
Characterization of Zygosaccharomyces lentus Yeast in Hungarian Botrytized Wines

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Examined Botrytized Wines
2.2. Yeasts Isolation from Aszú Wines
2.3. Examined Yeast Strains
2.4. Molecular Taxonomic Identification
2.5. Molecular Typing of the Strains
2.6. Physiological Characterization of Z. lentus Strains
2.6.1. Assessing the Assimilation of Different Carbon and Nitrogen Sources
2.6.2. Phenotypic Characterization via Drop Tests
2.6.3. Screening Enzymatic Activities
2.6.4. Oenological Characterization
2.7. Data Analyses
3. Results
3.1. Examined Botrytized Wines
3.2. Yeast Strains Isolated from Aszú Wines
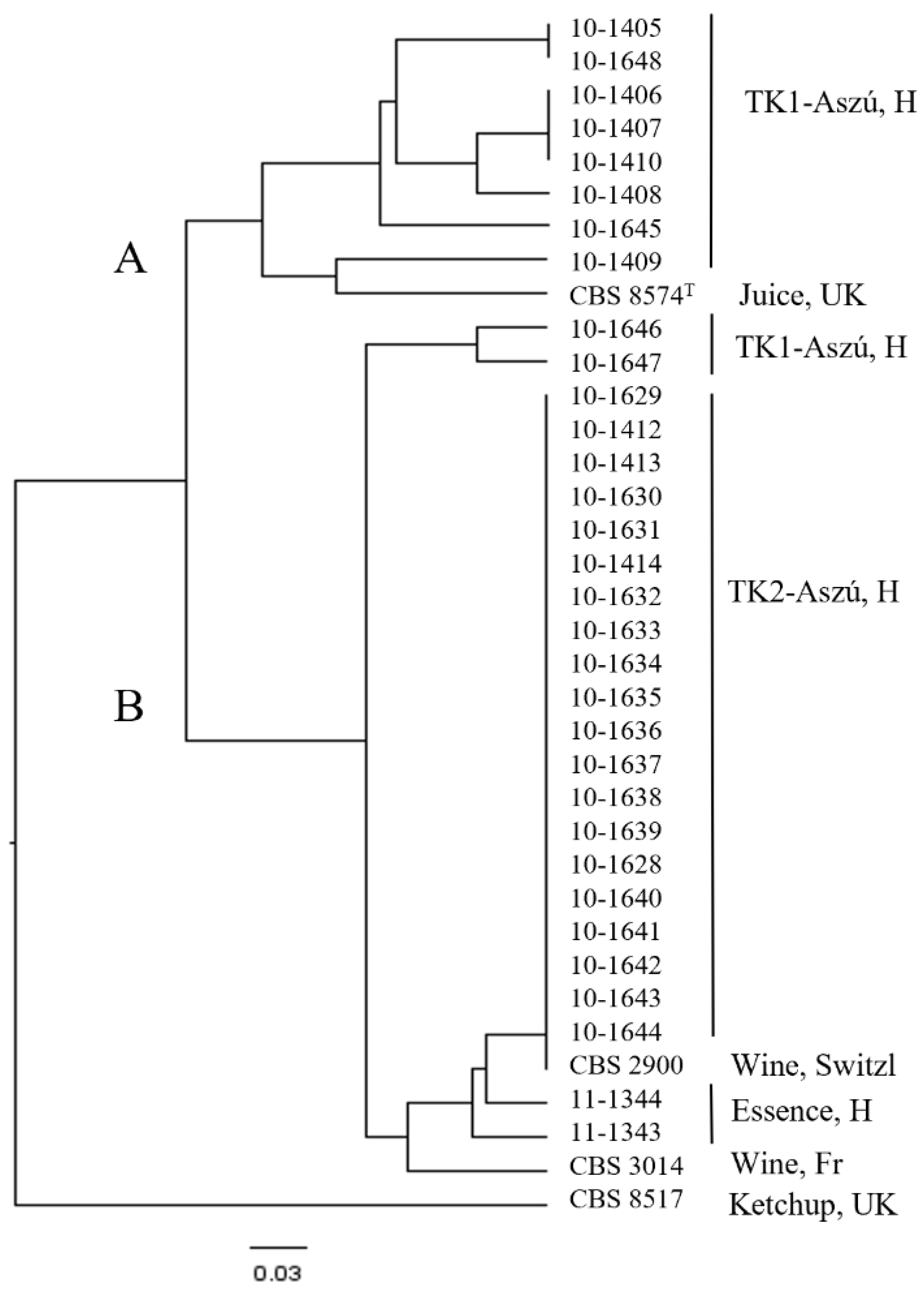
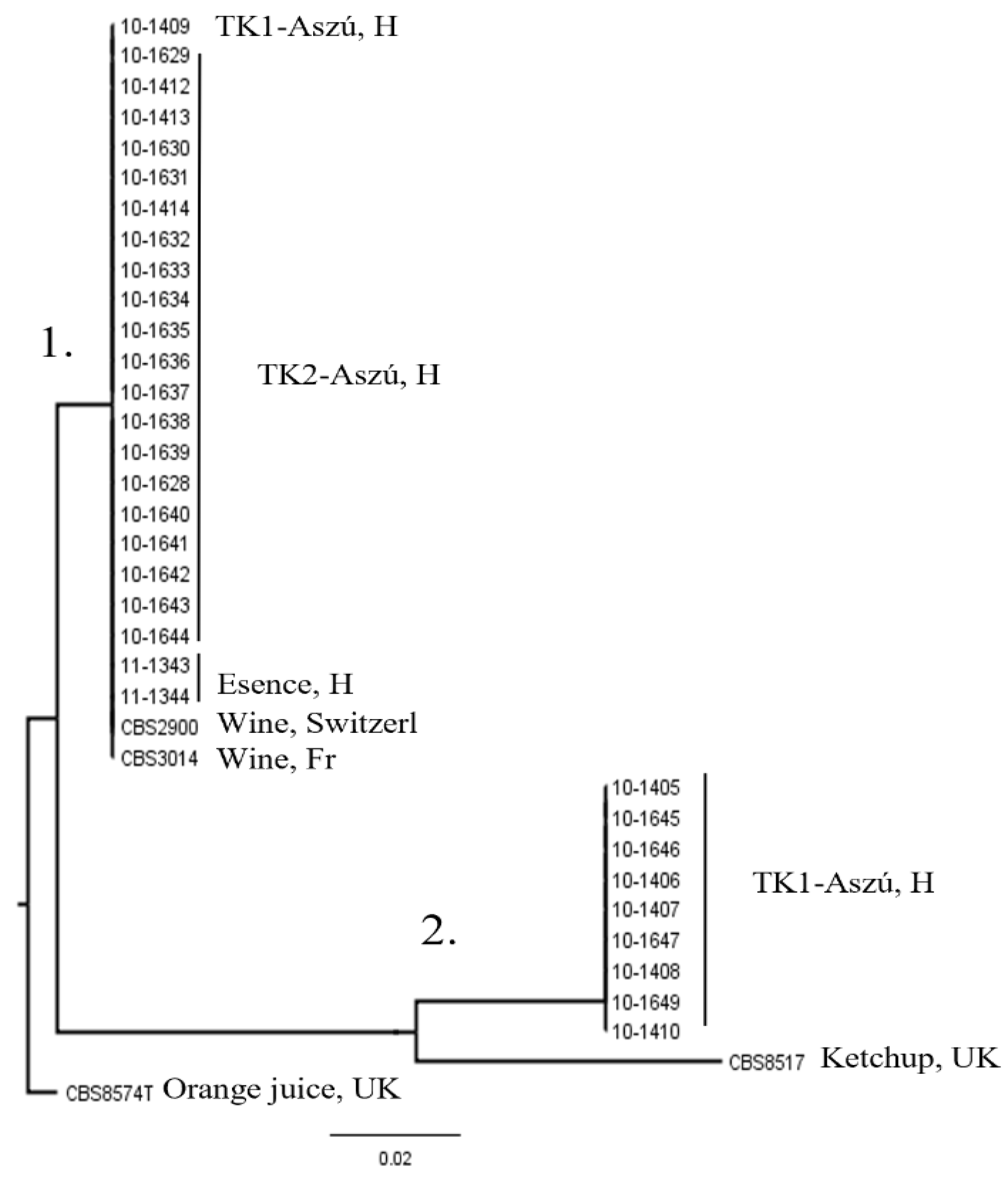
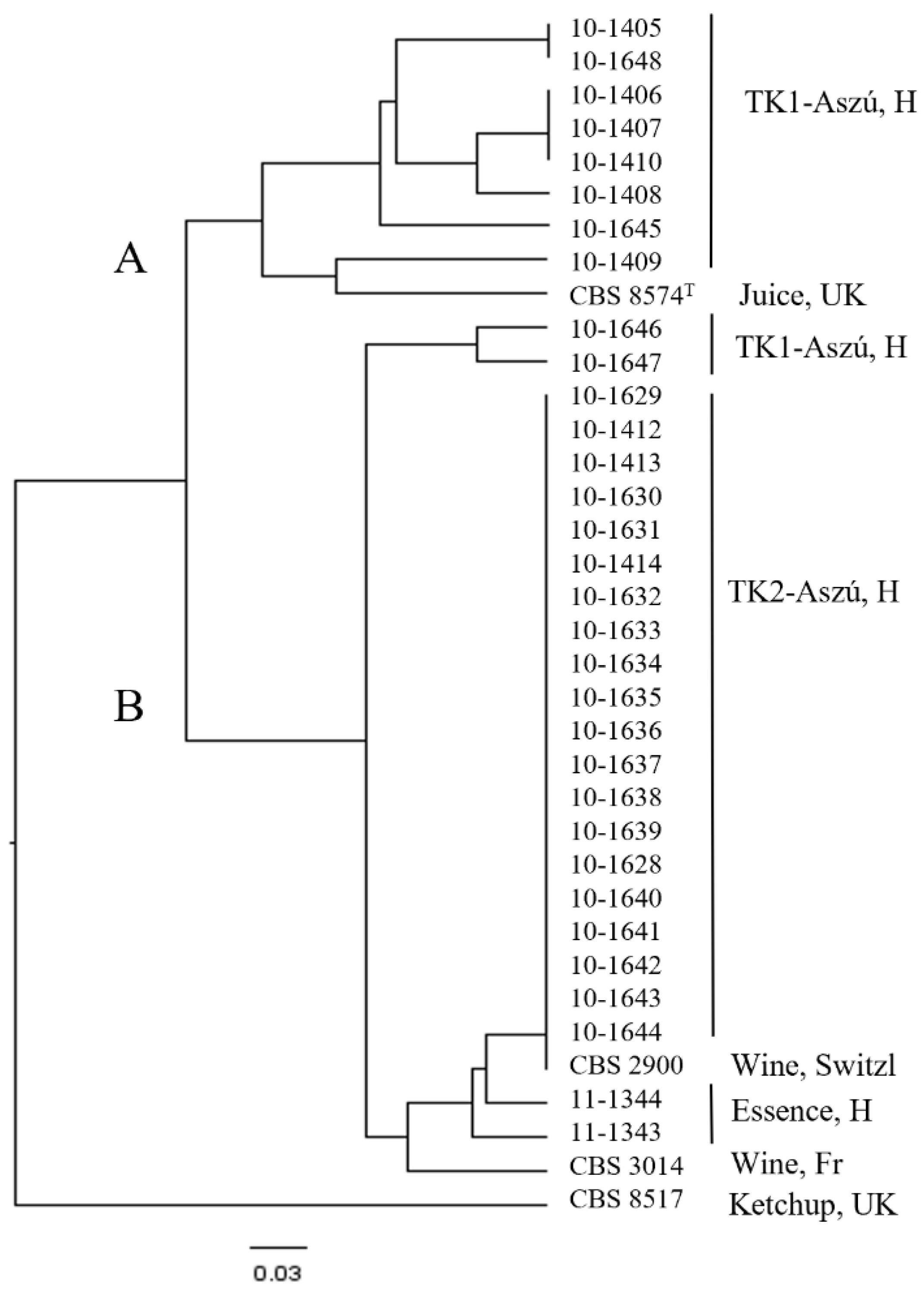
