
Biofilm Formation by Mutant Strains of Bacilli under Different Stress Conditions
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
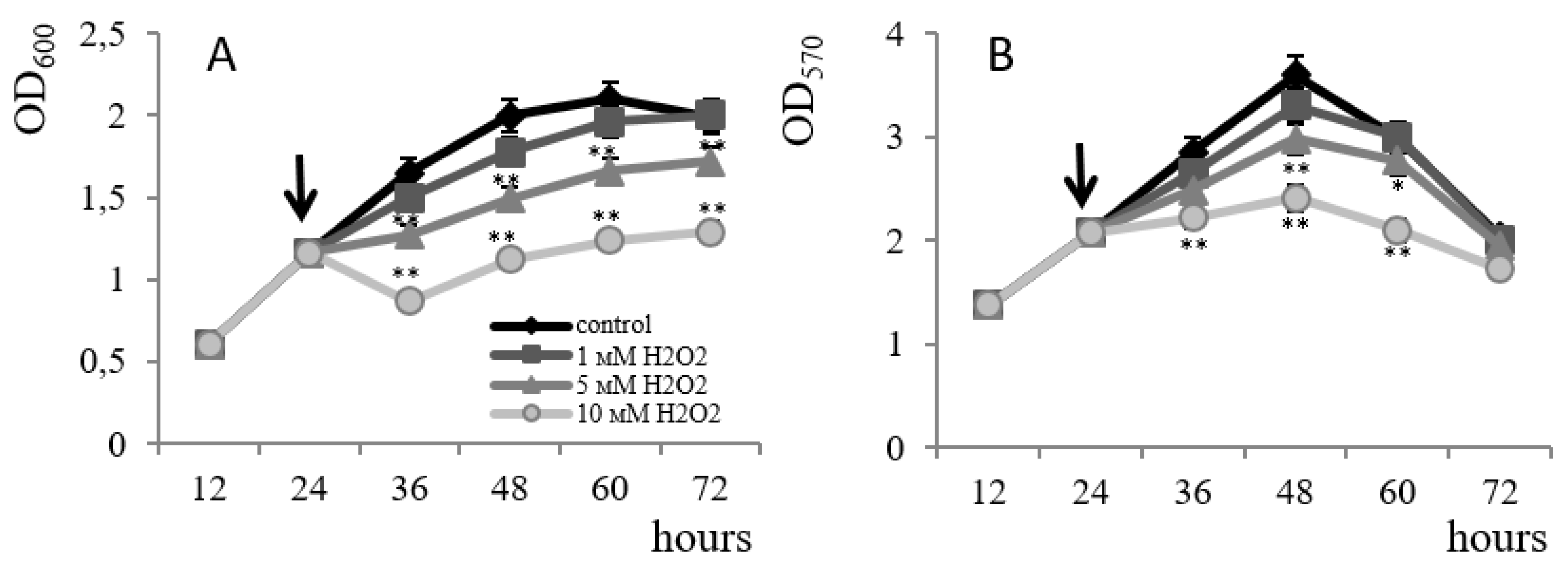
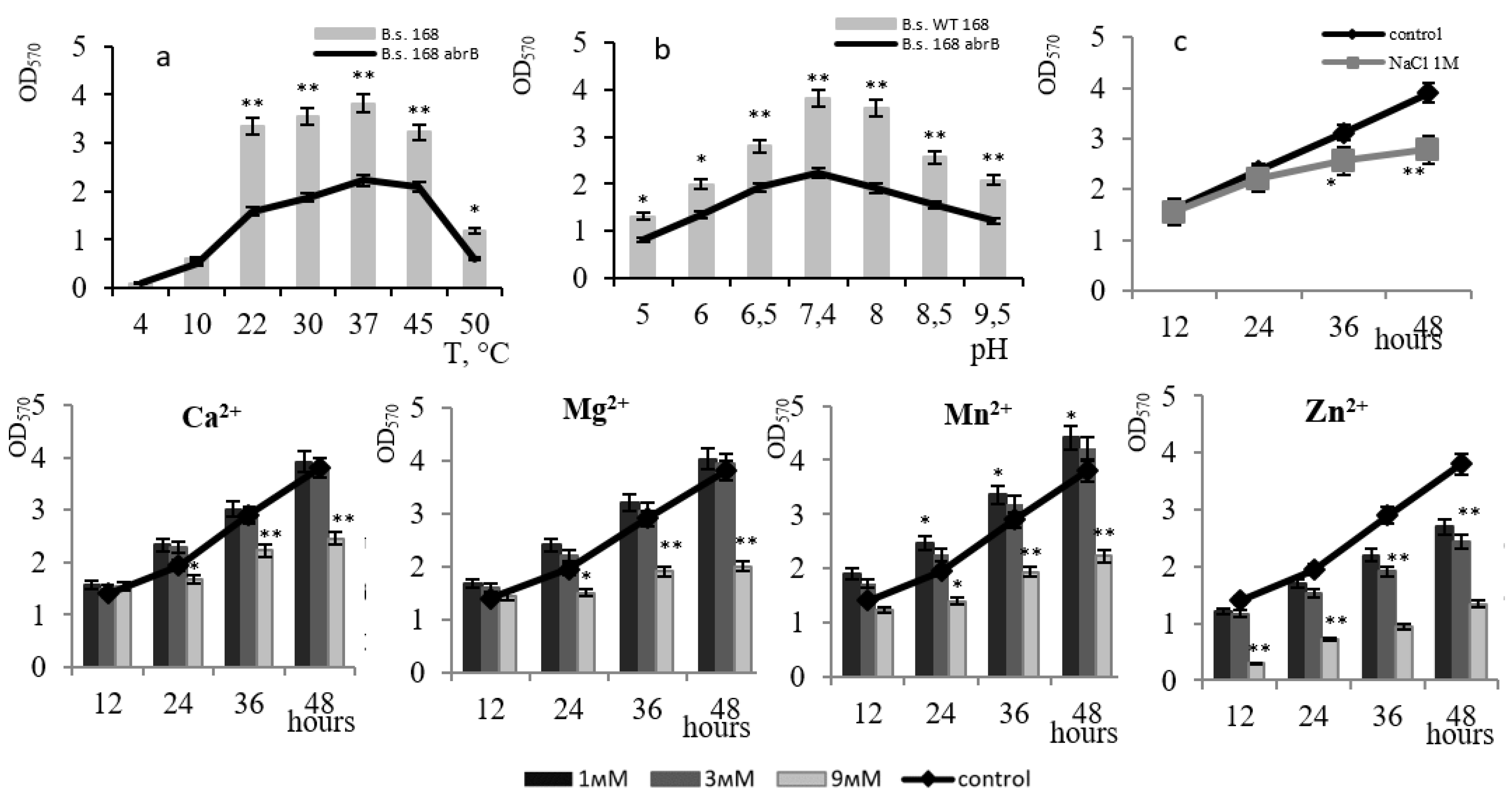
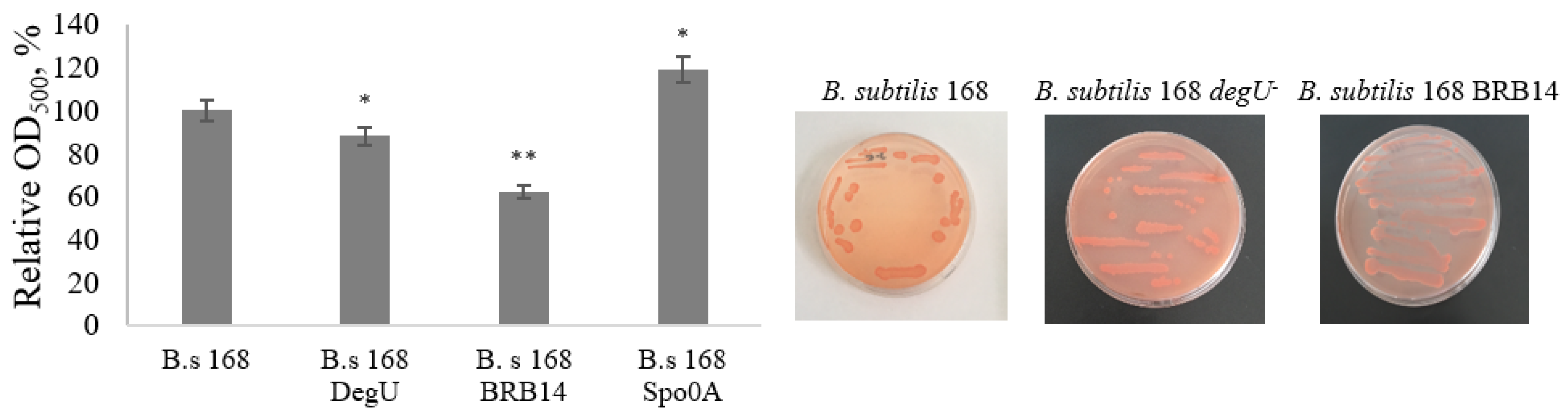
3. Results
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, R.; Feng, H.; Xu, Z.; Zhang, N.; Liu, Y.; Shao, J.; Shen, Q.; Zhang, R. Identification of adhesins in plant beneficial rhizobacteria Bacillus velezensis SQR9 and their effect on root colonization. Mol. Plant-Microbe Interact. 2022, 35, 64–72. [Google Scholar] [CrossRef]
- Feng, H.; Fu, R.; Hou, H.; Lv, Y.; Zhang, N.; Liu, Y.; Xu, Z.; Miao, Y.; Krell, T.; Shen, Q.; et al. Chemotaxis of beneficial rhizobacteria to root exudates: The first step towards root–microbe rhizosphere interactions. Int. J. Mol. Sci. 2021, 22, 6655. [Google Scholar] [CrossRef]
- Allard-Massicotte, R.; Tessier, L.; Lécuyer, F.; Lakshmanan, V.; Lucier, J.-F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.V.; Beauregard, P.B. Bacillus subtilis early colonization of Arabidopsis thaliana roots involve multiple chemotaxis receptors. MBio 2016, 7, e01664-16. [Google Scholar] [CrossRef] [Green Version]
- Hashem, A.; Tabassum, B.; Abd Allah, E.F. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef]
- Arnaouteli, S.; Bamford, N.C.; Stanley-Wall, N.R.; Kovács, A.T. Bacillus subtilis biofilm formation and social interactions. Nat. Rev. Microbiol. 2021, 19, 600–614. [Google Scholar] [CrossRef]
- Cairns, L.S.; Hobley, L.; Stanley-Wall, N.R. Biofilm formation by Bacillus subtilis: New insights into regulatory strategies and assembly mechanisms. Mol. Microbiol. 2014, 93, 587–598. [Google Scholar] [CrossRef] [Green Version]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Feng, H.; Fu, R.; Zhang, N.; Du, W.; Shen, Q.; Zhang, R. Induced root-secreted D-galactose functions as a chemoattractant and enhances the biofilm formation of Bacillus velezensis SQR9 in an McpA-dependent manner. Appl. Microbiol. Biotechnol. 2020, 104, 785–797. [Google Scholar] [CrossRef]
