
Whole Genome Sequence Analysis of Cupriavidus necator C39, a Multiple Heavy Metal(loid) and Antibiotic Resistant Bacterium Isolated from a Gold/Copper Mine
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation of Bacterial Strain C39
2.3. Determination of Minimum Inhibitory Concentration (MIC) of Heavy Metals and Antibiotics
2.4. Determination of Growth on Aromatic Compounds
2.5. Whole Genome Sequencing
2.6. Genome Assembly and Annotation
2.7. Phylogenetic Analysis
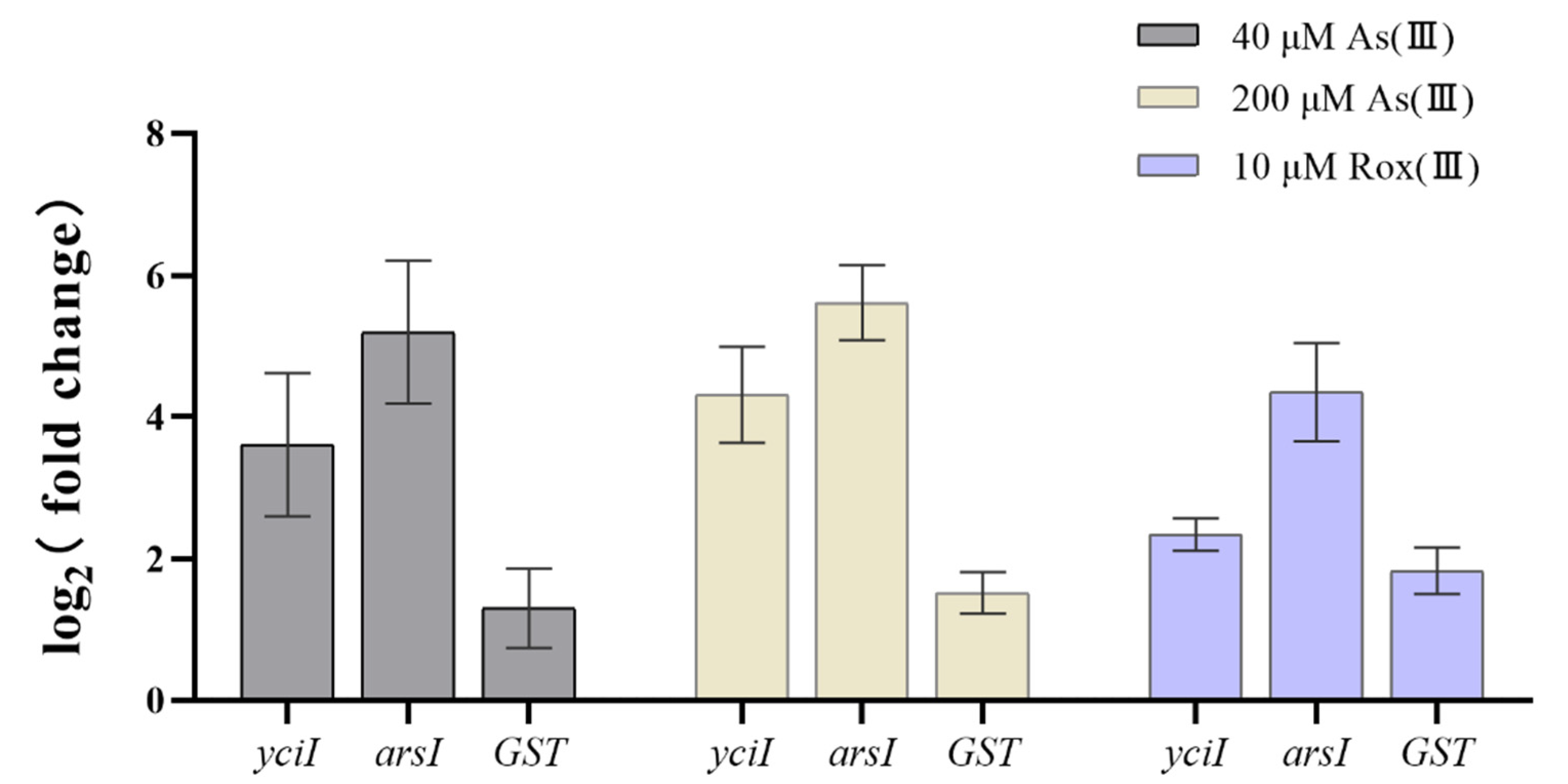
2.8. RT-qPCR Analysis
3. Results and Discussion
3.1. Minimum Inhibitory Concentration (MIC) Results of Cupriavidus Necator C39
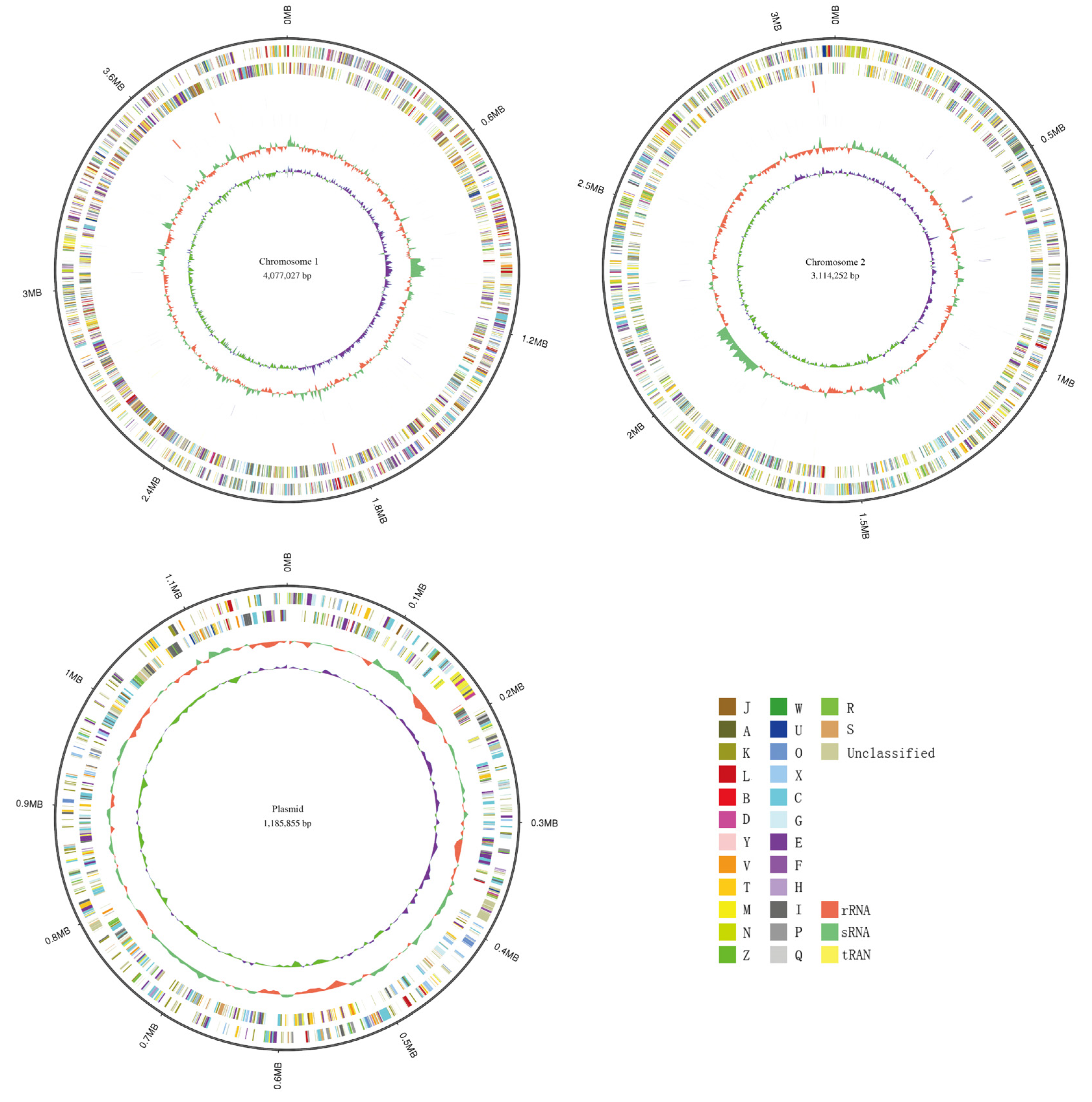
3.2. Genomic Assembly and Features
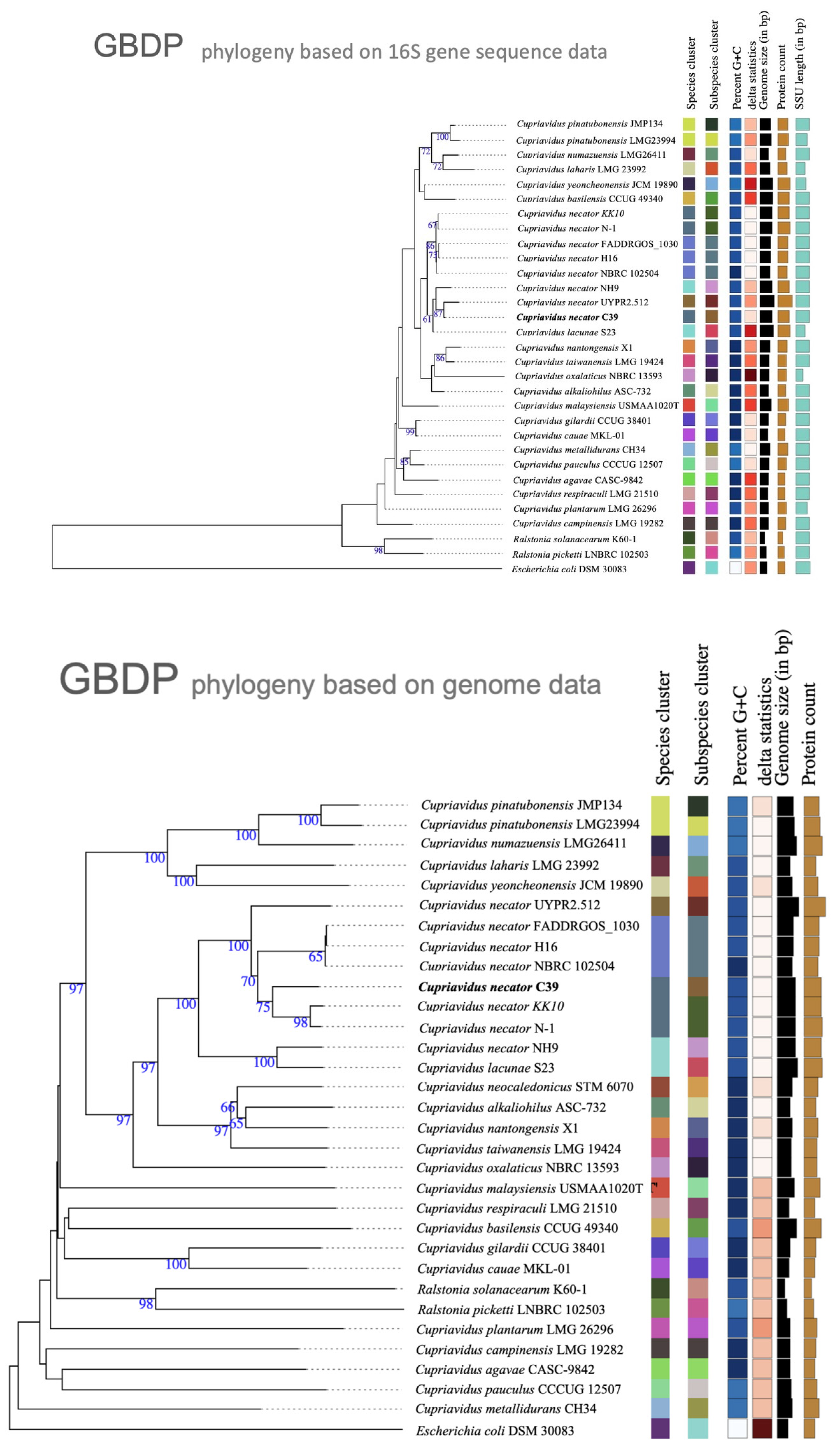
3.3. Phylogenetic Characterization
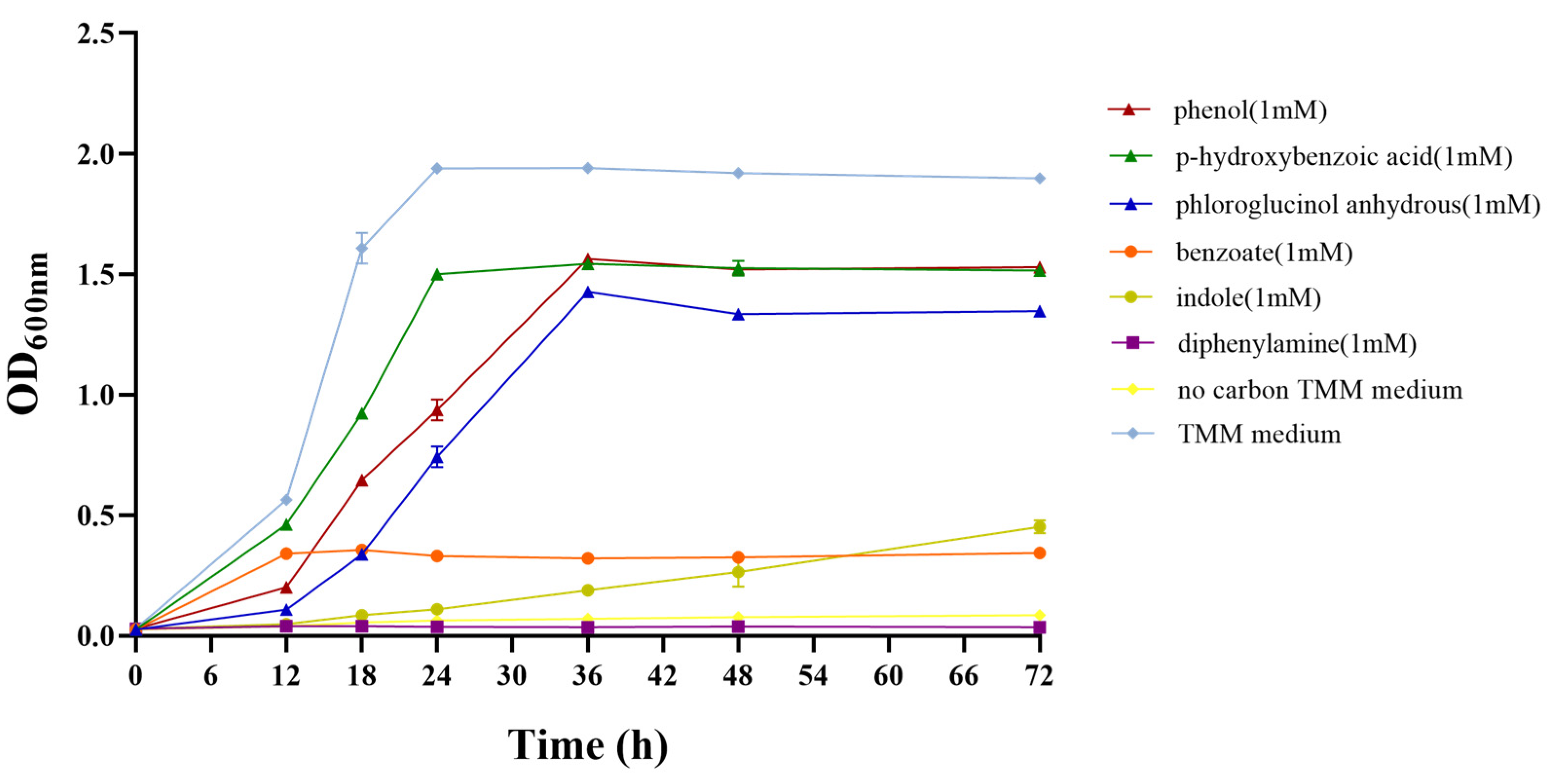
3.4. Degradation of Aromatic Compounds
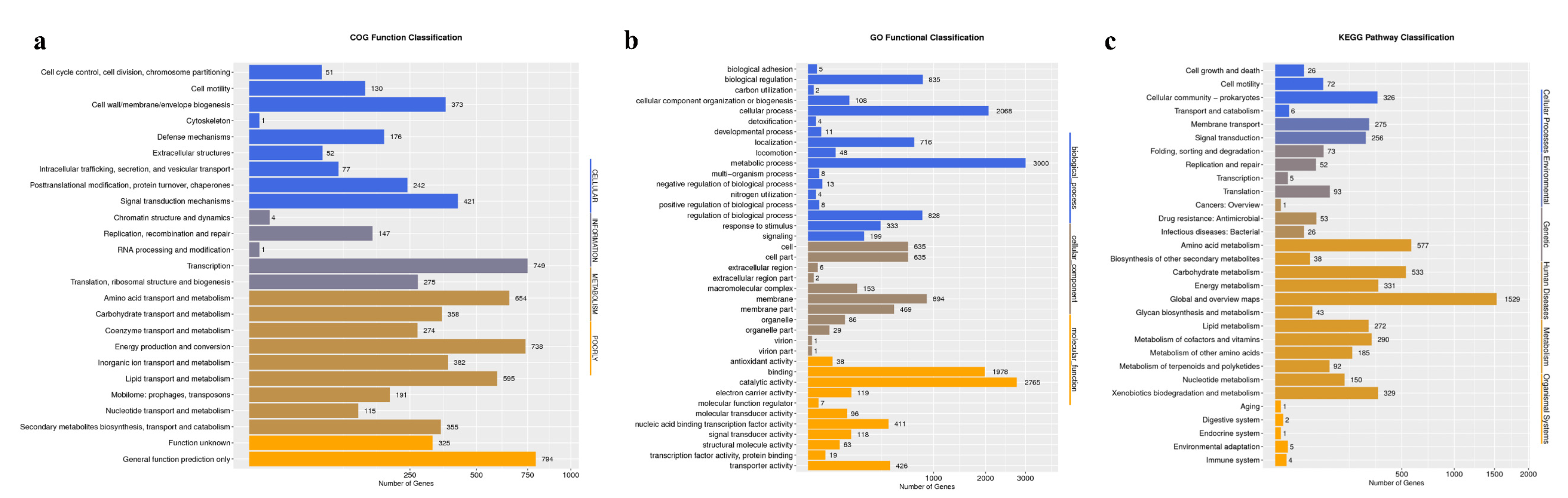
3.5. Functional Annotations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lee, S.; Khanal, A.; Cho, A.H.; Lee, H.; Kang, M.S.; Unno, T.; Hur, H.G.; Lee, J.H. Cupriavidus sp. strain Ni-2 resistant to high concentration of nickel and its genes responsible for the tolerance by genome comparison. Arch. Microbiol. 2019, 201, 1323–1331. [Google Scholar] [CrossRef]
- Perez-Pantoja, D.; De la Lglesia, R.; Pieper, D.H.; Gonzalez, B. Metabolic reconstruction of aromatic compounds degradation from the genome of the amazing pollutant-degrading bacterium Cupriavidus necator JMP134. Fems Microbiol. Rev. 2008, 32, 736–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sohn, Y.J.; Son, J.; Jo, S.Y.; Park, S.Y.; Yoo, J.I.; Baritugo, K.A.; Na, J.G.; Choi, J.I.; Kim, H.T.; Joo, J.C.; et al. Chemoautotroph Cupriavidus necator as a potential game-changer for global warming and plastic waste problem: A review. Bioresour. Technol. 2021, 340, e125693. [Google Scholar] [CrossRef] [PubMed]
