathodic Polarization on Typical Bacteria. Rev. Environ. Sci. Biotechnol. 2018, 17, 431–446. [Google Scholar] [CrossRef]
- Bhagobaty, R.K. Culture Dependent Methods for Enumeration of Sulphate Reducing Bacteria (SRB) in the Oil and Gas Industry. Rev. Environ. Sci. Biotechnol. 2014, 13, 11–16. [Google Scholar] [CrossRef]
- Sakamoto, I.K.; Maintinguer, S.I.; AmâncioVaresche, M.B. Phylogenetic Characterization and Quantification by Most Probable Number of the Microbial Communities of Biomass from the Upflow Anaerobic Sludge Blanket Reactor under Sulfidogenic Conditions. Acta Sci.—Technol. 2019, 41, e39128. [Google Scholar] [CrossRef]
- Pedrós-Alió, C. The Rare Bacterial Biosphere. Ann. Rev. Mar. Sci. 2011, 4, 449–466. [Google Scholar] [CrossRef] [PubMed]
- Lomans, B.P.; Global, S.; International, S.; Paula, R.D.; Geissler, B. Proposal of Improved Biomonitoring Standard for Purpose of Microbiologically Influenced Corrosion Risk Assessment. In Proceedings of the Society of Petroleum Enginners, Dubai, United Arab Emirates, 26–28 September 2016; pp. 1–19. [Google Scholar]
- Liduino, V.S.; Cravo-Laureau, C.; Noel, C.; Carbon, A.; Duran, R.; Lutterbach, M.T.; Camporese Sérvulo, E.F. Comparison of Flow Regimes on Biocorrosion of Steel Pipe Weldments: Community Composition and Diversity of Biofilms. Int. Biodeterior. Biodegrad. 2019, 143, 104717. [Google Scholar] [CrossRef]
- Okoro, C.C.; Amund, O.O. Microbial Community Structure of a Low Sulfate Oil Producing Facility Indicate Dominance of Oil Degrading/Nitrate Reducing Bacteria and Methanogens. Pet. Sci. Technol. ISSN 2018, 36, 293–301. [Google Scholar] [CrossRef]
- Okoro, C.C.; Amund, O. Induction of Souring and Corrosion by Anaerobic Microbial Activities in Offshore and Nearshore Oil-Producing Facilities in Nigeria. Pet. Sci. Technol. 2015, 33, 1149–1156. [Google Scholar] [CrossRef]
- Larsen, J.; Sørensen, K.; Højris, B.; Skovhus, T.L. Significance of Troublesome Sulfate-Reducing Prokaryotes (SRP) in Oil Field Systems. In Proceedings of the NACE International Conference & Expo; Division: Houston, TX, USA, 2009; pp. 1–19. [Google Scholar]
- Keasler, V.; Bennett, B.; Keller, C.; Whalen, P.; Cairns, J.; Paula, R.M. De International Biodeterioration & Biodegradation Expanding the Microbial Monitoring Toolkit: Evaluation of Traditional and Molecular Monitoring Methods. Int. Biodeterior. Biodegradation 2013, 81, 51–56. [Google Scholar] [CrossRef]
- Skovhus, T.L.; Eckert, R.B.; Rodrigues, E. Management and Control of Microbiologically Influenced Corrosion (MIC) in the Oil and Gas Industry—Overview and a North Sea Case Study. J. Biotechnol. 2017, 256, 31–45. [Google Scholar] [CrossRef]
- Beale, D.J.; Karpe, A.V.; Jadhav, S.; Muster, T.H.; Palombo, E.A. Omics-Based Approaches and Their Use in the Assessment of Microbial-Influenced Corrosion of Metals. Corros. Rev. 2015, 34, 1–15. [Google Scholar] [CrossRef]
- Handelsman, J.; Rondon, M.R.; Brady, S.F.; Clardy, J.; Goodman, R.M. Molecular Biological Access to the Chemistry of Unknown Soil Microbes: A New Frontier for Natural Products. Chem. Biol. 1998, 5, 5. [Google Scholar] [CrossRef]
- Scholz, M.B.; Lo, C.C.; Chain, P.S.G. Next Generation Sequencing and Bioinformatic Bottlenecks: The Current State of Metagenomic Data Analysis. Curr. Opin. Biotechnol. 2012, 23, 9–15. [Google Scholar] [CrossRef]
- Stark, L.; Giersch, T.; Wünschiers, R. Efficiency of RNA Extraction from Selected Bacteria in the Context of Biogas Production and Metatranscriptomics. Anaerobe 2014, 29, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Petrobras PE-2P&D-02684—Determinação de Ânions Comuns Em Soluções Aquosas Por Cromatografia de Íons 2022, 1–11. Available online: https://bit.ly/3L8t86f (accessed on 15 October 2022).
