
Interkingdom Signaling of the Insect Pathogen Photorhabdus luminescens with Plants Via the LuxR solo SdiA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analyses
2.2. Bacterial Strains, Generation of Deletion Mutants and Expression Vectors
2.3. Motility and Biofilm Assays
2.4. Extraction of Plant Root Exudates (PREs)
2.5. Heterologous Overexpression of SdiA and Purification of 6His-SdiA
2.6. Nano Differential Scanning Fluorimetry (nanoDSF)
2.7. Surface Plasmon Resonance (SPR) Spectroscopy
2.8. Real-Time qPCR (RT-PCR)
2.9. Preparative High-Performance Liquid Chromatography (HPLC)
3. Results and Discussion
3.1. Structural Properties of SdiA
3.2. Influence of SdiA on Motility and Biofilm Formation
3.3. Plant Root Exsudates and AHLs Influence Protein Stability Suggesting Putative Signals for SdiA
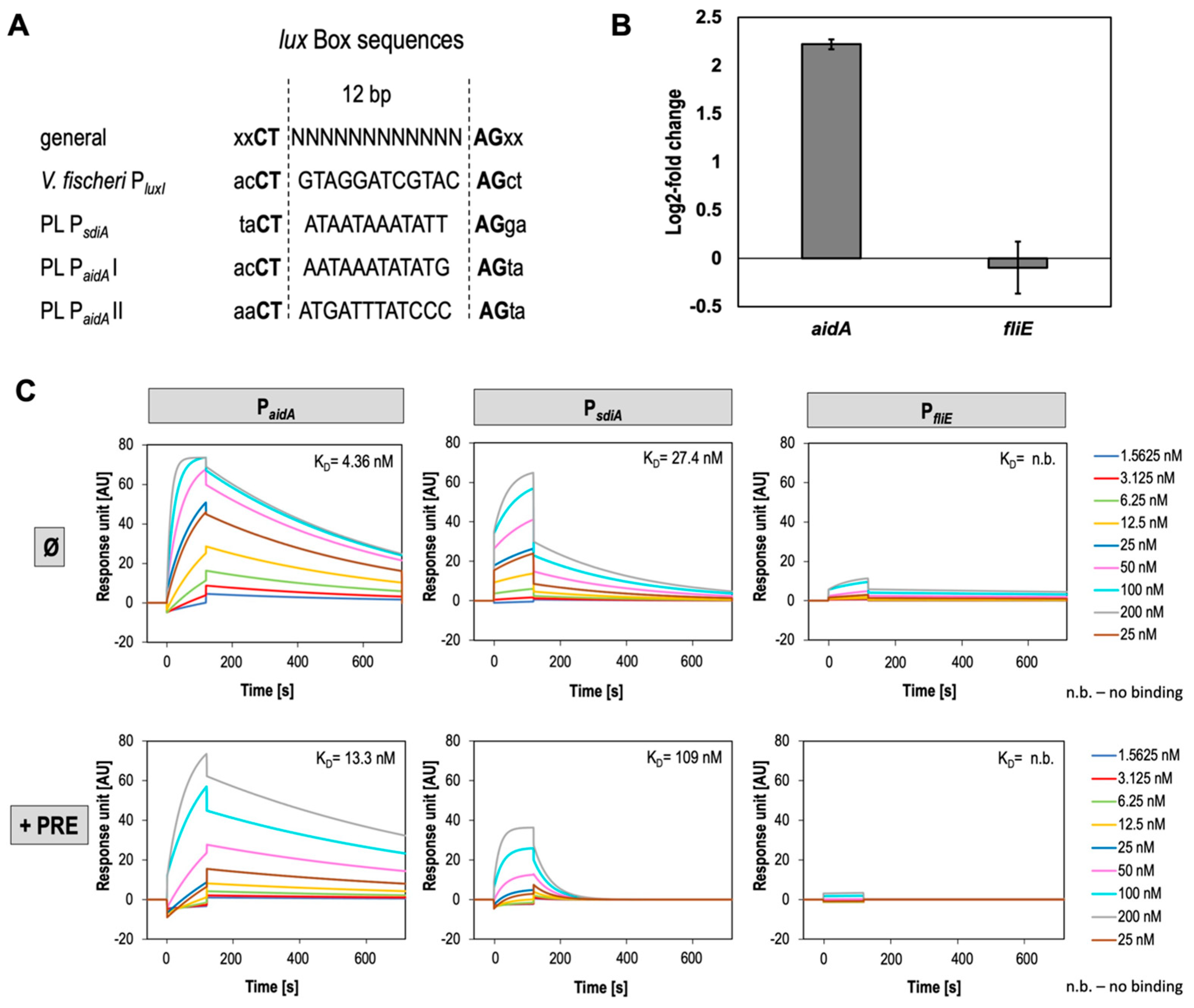
3.4. SdiA Binds PsdiA and PaidA with High Affinity and Negatively Affects Expression of AidA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Greenberg, E.P. The C-terminal region of the Vibrio fischeri LuxR protein contains an inducer-independent lux gene activating domain. PNAS 1991, 88, 11115–11119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanzelka, B.L.; Greenberg, E.P. Evidence That the N-terminal region of the Vibrio fischeri LuxR protein constitutes an autoinducer-binding domain. J. Bacteriol. 1995, 177, 815–817. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Zheng, C.; Chitsaz, F.; Derbyshire, M.K.; Geer, L.Y.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Lanczycki, C.J.; et al. CDD: Conserved domains and protein three-dimensional structure. Nucleic. Acids Res. 2012, 41, D348–D352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuqua, C.; Winans, S.C.; Greenberg, E.P. Census and consensus in bacterial ecosystems: The LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu. Rev. Microbiol. 1996, 50, 727–751. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Fuqua, C.; Parsek, M.R.; Greenberg, E.P. Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet 2001, 35, 439–468. [Google Scholar] [CrossRef]
- Case, R.J.; Labbate, M.; Kjelleberg, S. AHL-driven quorum-sensing circuits: Their frequency and function among the proteobacteria. ISME J. 2008, 2, 345–349. [Google Scholar] [CrossRef]
- Patankar, A.V.; González, J.E. Orphan LuxR regulators of quorum sensing. FEMS Microbiol. Rev. 2009, 33, 739–756. [Google Scholar] [CrossRef] [Green Version]
- Subramoni, S.; Venturi, V. LuxR-family ‘solos’: Bachelor sensors/regulators of signalling molecules. Microbiol 2009, 155, 1377–1385. [Google Scholar] [CrossRef] [Green Version]
- Bez, C.; Geller, A.M.; Levy, A.; Venturi, V. Cell-cell signaling proteobacterial LuxR solos: A treasure trove of subgroups having different origins, ligands, and ecological roles. mSystems 2023, e01039-22. [Google Scholar] [CrossRef] [PubMed]