- Makkar, N.S.; Casida, L.E. Cupriavidus necator gen. nov., sp. nov.; a nonobligate bacterial predator of bacteria in soil. Int. J. Syst. Evol. Microbiol. 1987, 37, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Budde, C.F.; Mahan, A.E.; Lu, J.N.; Rha, C.; Sinskey, A.J. Roles of multiple acetoacetyl coenzyme a reductases in polyhydroxybutyrate biosynthesis in Ralstonia eutropha H16. J. Bacteriol. 2010, 192, 5319–5328. [Google Scholar] [CrossRef] [Green Version]
- Brigham, C.J.; Gai, C.S.; Lu, J.; Speth, D.R.; Worden, R.M.; Sinskey, A.J. Engineering Ralstonia eutropha for production of isobutanol from CO2, H2, and O2. In Advanced Biofuels and Bioproducts; Lee, J., Ed.; Springer: New York, NY, USA, 2013; Volume 3, pp. 1065–1090. [Google Scholar] [CrossRef]
- Lee, H.M.; Jeon, B.Y.; Oh, M.K. Microbial production of ethanol from acetate by engineered Ralstonia eutropha. Biotechnol. Bioprocess Eng. 2016, 21, 402–407. [Google Scholar] [CrossRef]
- Lu, J.N.; Brigham, C.J.; Gai, C.S.; Sinskey, A.J. Studies on the production of branched-chain alcohols in engineered Ralstonia eutropha. Appl. Microbiol. Biotechnol. 2012, 96, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Grousseau, E.; Lu, J.N.; Gorret, N.; Guillouet, S.E.; Sinskey, A.J. Isopropanol production with engineered Cupriavidus necator as bioproduction platform. Appl. Microbiol. Biotechnol. 2014, 98, 4277–4290. [Google Scholar] [CrossRef] [Green Version]
- Muller, J.; MacEachran, D.; Burd, H.; Sathitsuksanoh, N.; Bi, C.H.; Yeh, Y.C.; Lee, T.S.; Hillson, N.J.; Chhabra, S.R.; Singer, S.W.; et al. Engineering of Ralstonia eutropha H16 for autotrophic and heterotrophic production of methyl ketones. Appl. Environ. Microbiol. 2013, 79, 4433–4439. [Google Scholar] [CrossRef] [Green Version]