- Petrobras PE-2P&D-01852—Determinação de Lactato, Acetato, Formato, Propionato e Butirato Em Soluções Aquasas Ppor Cromatografia de Troca Iônica 2020, 1–13. Available online: https://bit.ly/3F9oi56 (accessed on 15 October 2022).
- Petrobras PE-2P&D-02104—Água e Fluidos Aquosos-Determinação de Alcalinidade 2022, 1–14. Available online: https://bit.ly/3Law1ng (accessed on 15 October 2022).
- McCrady, M.H. The Numerical Interpretation of Fermentation-Tube Results. J. Infect. Dis. 1915, 17, 183–212. [Google Scholar] [CrossRef]
- Petrobras PE-2P&D-01590-A—Preparo de Meios de Cultura Postgate“E” Modificado Para Bactérias Redutoras de Sulfato Mesofílicas (M-BRS), 2017. Available online: https://bit.ly/3J1uNbk (accessed on 15 October 2022).
- Petrobras PE-2P&D-01570—Técnica Microbilogia Para Detecção e Quantificação de Micro-Organismo—Bactérias Produtoras de Ácidos (BPA), 2021, 1–9. Available online: https://bit.ly/3Jpi1EJ (accessed on 15 October 2022).
- Petrobras PE-2P&D-01598—Contagem de Bactérias Anaeróbias Heterotróficas Totais (BANHT), 2019. Available online: https://bit.ly/3F4YYgI (accessed on 15 October 2022).
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S RRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2010, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and Clustering Orders of Magnitude Faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet.journal 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Andrews, S. A Quality Control Tool for High Throughput Sequence Data 2010, 4–7. Available online: https://www.bioinformatics.babraham.ac.uk/projects/download.html#fastqc (accessed on 25 November 2022).
- Edgar, R.C. SINTAX: A Simple Non-Bayesian Taxonomy Classifier for 16S and ITS Sequences. biorxiv 2016. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Glo, F.O.; Yarza, P. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Package, T. Package ‘Dplyr’2023, 1–104. Available online: https://dplyr.tidyverse.org (accessed on 25 November 2022).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Melo, A.S. O Que Ganhamos ‘Confundindo’ Riqueza de Espécies e Equabilidade Em Um Índice de Diversidade? Biota Neotrop 2008, 8, 22–26. [Google Scholar] [CrossRef]
- Conway, J.R.; Lex, A.; Gehlenborg, N. UpSetR: An R Package for the Visualization of Intersecting Sets and Their Properties. Bioinformatics 2017, 33, 2938–2940. [Google Scholar] [CrossRef] [PubMed]
- Jansen, F.; Dell, N. Package ‘ Indicspecies’. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for Visualization and Analysis of Biological Networks. Data Min. Proteom. Stand. Appl. 2011, 696, 291–303. [Google Scholar]
- Kassambara, A.; Mundt, F. Extract and Visualize the Results of Multivariate Data Analyses. Packag. ‘factoextra’ 2020, 45. Available online: https://cran.r-project.org/web/packages/factoextra/factoextra.pdf (accessed on 25 November 2022).
- Wickham, H. Ggplot2. Wiley Interdiscip. Rev. Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Ripley, B.D. The R Project in Statistical Computing. MSOR Connect. Newsl. LTSN Maths Stats OR Netw. 2001, 1, 23–25. [Google Scholar] [CrossRef]
- Galili, T.; Callaghan, A.O.; Sidi, J.; Sievert, C. Heatmaply: An R Package for Creating Interactive Cluster Heatmaps for Online Publishing. Bioinformatics 2018, 34, 1600–1602. [Google Scholar] [CrossRef]
- Clesceri, L.S.; Greenberg, A.E.; Eaton, A.D.; American Public Health Association (APHA); American Water Works Association (AWWA); Water Environment Federation (WEF). Standard Methods Examination of Water and Wastewater; Water Environment Federation (WEF): Alexandria, VA, USA, 2012.
- Waldow, V.D.A. Microbiologia Do Petróleo e Métodos Moleculares Para Caracterização de Cominudades Microbianas. Available online: https://bit.ly/3FCSuFb (accessed on 7 March 2022).