- Michael, B.; Smith, J.N.; Swift, S.; Heffron, F.; Ahmer, B.M.M. SdiA of Salmonella enterica is a LuxR homolog that detects mixed microbial communities. J. Bacteriol. 2001, 183, 5733–5742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Almeida, F.A.; Pinto, U.M.; Vanetti, M.C.D. Novel insights from molecular docking of SdiA from Salmonella enteritidis and Escherichia coli with quorum sensing and quorum quenching molecules. Microb. Pathog. 2016, 99, 178–190. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Wüthrich, D.; Kuhnert, P.; Arce, C.C.M.; Thönen, L.; Ruiz, C.; Zhang, X.; Robert, C.A.M.; Karimi, J.; Kamali, S.; et al. Whole-genome-based revisit of Photorhabdus phylogeny: Proposal for the elevation of most Photorhabdus subspecies to the species level and description of one novel species Photorhabdus bodei sp. nov. and one novel subspecies Photorhabdus laumondii subsp. clarkei subsp. Nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 2664–2681. [Google Scholar] [CrossRef]
- Heermann, R.; Fuchs, T.M. Comparative Analysis of the Photorhabdus luminescens and the Yersinia enterocolitica genomes: Uncovering candidate genes involved in insect pathogenicity. BMC Genom. 2008, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Brameyer, S.; Kresovic, D.; Bode, H.B.; Heermann, R. LuxR solos in Photorhabdus species. Front Cell Infect. Microbiol. 2014, 4, 166. [Google Scholar] [CrossRef] [Green Version]
- Brachmann, A.O.; Brameyer, S.; Kresovic, D.; Hitkova, I.; Kopp, Y.; Manske, C.; Schubert, K.; Bode, H.B.; Heermann, R. Pyrones as bacterial signaling molecules. Nat. Chem. Biol. 2013, 9, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [Green Version]
- Bez, C.; Covaceuszach, S.; Bertani, I.; Choudhary, K.S.; Venturi, V. LuxR Solos from environmental fluorescent Pseudomonads. mSphere 2021, 6, e01322-20. [Google Scholar] [CrossRef]
- Dominelli, N.; Jäger, H.Y.; Langer, A.; Brachmann, A.; Heermann, R. High-throughput sequencing analysis reveals genomic similarity in phenotypic heterogeneous Photorhabdus luminescens cell populations. Ann. Microbiol. 2022, 72, 20. [Google Scholar] [CrossRef]
- Eckstein, S.; Dominelli, N.; Brachmann, A.; Heermann, R. Phenotypic heterogeneity of the insect pathogen Photorhabdus luminescens: Insights into the fate of secondary cells. Appl. Environ. Microbiol. 2019, 85, e01910-19. [Google Scholar] [CrossRef]
- Regaiolo, A.; Dominelli, N.; Andresen, K.; Heermann, R. The biocontrol agent and insect pathogen Photorhabdus luminescens interacts with plant roots. Appl. Environ. Microbiol 2020, 86, e00891-20. [Google Scholar] [CrossRef] [PubMed]
- Dominelli, N.; Platz, F.; Heermann, R. The insect pathogen Photorhabdus luminescens protects plants from phytopathogenic Fusarium graminearum via chitin degradation. Appl. Environ. Microbiol. 2022, 88, e00645-22. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic. Acids. Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guex, N.; Peitsch, M.C.; Schwede, T. Automated comparative protein structure modeling with SWISS-MODEL and Swiss-PdbViewer: A historical perspective. Electrophoresis 2009, 30, S162–S173. [Google Scholar] [CrossRef]
- Bertoni, M.; Kiefer, F.; Biasini, M.; Bordoli, L.; Schwede, T. modeling protein quaternary structure of homo- and hetero-oligomers beyond binary interactions by homology. Sci. Rep. 2017, 7, 10480. [Google Scholar] [CrossRef] [Green Version]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.P.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL repository—New features and functionality. Nucleic. Acids. Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic. Acids. Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinf. 2020, 36, 1765–1771. [Google Scholar] [CrossRef]