- Jiao, H.; Xu, W.; Hu, Y.; Tian, R.; Wang, Z. Citric acid in rice root exudates enhanced the colonization and plant growth-promoting ability of Bacillus altitudinis LZP02. Microbiol. Spectr. 2022, 10, e0100222. [Google Scholar] [CrossRef]
- Vora, S.M.; Joshi, P.; Belwalkar, M.; Archana, G. Root exudates influence chemotaxis and colonization of diverse plant growth promoting rhizobacteria in the pigeon pea-maize intercropping system. Rhizosphere 2021, 18, 100331. [Google Scholar] [CrossRef]
- Pandin, C.; Le Coq, D.; Canette, A.; Aymerich, S.; Briandet, R. Should the biofilm mode of life be taken into consideration for microbial biocontrol agents? Microb. Biotechnol. 2017, 10, 719–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahapatra, S.; Yadav, R.; Ramakrishna, W. Bacillus subtilis impact on plant growth, soil health and environment: Dr. Jekyll and Mr. Hyde. J. Appl. Microbiol. 2022, 132, 3543–3562. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, M.; Kagihiro, S.; Haruki, M.; Takano, K.; Branda, S.; Kolter, R.; Kanaya, S. Biofilm formation by a Bacillus subtilis strain that produces γ-polyglutamate. Microbiology 2006, 152, 2801–2807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar] [CrossRef]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. Curr. Protoc. Microbiol. 2005, 1, Unit 1B.1. [Google Scholar] [CrossRef] [Green Version]
- Reichhardt, C.; Jacobson, A.N. Congo Red interactions with curli-producing E. coli and native curli amyloid fibers. PLoS ONE 2015, 10, e0140388. [Google Scholar] [CrossRef] [Green Version]
- Gophna, U.; Barlev, M.; Seijffers, R.; Oelschlager, T.A.; Hacker, J.; Ron, E.Z. Curli fibers mediate internalization of Escherichia coli by eukaryotic cells. Infect. Immun. 2001, 69, 2659–2665. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.R.; Mustafa, A.; Hyder, S.; Valipour, M.; Rizvi, Z.F.; Gondal, A.S.; Yousuf, Z.; Iqbal, R.; Daraz, U. Bacillus spp. as bioagents: Uses and application for sustainable agriculture. Biology 2022, 11, 1763. [Google Scholar] [CrossRef]
- Ahmad, I.; Akhtar, M.J.; Mehmood, S.; Akhter, K.; Tahir, M.; Saeed, M.F.; Hussain, M.B.; Hussain, S. Combined application of compost and Bacillus sp. CIK-512 ameliorated the lead toxicity in radish by regulating the homeostasis of antioxidants and lead. Ecotoxicol. Environ. Saf. 2018, 148, 805–812. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef]