- Skovhus, T.L.; Enning, D.; Lee, J.S. Microbiologically Influenced Corrosion in the Upstream Oil and Gas Industry; CRC Press: Boca Raton, FL, USA, 2017; ISBN 9781498726603. [Google Scholar]
- Bermont-Bouis, D.; Janvier, M.; Grimont, P.A.D.; Dupont, I.; Vallaeys, T. Both Sulfate-Reducing Bacteria and Enterobacteriaceae Take Part in Marine Biocorrosion of Carbon Steel. J. Appl. Microbiol. 2007, 102, 161–168. [Google Scholar] [CrossRef]
- Conlette, O.C.; Emmanuel, N.E.; Chijoke, O.G. Methanogen Population of an Oil Production Skimmer Pit and the Effects of Environmental Factors and Substrate Availability on Methanogenesis and Corrosion Rates. Microb. Ecol. 2016, 72, 175–184. [Google Scholar] [CrossRef]
- Singh, A.; Müller, B.; Schnürer, A. Profiling Temporal Dynamics of Acetogenic Communities in Anaerobic Digesters Using next—Generation Sequencing and T—RFLP. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, W.; Ji, F.; Wu, J.; Nie, Y.; Ren, C.; Xu, Y. Profiling Prokaryotic Community in Pit Mud of Chinese Strong-Aroma Type Liquor by Using Oligotrophic Culturing. Int. J. Food Microbiol. 2021, 337, 108951. [Google Scholar] [CrossRef] [PubMed]
- Semenova, E.M.; Grouzdev, D.S.; Tourova, T.P.; Nazina, T.N. Physiology and Genomic Characteristics of Geotoga Petraea, a Bacterium Isolated from a Low-Temperature Petroleum Reservoir (Russia). Microbiology 2019, 88, 662–670. [Google Scholar] [CrossRef]
- Sterflinger, K.; Little, B.; Pinar, G.; Pinzari, F.; de los Rios, A.; Gu, J.D. Future Directions and Challenges in Biodeterioration Research on Historic Materials and Cultural Properties. Int. Biodeterior. Biodegrad. 2018, 129, 10–12. [Google Scholar] [CrossRef]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive Functional Profiling of Microbial Communities Using 16S RRNA Marker Gene Sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Crispim, J.S.; Dias, R.S.; Marcus, P.; Vidigal, P.; De Sousa, M.P.; Canêdo, C.; Santana, M.F. Screening and Characterization of Prophages in Desulfovibrio Genomes. Sci. Rep. 2018, 8, 9273. [Google Scholar] [CrossRef]
- Plugge, C.M.; Balk, M.; Stams, A.J.M. Desulfotomaculum thermobenzoicum Subsp. Thermosyntrophicum Subsp. Nov., a Thermophilic, Syntrophic, Propionate-Oxidizing, Spore-Forming Bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 391–399. [Google Scholar]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Sthal, D.A. Microbiologia de Brock, 14th ed.; Fonseca, F.G.D., Ed.; Artmed: Porto Alegre, Brazil, 2016; Volume 13, ISBN 9780321897398. [Google Scholar]
- Köberl, M.; Müller, H.; Ramadan, E.M.; Berg, G. Desert Farming Benefits from Microbial Potential in Arid Soils and Promotes Diversity and Plant Health. PLoS One 2011, 6, e24452. [Google Scholar] [CrossRef]
- Chernicharo, C.A.D.L. Reatores Anaeróbios-Princípios Do Tratamento Biológico de Águas Residuárias, 2nd ed.; Polytécnica: Belo Horizonte, Brazil, 2007. [Google Scholar]
- Gomes, S.D.; Fuess, L.T.; Mañunga, T.; Feitosa de Lima Gomes, P.C.; Zaiat, M. Bacteriocins of Lactic Acid Bacteria as a Hindering Factor for Biohydrogen Production from Cassava Flour Wastewater in a Continuous Multiple Tube Reactor. Int. J. Hydrogen Energy 2016, 41, 8120–8131. [Google Scholar] [CrossRef]
- Koyama, M.; Yamamoto, S.; Ishikawa, K.; Ban, S.; Toda, T. Inhibition of Anaerobic Digestion by Dissolved Lignin Derived from Alkaline Pre-Treatment of an Aquatic Macrophyte. Chem. Eng. J. 2016, 311, 55–62. [Google Scholar] [CrossRef]