3.3. Molecular Identification
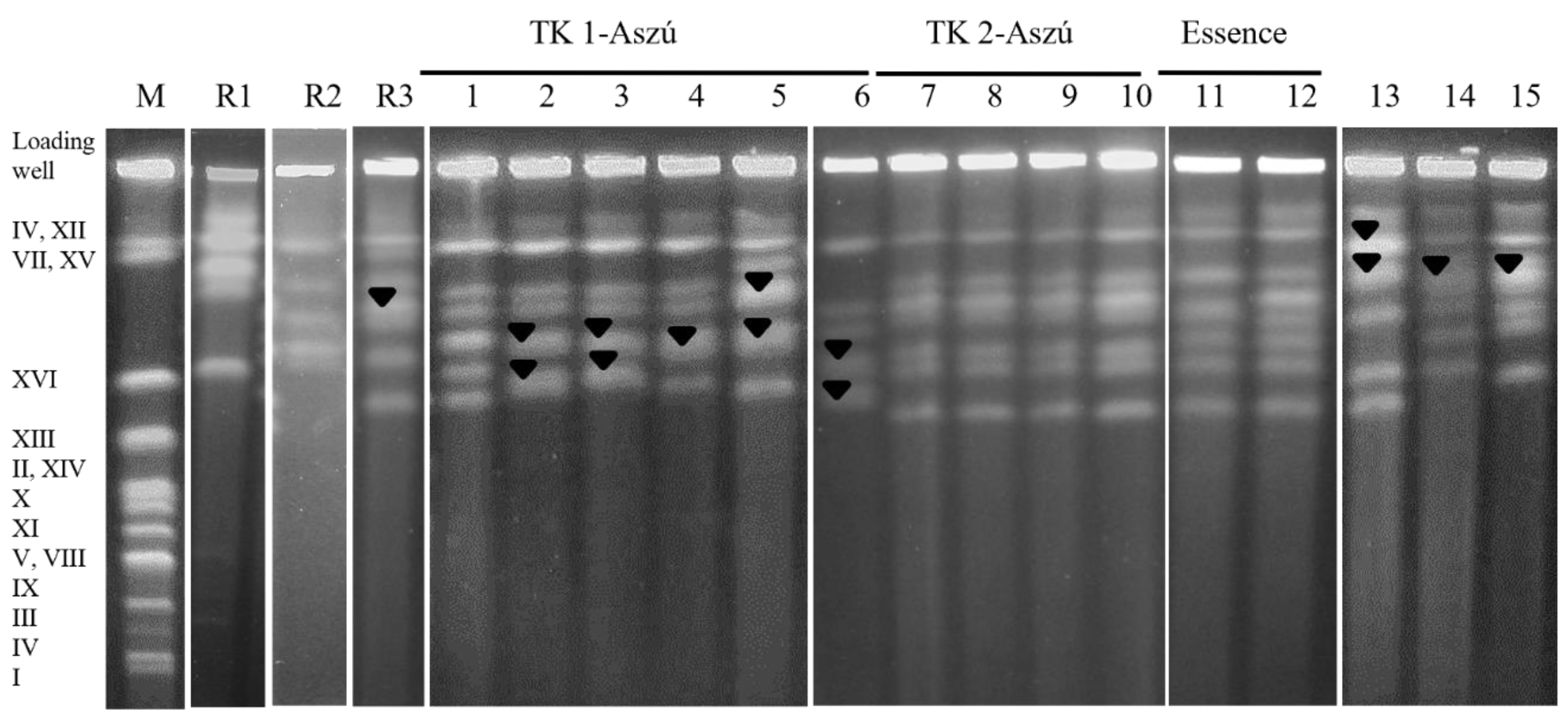
3.4. Molecular Typing of the Strains
3.5. Physiological and Oenological Characterization
4. Discussion
5. Conclusions
- Zygosaccharomyces lentus is a potential contaminant yeast that may persist in the dessert wines we studied, despite high ethanol content and low temperatures, and may cause secondary fermentation during ageing.