- Przybylski, D.; Rohwerder, T.; Dilssner, C.; Maskow, T.; Harms, H.; Muller, R.H. Exploiting mixtures of H2, CO2, and O2 for improved production of methacrylate precursor 2-hydroxyisobutyric acid by engineered Cupriavidus necator strains. Appl. Microbiol. Biotechnol. 2015, 99, 2131–2145. [Google Scholar] [CrossRef]
- Park, J.M.; Kim, T.Y.; Lee, S.Y. Genome-scale reconstruction and in silico analysis of the Ralstonia eutropha H16 for polyhydroxyalkanoate synthesis, lithoautotrophic growth, and 2-methyl citric acid production. BMC Syst. Biol. 2011, 5, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poehlein, A.; Kusian, B.; Friedrich, B.; Daniel, R.; Bowien, B. Complete genome sequence of the type strain Cupriavidus necator N-1. J. Bacteriol. 2011, 193, 5017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lykidis, A.; Perez-Pantoja, D.; Ledger, T.; Mavromatis, K.; Anderson, I.J.; Ivanova, N.N.; Hooper, S.D.; Lapidus, A.; Lucas, S.; Gonzalez, B.; et al. The complete multipartite genome sequence of Cupriavidus necator JMP134, a versatile pollutant degrader. PLoS ONE 2010, 5, e9729. [Google Scholar] [CrossRef]
- Xin, Y.F.; Gao, R.; Cui, F.F.; Lu, C.J.; Liu, H.L.; Liu, H.W.; Xia, Y.Z.; Xun, L.Y. The heterotrophic bacterium Cupriavidus pinatubonensis JMP134 oxidizes sulfide to sulfate with thiosulfate as a key intermediate. Appl. Environ. Microbiol. 2020, 86, e01835-e20. [Google Scholar] [CrossRef] [PubMed]
- Little, G.T.; Ehsaan, M.; Arenas-Lopez, C.; Jawed, K.; Winzer, K.; Kovacs, K.; Minton, N.P. Complete genome sequence of Cupriavidus necator H16 (DSM 428). Microbiol. Resour. Ann. 2019, 8, e00814–e00819. [Google Scholar] [CrossRef] [Green Version]