- Mand, J.; Park, H.S.; Jack, T.R.; Voordouw, G. The Role of Acetogens in Microbially Influenced Corrosion of Steel. Front. Microbiol. 2014, 5, 268. [Google Scholar] [CrossRef]
- Callado, N.H.; Helena, M.; Damianovic, Z.; Foresti, E. Influência Da Razão DQO/[SO 42-] e Da Concentração de Na + Na Remoção de Matéria Orgânica e Sulfato Em Reator UASB. Eng. Sanit. e Ambient. 2017, 22, 381–390. [Google Scholar] [CrossRef]
- Khalid, A.; Arshad, M.; Anjum, M.; Mahmood, T.; Dawson, L. The Anaerobic Digestion of Solid Organic Waste. Waste Manag. 2011, 31, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ren, N.; Wang, A. Contributions of Fermentative Acidogenic Bacteria and Sulfate-Reducing Bacteria to Lactate Degradation and Sulfate Reduction. Chemosphere 2008, 72, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Grigoryan, A.A.; Cornish, S.L.; Buziak, B.; Lin, S.; Cavallaro, A.; Arensdorf, J.J.; Voordouw, G. Competitive Oxidation of Volatile Fatty Acids by Sulfate- and Nitrate-Reducing Bacteria from an Oil Field in Argentina†. Appl. Environ. Microbiol. 2008, 74, 4324–4335. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, L.P.; Richards, F.D. Oxygenation of Hydrogen Sulfide in Seawater at Constant Salinity, Temperature, and PH. Environ. Sci. Technol. 1969, 3, 838–843. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Unit | p_1.00 | p_2.75 |
|---|---|---|---|
| pH | - | 7.00 | 6.00 |
| Lactate | mg/L | n.d | n.d |
| Acetate | mg/L | 970.00 | 1500.00 |
| Propionate | mg/L | n.d | n.d |
| Formate | mg/L | n.d | n.d |
| Butyrate | mg/L | 29.00 | 67.00 |
| Sulfate (SO42−) | mg/L | 310.00 | 170.00 |
| Soluble sulfides (S2−) | mg/L | 57.80 | 54.90 |
| Chloride (Cl−) | mg/L | 2.60 | 2.80 |
| Iron | mg/L | 0.48 | 0.59 |
| Alkalinity | meqs/L | 25.50 | 28.70 |
| Salinity | PPT | 37.50 | 36.40 |
| Electrical conductivity (EC) | Ms | 65.00 | 60.71 |
| Dissolved solids (DS) | PPT | 31.55 | 30.25 |
| Turbidity (Turb) | NTU | 90.125 | 106.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dutra, J.; García, G.; Gomes, R.; Cardoso, M.; Côrtes, Á.; Silva, T.; de Jesus, L.; Rodrigues, L.; Freitas, A.; Waldow, V.; et al. Effective Biocorrosive Control in Oil Industry Facilities: 16S rRNA Gene Metabarcoding for Monitoring Microbial Communities in Produced Water. Microorganisms 2023, 11, 846. https://doi.org/10.3390/microorganisms11040846
Dutra J, García G, Gomes R, Cardoso M, Côrtes Á, Silva T, de Jesus L, Rodrigues L, Freitas A, Waldow V, et al. Effective Biocorrosive Control in Oil Industry Facilities: 16S rRNA Gene Metabarcoding for Monitoring Microbial Communities in Produced Water. Microorganisms. 2023; 11(4):846. https://doi.org/10.3390/microorganisms11040846
Chicago/Turabian StyleDutra, Joyce, Glen García, Rosimeire Gomes, Mariana Cardoso, Árley Côrtes, Tales Silva, Luís de Jesus, Luciano Rodrigues, Andria Freitas, Vinicius Waldow, and et al. 2023. "Effective Biocorrosive Control in Oil Industry Facilities: 16S rRNA Gene Metabarcoding for Monitoring Microbial Communities in Produced Water" Microorganisms 11, no. 4: 846. https://doi.org/10.3390/microorganisms11040846
APA StyleDutra, J., García, G., Gomes, R., Cardoso, M., Côrtes, Á., Silva, T., de Jesus, L., Rodrigues, L., Freitas, A., Waldow, V., Laguna, J., Campos, G., Américo, M., Akamine, R., de Sousa, M., Groposo, C., Figueiredo, H., Azevedo, V., & Góes-Neto, A. (2023). Effective Biocorrosive Control in Oil Industry Facilities: 16S rRNA Gene Metabarcoding for Monitoring Microbial Communities in Produced Water. Microorganisms, 11(4), 846. https://doi.org/10.3390/microorganisms11040846