- The results we have obtained reveal that the Z. lentus species is heterogeneous in morphological, physiological, and molecular properties.
- Mitochondrial RFLP appears poorly discriminant at the strain level. Our results revealed a high level of similarity in the mitochondrial genome and isolates from wines grouped irrespective of their geographic origins.
- In contrast, greater heterogeneity in the nuclear genome was observed in microsatellite analysis and electrophoretic karyotyping.
- We conclude that it is not possible to determine a species-specific chromosomal pattern characteristic of all strains of the Z. lentus species, because of the diversity in the karyotypes of the strains.
- It was shown that even 50 mg/L of free SO2 is not sufficient to inhibit the growth of Z. lentus.
- The fermentation capacities of the Z. lentus strains were lower than that of the S. cerevisiae starter strain and fermentations were sluggish. Nevertheless, every tested isolate utilized half or more of the starting sugar content and produced varying amounts of alcohol and acetic acid.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sipiczki, M.; Csoma, H.; Antunovics, Z.; Pfliegler, W. Biodiversity in yeast populations associated with botrytised wine making. Mitt. Klosterneubg. 2010, 60, 387–394. [Google Scholar]
- Sipiczki, M. Yeasts in Botrytized Wine Making. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G.H., Eds.; Springer Nature: New York, NY, USA, 2019; pp. 229–261. [Google Scholar]
- Magyar, I. Botrytized wines. Adv. Food Nutr. Res. 2011, 63, 147–206. [Google Scholar] [PubMed]
- Fleet, G.H.; Lafon-Lafourcade, S.; Ribereau-Gayon, P. Evolution of yeasts and lactic acid bacteria during fermentation and storage of Bordeaux wines. Appl. Environ. Microbiol. 1984, 48, 1034–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, D.A.; Eric, A.J.; Cocolin, L. Yeast diversity and persistence in Botrytis-affected wine fermentations. Appl. Environ. Microbiol. 2002, 68, 4884–4893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisiotou, A.A.; Spiropoulos, A.E.; Nychas, G.-J.E. Yeast Community Structures and Dynamics in Healthy and Botrytis-Affected Grape Must Fermentations. Appl. Environ. Microbiol. 2007, 73, 6705–6713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csoma, H.; Sipiczki, M. Taxonomic Investigation of the Yeast Biota of Botrytized Grapes and “Essence” in the Tokaj Wine Region. In Book of Abstracts, 8th International Enology Symposium; Vigne et Vin Publications Internationales: Villenave d’Ornon, France, 2007; p. 174. [Google Scholar]
- Csoma, H.; Kállai, Z.; Antunovics, Z.; Czentye, K.; Sipiczki, M. Vinification without Saccharomyces: Interacting osmotolerant and “spoilage” yeast communities in fermenting and ageing botrytised high-sugar wines (Tokaj Essence). Microorganisms 2021, 9, 19. [Google Scholar] [CrossRef]
- Magyar, I.; Kardos, C.; Pomázi, A.; Maráz, A. Dynamics of Saccharomyces and non-Saccharomyces yeast populations during fermentation of Tokaj Aszú wines. Oenologie 99. 6e Symposium International d’Oenologie. In Proceedings of the “Oenologie 99”, Bordeaux, France, 10–12 June 1999; pp. 394–398. [Google Scholar]
- Sipiczki, M.; Romano, P.; Lipani, G.; Miklos, I.; Antunovics, Z. Analysis of yeasts derived from natural fermentation in a Tokaj winery. Antonie Van Leeuwenhoek 2001, 79, 97–105. [Google Scholar] [CrossRef]
- Sipiczki, M. Candida zemplinina sp. nov., an osmotolerant and psychrotolerant yeast that ferments sweet botrytized wines. Int. J. Syst. Evol. Microbiol. 2003, 53, 2079–2083. [Google Scholar] [CrossRef] [Green Version]
- Antunovics, Z.; Csoma, H.; Sipiczki, M. Molecular and genetic analysis of the yeast flora of botrytized Tokaj wines. Bulletin de l’OIV 2003, 76, 380–397. [Google Scholar]
- Torriani, S.; Lorenzini, M.; Salvetti, E.; Felis, G.E. Zygosaccharomyces gambellarensis sp. nov., an ascosporogenous yeast isolated from an Italian ’passito’ style wine. Int. J. Syst. Evol. Microbiol. 2011, 61, 3084–3088. [Google Scholar] [CrossRef] [Green Version]