- Khedher, S.B.; Mejdoub-Trabelsi, B.; Tounsi, S. Biological potential of Bacillus subtilis V26 for the control of Fusarium wilt and tuber dry rot on potato caused by Fusarium species and the promotion of plant growth. Biol. Control 2021, 152, 104444. [Google Scholar] [CrossRef]
- Chu, D.; Wang, X.; Fan, J.; Peng, L.; Liang, J.; Zheng, Y.; Wang, R.; You, X.; Jing, C.; Clark, J.; et al. Ulva prolifera polysaccharide enhances the root colonisation by Bacillus amyloliquefaciens strain Cas02. Carbohydr. Polym. 2023, 299, 120171. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yan, H.; Zhang, J.; Gao, Y.; Xu, W.; Shang, J.; Luo, Y. Inhibition of biofilm formation by Cd2+ on Bacillus subtilis 1JN2 depressed its biocontrol efficiency against Ralstonia wilt on tomato. Microbiol. Res. 2018, 215, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Flint, S.; Palmer, J. Magnesium and calcium ions: Roles in bacterial cell attachment and biofilm structure maturation. Biofouling 2019, 35, 959–974. [Google Scholar] [CrossRef]
- Oomes, S.J.; Jonker, M.J.; Wittink, F.R.; Hehenkamp, J.O.; Breit, T.M.; Brul, T.M. The effect of calcium on the transcriptome of sporulating B. subtilis cells. Int. J. Food Microbiol. 2009, 133, 234–242. [Google Scholar] [CrossRef]
- Tian, L.; Chen, X.D.; Yang, Q.P.; Chen, J.C.; Shi, L.; Li, Q. Effect of calcium ions on the evolution of biofouling by Bacillus subtilis in plate heat exchangers simulating the heat pump system used with treated sewage in the 2008 Olympic Village. Colloids Surf. B: Biointerfaces 2012, 94, 309–316. [Google Scholar] [CrossRef]
- Keren-Paz, A.; Maan, H.; Karunker, I.; Olender, T.; Kapishnikov, S.; Dersch, S.; Kartvelishvily, E.; Wolf, S.G.; Gal, A.; Graumann, P.L.; et al. The roles of intracellular and extracellular calcium in Bacillus subtilis biofilms. Iscience 2022, 25, 104308. [Google Scholar] [CrossRef]
- Shemesh, M.; Chai, Y. A combination of glycerol and manganese promotes biofilm formation in Bacillus subtilis via histidine kinase KinD signaling. J. Bacteriol. 2013, 195, 2747–2754. [Google Scholar] [CrossRef] [Green Version]