- Mori, J.F.; Nagai, M.; Kanaly, R.A. Complete genome sequence of Cupriavidus necator KK10, an azaarene-degrading and polyhydroxyalkanoate-producing soil bacterium. Microbiol. Resour. Ann. 2021, 10, e0042321. [Google Scholar] [CrossRef]
- Moriuchi, R.; Dohra, H.; Kanesaki, Y.; Ogawa, N. Complete genome sequence of 3-chlorobenzoate-degrading bacterium Cupriavidus necator NH9 and reclassification of the strains of the genera Cupriavidus and Ralstonia based on phylogenetic and whole-genome sequence analyses. Front. Microbiol. 2019, 10, 133. [Google Scholar] [CrossRef]
- Rodriguez-Esperon, M.C.; Eastman, G.; Sandes, L.; Garabato, F.; Eastman, I.; Iriarte, A.; Fabiano, E.; Sotelo-Silveira, J.R.; Platero, R. Genomics and transcriptomics insights into luteolin effects on the beta-rhizobial strain Cupriavidus necator UYPR2.512. Environ. Microbiol. 2022, 24, 240–264. [Google Scholar] [CrossRef]
- Yonezuka, K.; Shimodaira, J.; Tabata, M.; Nagase, S.; Kasai, D.; Hosoyama, A.; Yamazoe, A.; Fujita, N.; Fukuda, M. Draft genome sequence of a chlorinated-ethene degrader, Cupriavidus necator strain PHE3-6 (NBRC 110655). Genome Announc. 2016, 4, e01743-e15. [Google Scholar] [CrossRef] [Green Version]
- Margaryan, A. Diversity and application of heavy-metal resistant microbes. In Microbes in Microbial Communities; Singh, R.P., Manchanda, G., Bhattacharjee, K., Panosyan, H., Eds.; Springer: Singapore, 2021; pp. 153–174. [Google Scholar] [CrossRef]
- Cai, X.; Zheng, X.; Zhang, D.; Iqbal, W.; Liu, C.; Yang, B.; Zhao, X.; Lu, X.; Mao, Y. Microbial characterization of heavy metal resistant bacterial strains isolated from an electroplating wastewater treatment plant. Ecotoxicol. Environ. Saf. 2019, 181, 472–480. [Google Scholar] [CrossRef]