- Zamora-Lagos, M.-A.; Eckstein, S.; Langer, A.; Gazanis, A.; Pfeiffer, F.; Habermann, B.; Heermann, R. Phenotypic and genomic comparison of Photorhabdus luminescens subsp. laumondii TT01 and a widely used rifampicin-resistant Photorhabdus luminescens laboratory strain. BMC Genom. 2018, 19, 854. [Google Scholar] [CrossRef] [Green Version]
- Lassak, J.; Henche, A.-L.; Binnenkade, L.; Thormann, K.M. ArcS, the cognate sensor kinase in an atypical Arc system of Shewanella oneidensis MR-1. Appl. Environ. Microbiol. 2010, 76, 3263–3274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoma, S.; Schobert, M. An improved Escherichia coli donor strain for diparental mating. FEMS Microbiol. Lett. 2009, 294, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, G.D.; Simpson, W.A.; Younger, J.J.; Baddour, L.M.; Barrett, F.F.; Melton, D.M.; Beachey, E.H. Adherence of coagulase-negative Staphylococci to plastic tissue culture plates: A quantitative model for the adherence of Staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A.; Kolter, R. Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signalling pathways: A genetic analysis. Mol. Microbiol. 1998, 28, 449–461. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. JoVE 2011, 47, 2437. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Niesen, F.H.; Berglund, H.; Vedadi, M. The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat. Protoc. 2007, 2, 2212–2221. [Google Scholar] [CrossRef]
- Alexander, C.G.; Wanner, R.; Johnson, C.M.; Breitsprecher, D.; Winter, G.; Duhr, S.; Baaske, P.; Ferguson, N. Novel microscale approaches for easy, rapid determination of protein stability in academic and commercial settings. Biochem. Biophys. Acta Proteins. Proteom. 2014, 1844, 2241–2250. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic. Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Simon, P. Q-Gene: Processing quantitative real-time RT–PCR data. Bioinformatics 2003, 19, 1439–1440. [Google Scholar] [CrossRef] [Green Version]
- Ahmer, B.M.M. Cell-to-cell signalling in Escherichia coli and Salmonella enterica: Quorum sensing in E. coli and Salmonella. Mol. Microbiol. 2004, 52, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Covaceuszach, S.; Degrassi, G.; Venturi, V.; Lamba, D. Structural insights into a novel interkingdom signaling circuit by cartography of the ligand-binding sites of the homologous quorum sensing LuxR-family. IJMS 2013, 14, 20578–20596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, J.F.; Venturi, V. A novel widespread interkingdom signaling circuit. Trends Plant Sci. 2013, 18, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, B.G.; Mevers, E.; Schaefer, A.L.; Pelletier, D.A.; Harwood, C.S.; Clardy, J.; Greenberg, E.P. A Plant-responsive bacterial-signaling system senses an ethanolamine derivative. Proc. Natl. Acad. Sci. USA 2018, 115, 9785–9790. [Google Scholar] [CrossRef] [Green Version]