- Mhatre, E.; Troszok, A.; Gallegos-Monterrosa, R.; Lindstädt, S.; Hölscher, T.; Kuipers, O.P.; Kovács, A.T. The impact of manganese on biofilm development of Bacillus subtilis. Microbiology 2016, 162, 1468–1478. [Google Scholar] [CrossRef]
- Ran, X.; Zhu, Z.; Long, H.; Tian, Q.; You, L.; Wu, X.; Liu, Q.; Huang, S.; Li, S.; Niu, X.; et al. Manganese stress adaptation mechanisms of Bacillus safensis strain ST7 from mine soil. Front. Microbiol. 2021, 12, 758889. [Google Scholar] [CrossRef]
- He, X.; Wang, J.; Abdoli, L.; Li, H. Mg2+/Ca2+ promotes the adhesion of marine bacteria and algae and enhances following biofilm formation in artificial seawater. Colloids Surf. B: Biointerfaces 2016, 146, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kuang, H.; Liu, Y.; Xu, H.; Aguilar, Z.P.; Xiong, Y.; Wei, H. Mechanism of enhanced antibacterial activity of ultra-fine ZnO in phosphate buffer solution with various organic acids. Environ. Pollut. 2016, 218, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Sooch, B.S.; Kauldhar, B.S.; Puri, M. Recent insights into microbial catalases: Isolation, production and purification. Biotechnol. Adv. 2014, 32, 1429–1447. [Google Scholar] [CrossRef]
- Shaeer, A.; Aslam, M.; Rashid, N. Structural and functional analyses of a novel manganese-catalase from Bacillus subtilis R5. Int. J. Biol. Macromol. 2021, 180, 222–233. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Shao, W.; Perego, M.; Hoch, J.A. Multiple histidine kinases regulate entry into stationary phase and sporulation in Bacillus subtilis. Mol. Microbiol. 2000, 38, 535–542. [Google Scholar] [CrossRef]
- Chen, Z.; Srivastava, P.; Zarazúa-Osorio, B.; Marathe, A.; Fujita, M.; Igoshin, O.A. Bacillus subtilis histidine kinase KinC activates biofilm formation by controlling heterogeneity of single-cell responses. MBio 2022, 13, e0169421. [Google Scholar] [CrossRef]