- Tuffin, I.M.; Hector, S.B.; Deane, S.M.; Rawlings, D.E. Resistance determinants of a highly arsenic-resistant strain of Leptospirillum ferriphilum isolated from a commercial biooxidation tank. Appl. Environ. Microbiol. 2006, 72, 2247–2253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fashola, M.O.; Ngole-Jeme, V.M.; Babalola, O.O. Heavy metal pollution from gold mines: Environmental effects and bacterial strategies for resistance. Int. J. Environ. Res. Public Health 2016, 13, 1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergeay, M.; Nies, D.; Schlegel, H.G.; Gerits, J.; Charles, P.; Van Gijsegem, F. Alcaligenes eutrophus CH34 is a facultative chemolithotroph with plasmid-bound resistance to heavy metals. J. Bacteriol. 1985, 162, 328–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The carbohydrate-active enzymes database (CAZy): An expert resource for glycogenomics. Nucleic Acids Res. 2009, 37, D233–D238. [Google Scholar] [CrossRef]
- Galperin, M.Y.; Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 2015, 43, D261–D269. [Google Scholar] [CrossRef] [PubMed]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with GLIMMER. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Goker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [Green Version]
- Ondov, B.D.; Treangen, T.J.; Melsted, P.; Mallonee, A.B.; Bergman, N.H.; Koren, S.; Phillippy, A.M. Mash: Fast genome and metagenome distance estimation using MinHash. Genome Biol. 2016, 17, 132. [Google Scholar] [CrossRef] [Green Version]
- Lagesen, K.; Hallin, P.; Rodland, E.A.; Staerfeldt, H.H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST plus: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Goker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, M.; Rossello-Mora, R.; Glockner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Mazhar, S.H.; Herzberg, M.; Ben