- Lleixà, J.; Kioroglou, D.; Mas, A.; del Carmen Portillo, M. Microbiome dynamics during spontaneous fermentations of sound grapes in comparison with sour rot and Botrytis infected grapes. Int. J. Food Microbiol. 2018, 281, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Suzzi, G. Higher alcohol and acetoin production by Zygosaccharomyces wine yeast. J. Appl. Bacteriol. 1993, 75, 543. [Google Scholar] [CrossRef]
- Gobbi, M.; DeVero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative aptitude of non-Saccharomyces wine yeast for reduction in the ethanol content in wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Zhu, X.; Navarro, Y.; Mas, A.; Torija, M.-J.; Beltran, G. A Rapid Method for Selecting Non-Saccharomyces Strains with a Low Ethanol Yield. Microorganisms 2020, 8, 658. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Zorman, T.; Belloch, C.; Querol, A. Molecular characterisation of the species of the genus Zygosaccharomyces system. Appl. Microbiol. 2003, 26, 404–411. [Google Scholar]
- Divol, B.; Lonvaud-Funel, A. Evidence for viable but nonculturable yeasts in Botrytis-affected wine. J. Appl. Microbiol. 2005, 99, 85–93. [Google Scholar] [CrossRef]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology: Volume 1. The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons: Chichester, UK, 2006; p. 497. [Google Scholar]
- Alonso, A.; Belda, I.; Santos, A.; Navascues, E.; Marquina, D. Advances in the control of the spoilage caused by Zygosaccharomyces species on sweet wines and concentrated grape musts. Food Control 2015, 51, 129–134. [Google Scholar] [CrossRef]
- Steels, H.; Bond, C.J.; Collins, M.D.; Roberts, I.N.; Stratford, M.; James, S.A. Zygosaccharomyces lentus sp. nov.; a new member of the yeast genus Zygosaccharomyces Barker. Int. J. Syst. Bacteriol. 1999a, 49, 323. [Google Scholar] [CrossRef] [Green Version]
- Solieri, L. The revenge of Zygosaccharomyces yeasts in food biotechnology and applied microbiology. World J. Microbiol. Biotechnol. 2021, 37, 96. [Google Scholar] [CrossRef]
- Martorell, P.; Stratford, M.; Steels, H.; Fernandez-Espinar, M.T.; Querol, A. Physiological characterization of spoilage strains of Zygosaccharomyces bailii and Zygosaccharomyces rouxii isolated from high sugar environments. Int. J. Food Microbiol. 2007, 114, 234–242. [Google Scholar] [CrossRef]
- Péter, G. Biodiversity of Zygosaccharomyces species in food systems. Acta Aliment. 2022, 51, 43–51. [Google Scholar] [CrossRef]
- Commission Regulation (EEC) No 2676/90 of 17 September 1990 Determining Community Methods for the Analysis of Wines; OJ L 272 03.10.1990, p. 1. Available online: http://data.europa.eu/eli/reg/1990/2676/oj (accessed on 21 February 2020).
- Querol, A.; Barrio, E.; Ramon, D. A comparative study of the different methods of yeast strain characterization. Syst. Appl. Microbiol. 1992, 15, 439–446. [Google Scholar] [CrossRef]
- O’Donnell, K. Fusarium and Its near Relatives. In The Fungal Holomorph: Mitotic; Meiotic and Pleomorphic Speciation in Fungal Systematics; Reynolds, D.R., Taylor, J.W., Eds.; CAB International: Wallingford, UK, 1993; pp. 225–233. [Google Scholar]
- Barnett, J.A.; Payne, R.W.; Yarrow, D. Yeasts: Characteristics and Identification, 2nd ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Belarbi, M.A.; Lemaresquier, M.H. La caratterizzazione dei lieviti. Vignevini 1994, 21, 57–59. [Google Scholar]
- Ganga, M.A.; Martínez, C. Effect of wine yeast monoculture practice on the biodiversity of non-Saccharomyces yeasts. J. Appl. Microbiol. 2004, 96, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Arévalo-Villena, M.; Úbeda Iranzo, J.F.; Briones Pérez, M.I. β-Glucosidase activity in wine yeasts: Application in enology. Enzyme Microb. Technol. 2007, 40, 420–425. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Torchio, F.; Rolle, L.; Gerbi, V.; Cocolin, L. Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: Physiological and molecular characterizations. Int. J. Food Microbiol. 2015, 199, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wickerham, L.J. Taxonomy of Yeasts; U.S. Deptartment of Agriculture: Washington, DC, USA, 1951.