- Grau, R.R.; de Oña, P.; Kunert, M.; Leñini, C.; Gallegos-Monterrosa, R.; Mhatre, E.; Vileta, D.; Donato, V.; Hölscher, T.; Boland, W.; et al. A duo of potassium-responsive histidine kinases governs the multicellular destiny of Bacillus subtilis. MBio 2015, 6, e00581. [Google Scholar] [CrossRef] [Green Version]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, 1621–1630. [Google Scholar] [CrossRef] [Green Version]
- Kolodkin-Gal, I.; Elsholz, A.K.; Muth, C.; Girguis, P.R.; Kolter, R.; Losick, R. Respiration control of multicellularity in Bacillus subtilis by a complex of the cytochrome chain with a membrane-embedded histidine kinase. Genes Dev. 2013, 27, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Rey, M.W.; Ramaiya, P.; Nelson, B.A.; Brody-Karpin, B.A.; Zaretsky, E.J.; Tang, M.; de Leon, A.L.; Xiang, H.; Gusti, V.; Clausen, I.G.; et al. Complete genome sequence of the industrial bacterium Bacillus licheniformis and comparisons with closely related Bacillus species. Genome Biol. 2004, 5, r77. [Google Scholar] [CrossRef] [Green Version]
- Cardoso, P.F.; Perchat, S.; Vilas-Boas, L.A.; Lereclus, D.; Vilas-Bôas, G.T. Diversity of the Rap-Phr quorum-sensing systems in the Bacillus cereus group. Curr. Genet. 2019, 65, 1367–1381. [Google Scholar] [CrossRef] [PubMed]
- Boguslawski, K.M.; Hill, P.A.; Griffith, K.L. Novel mechanisms of controlling the activities of the transcription factors Spo0A and ComA by the plasmid-encoded quorum sensing regulators Rap60-Phr60 in Bacillus subtilis. Mol. Microbiol. 2015, 96, 325–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallegos-Monterrosa, R.; Christensen, M.N.; Barchewitz, T.; Koppenhöfer, S.; Priyadarshini, B.; Bálint, B.; Maróti, G.; Kempen, P.J.; Dragoš, A.; Kovács, Á.T. Impact of Rap-Phr system abundance on adaptation of Bacillus subtilis. Commun. Biol. 2021, 4, 468. [Google Scholar] [CrossRef] [PubMed]
- Arbour, C.A.; Nagar, R.; Bernstein, H.M.; Ghosh, S.; Al-Sammarraie, Y.; Dorfmueller, H.C.; Ferguson, M.A.J.; Stanley-Wall, N.R.; Imperiali, B. Preprint defining early steps in B. subtilis biofilm biosynthesis. bioRxiv 2023. [Google Scholar] [CrossRef]
- Musik, J.E.; Zalucki, Y.M.; Day, C.J.; Jennings, M.P. Expression of the Bacillus subtilis TasA signal peptide leads to cell death in Escherichia coli due to inefficient cleavage by LepB. Biochim. et Biophys. Acta (BBA)-Biomembr. 2021, 1863, 183768. [Google Scholar] [CrossRef]