Fekih, I.; Zhang, C.K.; Bello, S.K.; Li, Y.P.; Su, J.M.; Xu, J.Q.; Feng, R.W.; Zhou, S.G.; et al. Comparative insights into the complete genome sequence of highly metal resistant Cupriavidus metallidurans strain BS1 isolated from a gold-copper mine. Front. Microbiol. 2020, 11, 47. [Google Scholar] [CrossRef] [Green Version]
- Amadou, C.; Pascal, G.; Mangenot, S.; Glew, M.; Bontemps, C.; Capela, D.; Carrere, S.; Cruveiller, S.; Dossat, C.; Lajus, A.; et al. Genome sequence of the beta-rhizobium Cupriavidus taiwanensis and comparative genomics of rhizobia. Genome Res. 2008, 18, 1472–1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, P.J.; Van Houdt, R.; Moors, H.; Monsieurs, P.; Morin, N.; Michaux, A.; Benotmane, M.A.; Leys, N.; Vallaeys, T.; Lapidus, A.; et al. The complete genome sequence of Cupriavidus metallidurans strain CH34, a master survivalist in harsh and anthropogenic environments. PLoS ONE 2010, 5, e10433. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, M.; Xiao, J.; Hao, L.; Crowley, D.E.; Zhang, Z.; Yu, J.; Huang, N.; Huo, M.; Wu, J. Genome sequence analysis of the naphthenic acid degrading and metal resistant bacterium Cupriavidus gilardii CR3. PLoS ONE 2015, 10, e0132881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Kim, K.H.; Oh, J.; Jeon, C.O. Cupriavidus lacunae sp. nov., isolated from pond-side soil. Antonie Van Leeuwenhoek 2019, 112, 543–551. [Google Scholar] [CrossRef]
- Abbaszade, G.; Szabo, A.; Vajna, B.; Farkas, R.; Szabo, C.; Toth, E. Whole genome sequence analysis of Cupriavidus campinensis S14E4C, a heavy metal resistant bacterium. Mol. Biol. Rep. 2020, 47, 3973–3985. [Google Scholar] [CrossRef]