- OIV-MA-AS323-04A: R2009 Standard; Sulfur Dioxide (Titrimetry). Compendium of International Methods of Wine and Must Analysis. OIV: Paris, France, 2016; Volume II, ISBN 979-10-91799-48-5.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef]
- Howell, K. Spoilage: Yeast Spoilage of Food and Beverages. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 5, pp. 113–117. [Google Scholar]
- Magyar, I.; Bene, Z. Morphological and taxonomic study on mycobiota of noble rotted grapes in the Tokaj wine district. Acta Aliment. 2006, 35, 237–246. [Google Scholar] [CrossRef]
- Nisiotou, A.A.; Nychas, G.J. Yeast populations residing on healthy or Botrytis-infected grapes from a vineyard in Attica, Greece. Appl. Environ. Microbiol. 2007, 73, 2765–2768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.; Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. Evaluation of the inhibitory effect of dimethyl dicarbonate (DMDC) against wine microorganisms. Food Microbiol. 2008, 25, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. Changes in sour rotten grape berry microbiota during ripening and wine fermentation. Int. J. Food Microbiol. 2012, 154, 152–161. [Google Scholar] [CrossRef] [PubMed]
- James, S.A.; Stratford, M. Spoilage Yeasts with Emphasis on the Genus Zygosaccharomyces. In Yeasts in Food; Boekhout, T., Robert, V., Eds.; Behr’s Verlag: Hamburg, Germany, 2003; pp. 171–191. [Google Scholar]
- Deák, T.; Reichart, O. Unaceptable Levels of Yeasts in Bottled Wine. In Methods for the Mycological Examination of Food; King, A., Pitt, J., Beuchat, L., Corry, J., Eds.; Plenum Press: New York, NY, USA, 1986; pp. 215–218. [Google Scholar]
- Sá-Correia, I.; Guerreiro, J.F.; Loureiro-Dias, M.C.; Leão, C.; Côrte-Real, M. Zygosaccharomyces. In Encyclopedia of Food Microbiology; Batt, C.A., Tortorello, M.L., Eds.; Elsevier Ltd.: Amsterdam, The Netherlands; Academic Press: New York, NY, USA, 2014; Volume 3, pp. 849–855. [Google Scholar]
- Steels, H.; James, S.A.; Roberts, I.N.; Stratford, M. Zygosaccharomyces lentus: A significant new osmophilic; preservative-resistant spoilage yeast; capable of growth at low temperature. J. Appl. Microbiol. 1999b, 87, 520–527. [Google Scholar] [CrossRef] [PubMed]
- Malfeito-Ferreira, M. Spoilage Yeasts in Red Wines. In Red Wine Technology; Morata, A., Ed.; Academic Press: Cambridge, MA, USA, 2019; Chapter 15; pp. 219–235. [Google Scholar]
- Baleiras-Couto, M.M.; Hartog, B.J.; Huis in’t Veld, J.H.J.; Hofstra, H.; van der Vossen, J.M.B.M. Identification of spoilage yeasts in a food-production chain by microsatellite polymerase chain reaction fingerprinting. Food Microbiol. 1996, 13, 59–67. [Google Scholar] [CrossRef]
- Martorell, P.; Fernández-Espinar, M.T.; Querol, A. Molecular monitoring of spoilage yeasts during the production of candied fruits nougats to determine food contamination sources. Int. J. Food Microbiol. 2005, 101, 293–302. [Google Scholar] [CrossRef]
- Solieri, L.; Cassanelli, S.; Croce, M.A.; Giudici, P. Genome size and ploidy level: New insights for elucidating relationships in Zygosaccharomyces species. Fungal Genet. Biol. 2008, 45, 1582–1590. [Google Scholar] [CrossRef]
- Schuller, D.; Valero, E.; Dequin, S.; Casal, M. Survey of molecular methods for the typing of wine yeast strains. FEMS Microbiol. Lett. 2004, 231, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Csoma, H.; Zakany, N.; Capece, A.; Romano, P.; Sipiczki, M. Biological diversity of Saccharomyces yeasts of spontaneously fermenting wines in four wine regions: Comparative genotypic and phenotypic analysis. Int. J. Food Microbiol. 2010, 140, 239–248. [Google Scholar] [CrossRef]