- Romero, D.; Vlamakis, H.; Losick, R.; Kolter, R. Functional analysis of the accessory protein TapA in Bacillus subtilis amyloid fiber assembly. J. Bacteriol. 2014, 196, 1505–1513. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Mutation Description | Source |
|---|---|---|
| Bacillus subtilis WT 168 | Natural isolate (wild-type) | Proffessor J. Stuelke, University of Göttingen, Germany |
| Protease-deficient strains | ||
| Bacillus subtilis BG20-36 | ΔnprE–522; Δapr–684 | Eugenio Ferrarri, Genencor Int. Inc., Rochester, NY, USA |
| Bacillus subtilis BRB08 | ΔtrpC2, ΔnprB, ΔaprE, Δepr, Δbpr, ΔnprE, Δmpr, Δvpr, ΔwprA | Cobra Biologics, Keele, UK |
| Bacillus subtilis BRB14 | ΔtrpC2, ΔnprB, ΔaprE, Δepr, Δbpr, ΔnprE, Δmpr, Δvpr, ΔwprA, ΔhtrA, ΔhtrB | Cobra Biologics, Keele, UK |
| Strains with regulatory mutations | ||
| Bacillus subtilis 168 abrB::kan (amyE::pAT606) | ΔabrB (Kan) | Dr. Prof. T. Masher Ludwig-Maximillian’s University, Munich, Germany |
| Bacillus subtilis 168 degU::kan (amyE::pAT606) | ΔdegU (Kan) | Dr. Prof. T. Masher Ludwig-Maximillian’s University, Munich, Germany |
| Bacillus subtilis 168 spo0A::tet (amyE::pAT612) | Δspo0A (Tet) | Dr. Prof. T. Masher Ludwig-Maximillian’s University, Munich, Germany |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharipova, M.; Rudakova, N.; Mardanova, A.; Evtugyn, V.; Akosah, Y.; Danilova, I.; Suleimanova, A. Biofilm Formation by Mutant Strains of Bacilli under Different Stress Conditions. Microorganisms 2023, 11, 1486. https://doi.org/10.3390/microorganisms11061486
Sharipova M, Rudakova N, Mardanova A, Evtugyn V, Akosah Y, Danilova I, Suleimanova A. Biofilm Formation by Mutant Strains of Bacilli under Different Stress Conditions. Microorganisms. 2023; 11(6):1486. https://doi.org/10.3390/microorganisms11061486
Chicago/Turabian StyleSharipova, Margarita, Natalia Rudakova, Ayslu Mardanova, Vladimir Evtugyn, Yaw Akosah, Iuliia Danilova, and Aliya Suleimanova. 2023. "Biofilm Formation by Mutant Strains of Bacilli under Different Stress Conditions" Microorganisms 11, no. 6: 1486. https://doi.org/10.3390/microorganisms11061486