- Li, X.; Zhang, L.; Wang, G. Genomic evidence reveals the extreme diversity and wide distribution of the arsenic-related genes in Burkholderiales. PLoS ONE 2014, 9, e92236. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.A.; Ellis, D.R.; Huang, X.; Norton, G.J.; Meharg, A.A.; Salt, D.E.; Csonka, L.N. Glutathione-S-transferase from the arsenic hyperaccumulator fern Pteris vittata can confer increased arsenate resistance in Escherichia coli. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Maury, L.; Sanchez-Riego, A.M.; Reyes, J.C.; Florencio, F.J. The glutathione/glutaredoxin system is essential for arsenate reduction in Synechocystis sp. strain PCC 6803. J. Bacteriol. 2009, 191, 3534–3543. [Google Scholar] [CrossRef] [Green Version]
- Hayes, R.P.; Lewis, K.M.; Xun, L.; Kang, C. Catalytic mechanism of 5-chlorohydroxyhydroquinone dehydrochlorinase from the YCII superfamily of largely unknown function. J. Biol. Chem. 2013, 288, 28447–28456. [Google Scholar] [CrossRef] [Green Version]
- Willis, M.A.; Song, F.; Zhuang, Z.; Krajewski, W.; Chalamasetty, V.R.; Reddy, P.; Howard, A.; Dunaway-Mariano, D.; Herzberg, O. Structure of YciI from Haemophilus influenzae (HI0828) reveals a ferredoxin-like alpha/beta-fold with a histidine/aspartate centered catalytic site. Proteins 2005, 59, 648–652. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heavy Metals | C39 | CH34 | BS1 | AE104 |
|---|---|---|---|---|
| As(III) | 2.5 mM | 2.5 mM | 3 mM | 2.5 mM |
| Au(III) | 70 μM | 100 μM | 110 μM | 80 μM |
| Cu(II) | 2 mM | 4.5 mM | 3 mM | 2.5 mM |
| Zn(II) | 2 mM | 12 mM | 12 mM | 0.5 mM |
| Ni(II) | 0.2 mM | 6 mM | 9 mM | 0.5 mM |
| Antibiotics | C39 | CH34 | BS1 | AE104 |
|---|---|---|---|---|
| kanamycin | 256 μg/mL | >256 μg/mL | >256 μg/mL | >256 μg/mL |
| streptomycin | >256 μg/mL | >256 μg/mL | >256 μg/mL | >256 μg/mL |