- Jeyaram, K.; Tamang, J.P.; Capece, A.; Romano, P.P. Geographical markers for Saccharomyces cerevisiae strains with similar technological origins domesticated for rice-based ethnic fermented beverages production in North East India. Antonie van Leeuwenhoek 2011, 100, 569–578. [Google Scholar] [CrossRef]
- Dizzy, M.; Bisson, L.F. Proteolytic activity of yeast strains during grape juice fermentation. Am. J. Enol. Vitic. 2000, 51, 155–167. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Rodríguez Assaf, L.A.; Toro, M.E.; Nally, M.C.; Vallejo, M.; Castellanos de Figueroa, L.I.; Combina, M.; Vazquez, F. Multi-enzyme production by pure and mixed cultures of Saccharomyces and non-Saccharomyces yeasts during wine fermentation. Int. J. Food Microbiol. 2012, 155, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; van Resemburg, P. Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Ough, C.S.; Were, L. Sulfur Dioxide and Sulfites. In Antimicrobials in Food, 3rd ed.; Davidson, P.M., Sofos, J.N., Branen, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 143–167. [Google Scholar]
- Donéche, B.J. Botrytized Wines. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 327–351. [Google Scholar]
- Henick-Kling, T.; Edinger, W.; Daniel, P.; Monk, P. Selective effects of sulfur dioxide and yeast starter culture addition on indigenous yeast populations and sensory characteristics of wine. J. Appl. Microbiol. 1998, 84, 865–876. [Google Scholar] [CrossRef]
- Henschke, P.A.; Jiranek, V. Yeasts-Metabolism of Nitrogen Compounds. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic: Chur, Switzerland, 1993; pp. 77–164. [Google Scholar]
- Office Internationale de la Vigne et du Vin. International Code of Oenological Practices; OIV: Paris, France, 2010; p. 274. [Google Scholar]
- Bely, M.; Rinaldi, A.; Dubourdieu, D. Influence of assimilable nitrogen on volatile acidity production by Saccharomyces cerevisiae during high sugar fermentation. J. Biosci. Bioeng. 2003, 96, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H.; Heard, G.M. Yeasts Growth during Fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- Capozzi, V.; Garofalo, C.; Chiriatti, M.A.; Grieco, F.; Spano, G. Microbial terroir and food innovation: The case of yeast biodiversity in wine. Microbiol. Res. 2015, 181, 75–83. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Another Identifier | Source, Country | Yeast Strains a |
|---|---|---|---|
| S. cerevisiae | YGSC X4005-11A | S.288c | |
| S. cerevisiae | Lalvin EC1118 | Wine, France | 10-1411 |
| Z. bailii | CBS 680T | Unknown | 10-1428 |
| Z. rouxii | CBS 732T | Concentrated black grape must | 10-514 |
| Z. lentus | CBS 8574T | Orange juice, UK | 10-1430 |
| Z. lentus | CBS 2900 | Wine, Switzerland | 10-1429 |
| Z. lentus | CBS 3014 | Wine, France | 10-1426 |
| Z. lentus | CBS 8517 | Ketchup, UK | 10-1427 |
| Z. lentus * | TK1-Aszú wine, Tokaj, Hungary | 10-1405, 10-1406, 10-1407, 10-1408, 10-1409, 10-1410, 10-1645, 10-1646, 10-1647, 10-1648 | |
| Z. lentus * | TK2-Aszú wine, Tokaj, Hungary | 10-1412, 10-1413, 10-1414, 10-1628, 10-1629, 10-1630, 10-1631, 10-1632, 10-1633, 10-1634, 10-1635, 10-1636, 10-1637, 10-1638, 10-1639, 10-1640, 10-1641, 10-1642, 10-1643, 10-1644 | |
| Z. lentus | Essence, Tokaj, Hungary | 11-1343, 11-1344 |
| Samples | Alcohol | Residual Sugar | Total Acidity | pH | SO2 | Acetic Acid | |
|---|---|---|---|---|---|---|---|
| Free | Total | ||||||