- Flavier, A.B.; Ganova-Raeva, L.M.; Schell, M.A.; Denny, T.P. Hierarchical autoinduction in Ralstonia solanacearum: Control of acyl-homoserine lactone production by a novel autoregulatory system responsive to 3-hydroxypalmitic acid methyl ester. J. Bacteriol. 1997, 179, 7089–7097. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; Babujee, L.; Jacobs, J.M.; Allen, C. Comparative transcriptome analysis reveals cool virulence factors of Ralstonia solanacearum race 3 biovar 2. PLoS ONE 2015, 10, e0139090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, L.C.M.; Ferreira, R.B.R.; Lostroh, C.P.; Greenberg, E.P. A mutational analysis defines Vibrio fischeri LuxR binding sites. J. Bacteriol. 2008, 190, 4392–4397. [Google Scholar] [CrossRef] [Green Version]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Mattick, J.S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 2002, 56, 289–314. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Tans-Kersten, J.; Huang, H.; Allen, C. Ralstonia solanacearum needs motility for invasive virulence on tomato. J. Bacteriol. 2001, 183, 3597–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corral, J.; Sebastià, P.; Coll, N.S.; Barbé, J.; Aranda, J.; Valls, M. Twitching and swimming motility play a role in Ralstonia solanacearum pathogenicity. mSphere 2020, 5. [Google Scholar] [CrossRef] [Green Version]
- Anetzberger, C.; Pirch, T.; Jung, K. Heterogeneity in quorum sensing-regulated bioluminescence of Vibrio harveyi. Mol. Microbiol. 2009, 73, 267–277. [Google Scholar] [CrossRef]
- Yang, Q.; Defoirdt, T. Quorum sensing positively regulates flagellar motility in pathogenic Vibrio harveyi: QS regulation of motility in Vibrio harveyi. Environ. Microbiol. 2015, 17, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Ahmer, B.M.M.; van Reeuwijk, J.; Timmers, C.D.; Valentine, P.J.; Heffron, F. Salmonella typhimurium encodes an SdiA homolog, a putative quorum sensor of the LuxR family, that regulates genes on the virulence plasmid. J. Bacteriol. 1998, 180, 1185–1193. [Google Scholar] [CrossRef] [Green Version]
- Kanamaru, K.; Kanamaru, K.; Tatsuno, I.; Tobe, T.; Sasakawa, C. SdiA, an Escherichia coli homologue of quorum-sensing regulators, controls the expression of virulence factors in enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 2000, 38, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.M.; Ferreira, R.B.R.; Buckner, M.M.C.; Finlay, B.B. Quorum sensing in bacterial virulence. Microbiol 2010, 156, 2271–2282. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.K.; Bearson, S.M.D.; Bearson, B.L. Evaluation of the effects of SdiA, a LuxR Homologue, on adherence and motility of Escherichia coli O157: H7. Microbiol 2010, 156, 1303–1312. [Google Scholar] [CrossRef] [Green Version]
- Tavio, M.M.; Aquili, V.D.; Poveda, J.B.; Antunes, N.T.; Sanchez-Cespedes, J.; Vila, J. Quorum-sensing regulator sdiA and marA overexpression is involved in in vitro-selected multidrug resistance of Escherichia coli. J. Antimicrob. Chemother. 2010, 65, 1178–1186. [Google Scholar] [CrossRef]
- Culler, H.; Couto, S.; Higa, J.; Ruiz, R.; Yang, M.; Bueris, V.; Franzolin, M.; Sircili, M. Role of SdiA on biofilm formation by atypical enteropathogenic Escherichia coli. Genes 2018, 9, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, A.; Ahmer, B.M.M. Effect of SdiA on biosensors of N-acylhomoserine lactones. J. Bacteriol. 2005, 187, 5054–5058. [Google Scholar] [CrossRef] [Green Version]
- Dyszel, J.L.; Soares, J.A.; Swearingen, M.C.; Lindsay, A.; Smith, J.N.; Ahmer, B.M.M. E. coli K-12 and EHEC genes regulated by SdiA. PLoS ONE 2010, 5, e8946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, D.T.; Terekhova, D.A.; Liou, L.; Hovde, C.J.; Sahl, J.W.; Patankar, A.V.; Gonzalez, J.E.; Edrington, T.S.; Rasko, D.A.; Sperandio, V. Chemical sensing in mammalian host-bacterial commensal associations. PNAS 2010, 107, 9831–9836. [Google Scholar] [CrossRef] [Green Version]
- Shimada, T.; Shimada, K.; Matsui, M.; Kitai, Y.; Igarashi, J.; Suga, H.; Ishihama, A. Roles of cell division control factor SdiA: Recognition of quorum sensing signals and modulation of transcription regulation targets. Genes Cells 2014, 19, 405–418. [Google Scholar] [CrossRef]
- Nguyen, Y.; Nguyen, N.X.; Rogers, J.L.; Liao, J.; MacMillan, J.B.; Jiang, Y.; Sperandio, V. Structural and mechanistic roles of novel chemical ligands on the SdiA quorum-sensing transcription regulator. mBio 2015, 6, e02429-14. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Guan, W.; Huang, Q.; Yang, Y.; Yan, W.; Sun, B.; Zhao, T. Quorum-sensing contributes to virulence, twitching motility, seed attachment and biofilm formation in the wild type strain Aac-5 of Acidovorax citrulli. Microb. Pathogen. 2016, 100, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, T.; Gomes, A.É.I.; Siqueira, N.M.G.; Assoni, L.; Darrieux, M.; Venter, H.; Ferraz, L.F.C. SdiA, a quorum-sensing regulator, suppresses fimbriae expression, biofilm formation, and quorum-sensing signaling molecules production in Klebsiella pneumoniae. Front Microbiol. 2021, 12, 597735. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-S.; Winans, S.C. LuxR-type quorum-sensing regulators that are detached from common scents: LuxR-type apoproteins. Mol. Microbiol. 2010, 77, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Jayaraman, A.; Wood, T.K. Indole is an inter-species biofilm signal mediated by SdiA. BMC Microbiol. 2007, 7, 42. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Maeda, T.; Hong, S.H.; Wood, T.K. Reconfiguring the quorum-sensing regulator SdiA of Escherichia coli to control biofilm formation via indole and N-acylhomoserine lactones. Appl. Environ. Microbiol. 2009, 75, 1703–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Martinez-Yamout, M.A.; Dickerson, T.J.; Brogan, A.P.; Wright, P.E.; Dyson, H.J. Structure of the Escherichia coli quorum sensing protein SdiA: Activation of the folding switch by acyl homoserine lactones. J. Mol. Biol. 2006, 355, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Teplitski, M.; Robinson, J.B.; Bauer, W.D. Plants secrete substances that mimic bacterial N-acyl homoserine lactone signal activities and affect population density-dependent behaviors in associated bacteria. Mol. Plant. Microbe Interact. 2000, 13, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Castang, S.; Reverchon, S.; Gouet, P.; Nasser, W. Direct evidence for the modulation of the activity of the Erwinia chrysanthemi quorum-sensing regulator ExpR by acylhomoserine lactone pheromone. J. Biol. Chem. 2006, 281, 29972–29987. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Dixit, V.; Leigh, N.; Chatterjee, A.K. ExpR, a LuxR homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol. 2005, 187, 4792–4803. [Google Scholar] [CrossRef] [Green Version]