| (%, v/v) | (g/L) | (g/L) | (mg/L) | (mg/L) | (g/L) | ||
| TK1-Aszú | 13.38 | 114.50 | 11.80 | 3.35 | 6 | 238 | 1.12 |
| TK2-Aszú | 8.11 | 283.00 | 8.20 | 3.47 | 5 | 45 | 0.93 |
| 11-13-Essence | 5.26 | 638.50 | 9.7 | 3.32 | 12 | 267 | 1.18 |
| Yeast Strain | Fermentative Vigour 1 (5 Days) | Fermentative Vigour 1 (10 Days) | Fermentative Vigour 1 (43 Days) | Reducing Sugars 2 (g/L) | Alcohol 2 (%, v/v) | Total Acidity 2 (g/L) | Acetic Acid 2 (g/L) |
|---|---|---|---|---|---|---|---|
| 10-1405 | 0.15 ± 0.02 b | 0.97 ± 0.40 b | 5.61 ± 0.26 b,d,f,h,i,k,n,p,r,s,u | 85.00 ± 8.76 b,c,f,h,i,k,m,p,q,s | 8.18 ± 0.34 b,d,f,h,j,l,n,p,r,s | 4.46 ± 0.65 a | 0.83 ± 0.18 a,c,f,g,i,k,n,p,r,s,u |
| 10-1406 | 0.15 ± 0.02 b | 0.80 ± 0.37 b | 5.26 ± 0.15 b,d,f,h,i,k,n,p,q,t,u | 106.00 ± 9.0 b,c,e,g,i,k,m,o,q,s | 6.18 ± 0.18 b,d,e,gi,k,m,o,q,t | 4.24 ± 0.27 a | 0.87 ± 0.18 a,c,f,g,i,k,n,p,r,s,u |
| 10-1408 | 0.21 ± 0.13 b | 1.14 ± 0.50 b,d | 6.45 ± 0.21 b,d,f,h,j,l,n,p,r,s | 38.71 ± 0.92 a,d,f,h,j,l,n,p,r | 8.66 ± 0.53 a,d,f,h,j,l,n,p,r,s | 4.62 ± 0.33 a | 0.99 ± 0.12 a,c,e,g,i,l,n,p,r,s |
| 10-1412 | 0.20 ± 0.16 b | 0.96 ± 0.40 b | 4.52 ± 0.13 b,d,e,g,i,k,m,o,q | 97.83 ± 8.52 b,c,e,g,i,k,m,o,q | 6.40 ± 0.42 b,d,e,g,i,k,m,o,q | 3.78 ± 0.39 b,c | 0.42 ± 0.12 b,d,f,h,j,k,o,q |
| 10-1413 | 0.16 ± 0.10 b | 0.85 ± 0.20 b | 4.19 ± 0.65 b,d,e,g,i,l,m,o | 110.33 ± 11.09 b,c,e,g,i,k,m,o | 6.10 ± 0.35 b,d,e,g,i,k,m,o | 3.86 ± 0.30 b,c | 0.33 ± 0.03 b,d,f,h,j,k,m,o |
| 10-1628 | 0.14 ± 0.10 b | 0.72 ± 0.19 b | 4.19 ± 0.24 b,d,e,g,i,k,m | 101.48 ± 6.35 b,c,e,g,i,k,m | 6.25 ± 0.40 b,d,e,g,i,k,m | 3.37 ± 0.02 b | 0.46 ± 0.08 b,d,f,h,j,k,m |
| 11-1343 | 0.15 ± 0.09 b | 1.07 ± 0.65 b | 5.15 ± 0.20 b,d,f,g,i,k | 93.58 ± 6.02 b,c,e,g,i,k | 6.87 ± 0.21 b,d,f,h,i,k | 4.06 ± 0.10 a | 0.57 ± 0.11 b,c,f,h,i,k |
| 11-1344 | 0.17 ± 0.09 b | 0.84 ± 0.11 b | 4.76 ± 0.73 b,d,f,g,i | 95.20 ± 4.30 b,c,e,g,i | 6.55 ± 0.27 b,d,e,g,i | 4.57 ± 0.04 a | 0.86 ± 0.13 a,c,f,g,i |
| CBS 3014 | 0.12 ± 0.08 b | 0.76 ± 0.30 b | 4.25 ± 0.900 b,d,e,g | 114.41 ± 13.09 b,c,e,g | 5.58 ± 0.22 b,c,e,g | 4.26 ± 0.12 a,c | 0.92 ± 0.10 a,c,e,g |
| CBS 2900 | 0.10 ± 0.00 b | 0.55 ± 0.04 b,c | 3.69 ± 0.77 b,c,e | 109.41 ± 1.28 b,c,e | 5.60 ± 0.04 b,c,e | 4.70 ± 0.29 a | 1.28 ± 0.03 a,c,e |
| CBS 8574T | 0.14 ± 0.01 b | 0.71 ± 0.17 b | 3.24 ± 0.32 b,c | 104.25 ± 1.25 b,c | 4.77 ± 0.30 b,c | 4.86 ± 0.20 a | 0.90 ± 0.10 a,c |
| EC1118 | 0.88 ± 0.20 a | 1.89 ± 0.16 a | 8.81 ± 0.62 a | 21.26 ± 10.90 a,d | 9.35 ± 0.39 a | 4.15 ± 0.04 a | 1.09 ± 0.08 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Csoma, H.; Acs-Szabo, L.; Papp, L.A.; Kállai, Z.; Miklós, I.; Sipiczki, M. Characterization of Zygosaccharomyces lentus Yeast in Hungarian Botrytized Wines. Microorganisms 2023, 11, 852. https://doi.org/10.3390/microorganisms11040852
Csoma H, Acs-Szabo L, Papp LA, Kállai Z, Miklós I, Sipiczki M. Characterization of Zygosaccharomyces lentus Yeast in Hungarian Botrytized Wines. Microorganisms. 2023; 11(4):852. https://doi.org/10.3390/microorganisms11040852
Chicago/Turabian StyleCsoma, Hajnalka, Lajos Acs-Szabo, László Attila Papp, Zoltán Kállai, Ida Miklós, and Matthias Sipiczki. 2023. "Characterization of Zygosaccharomyces lentus Yeast in Hungarian Botrytized Wines" Microorganisms 11, no. 4: 852. https://doi.org/10.3390/microorganisms11040852
APA StyleCsoma, H., Acs-Szabo, L., Papp, L. A., Kállai, Z., Miklós, I., & Sipiczki, M. (2023). Characterization of Zygosaccharomyces lentus Yeast in Hungarian Botrytized Wines. Microorganisms, 11(4), 852. https://doi.org/10.3390/microorganisms11040852