- Minogue, T.D.; Trebra, M.W.; Bernhard, F.; von Bodman, S.B. The autoregulatory role of EsaR, a quorum-sensing regulator in Pantoea stewartii ssp. stewartii: Evidence for a repressor function: Autoregulatory role of EsaR. Mol. Microbiol. 2002, 44, 1625–1635. [Google Scholar] [CrossRef] [Green Version]
- Minogue, T.D.; Carlier, A.L.; Koutsoudis, M.D.; von Bodman, S.B. The cell density-dependent expression of stewartan exopolysaccharide in Pantoea stewartii ssp. stewartii is a function of EsaR-mediated repression of the rcsA gene: Quorum sensing control of stewartan EPS. Mol. Microbiol. 2005, 56, 189–203. [Google Scholar] [CrossRef]
- Devine, J.H.; Shadel, G.S.; Baldwin, T.O. Identification of the operator of the lux regulon from the Vibrio fischeri strain ATCC7744. PNAS 1989, 86, 5688–5692. [Google Scholar] [CrossRef] [Green Version]
- Stevens, A.M.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Essential elements for activation of the luminescence genes. J. Bacteriol. 1997, 179, 557–562. [Google Scholar] [CrossRef] [Green Version]
- Egland, K.A.; Greenberg, E.P. Quorum sensing in Vibrio fischeri: Elements of the luxI promoter. Mol. Microbiol. 1999, 31, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Shadel, G.S.; Baldwin, T.O. The Vibrio fischeri LuxR protein is capable of bidirectional stimulation of transcription and both positive and negative regulation of the luxR gene. J. Bacteriol. 1991, 173, 568–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence 5′-> 3′ |
|---|---|
| [Btn]-PsdiA fwd | [Btn]-GATTATTAGGATTTCAATCCTATTGATAT |
| PsdiA rev | TCAATGTCCTCTTGAAAATTAAG |
| [Btn]-PaidA fwd | [Btn]-GACACCTCTTTACATATTTAAACTATT |
| PaidA rev | CTATATGAAGCAATACCTAATAAATATATG |
| [Btn]-PfliE fwd | [Btn]-GTCATTATTCGCTGTTCACTC |
| PfliE rev | AAAAACCTCGTGTTAAACCAC |
| aidA-qPCR-fwd | TCCAACAGTTATCCGTCAGC |
| aidA-qPCR-rev | GCCCTCCATCTAATATTCGCA |
| fliE-qPCR-fwd | GTGCTGCAACTGATGCAAG |
| fliE-qPCR-rev | GAGCTCGTTTTGTGGCATTC |
| rpoD-qPCR-fwd | CGGAAGATATCGTCGATTCCGA |
| rpoD-qPCR-rev | TGTCGTTAGCGGTTTCTGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dominelli, N.; Regaiolo, A.; Willy, L.; Heermann, R. Interkingdom Signaling of the Insect Pathogen Photorhabdus luminescens with Plants Via the LuxR solo SdiA. Microorganisms 2023, 11, 890. https://doi.org/10.3390/microorganisms11040890
Dominelli N, Regaiolo A, Willy L, Heermann R. Interkingdom Signaling of the Insect Pathogen Photorhabdus luminescens with Plants Via the LuxR solo SdiA. Microorganisms. 2023; 11(4):890. https://doi.org/10.3390/microorganisms11040890
Chicago/Turabian StyleDominelli, Nazzareno, Alice Regaiolo, Leon Willy, and Ralf Heermann. 2023. "Interkingdom Signaling of the Insect Pathogen Photorhabdus luminescens with Plants Via the LuxR solo SdiA" Microorganisms 11, no. 4: 890. https://doi.org/10.3390/microorganisms11040890