| gentamycin | >256 μg/mL | >256 μg/mL | >256 μg/mL | >256 μg/mL |
| ampicillin | >128 μg/mL | >128 μg/mL | >128 μg/mL | >128 μg/mL |
| chloramphenicol | 32 μg/mL | >256 μg/mL | >256 μg/mL | >256 μg/mL |
| rifampin | 12 μg/mL | 64 μg/mL | 28 μg/mL | 64 μg/mL |
| tetracycline | 3 μg/mL | 4 μg/mL | 5 μg/mL | 4 μg/mL |
| Genes | Functions | Metal(loid) | Locus Tag |
|---|---|---|---|
| cusA | efflux RND transporter permease subunit | Cu | JJQ59_11025 |
| cusB | efflux RND transporter periplasmic adaptor subunit | Cu | JJQ59_11020 |
| cusC | TolC family protein | Cu | JJQ59_11015 |
| PCuC | copper chaperone PCu(A)C | Cu | JJQ59_01040 |
| copQ1 | copper resistance protein1 | Cu | JJQ59_37555 |
| copQ2 | copper resistance protein | Cu | JJQ59_23855 |
| copS | heavy metal sensor histidine kinase | Cu | JJQ59_26675 |
| copS | heavy metal sensor histidine kinase | Cu | JJQ59_30460 |
| copR | heavy metal response regulator transcription factor | Cu | JJQ59_30455 |
| copA | copper resistance system multicopper oxidase | Cu | JJQ59_30450 |
| copB | copper resistance protein B | Cu | JJQ59_30445 |
| copC | copper homeostasis periplasmic binding protein | Cu | JJQ59_30440 |
| copD | copper homeostasis membrane protein | Cu | JJQ59_30435 |
| ctpA | copper translocating P-type ATPase | Cu | JJQ59_11645 |
| ctpA | copper translocating P-type ATPase | Cu | JJQ59_10950 |
| cueR | Cu(I)-responsive transcriptional regulator | Cu | JJQ59_18470 |
| ctpA | copper-translocating P-type ATPase | Cu | JJQ59_18475 |
| copZ | heavy-metal-associated domain-containing protein | Cu | JJQ59_18480 |
| atpX | heavy metal translocating P-type ATPase | JJQ59_35770 | |
| czcB | efflux RND transporter periplasmic adaptor subunit | Co/Zn/Cd | JJQ59_31505 |
| czcC | TolC family protein | Co/Zn/Cd | JJQ59_31510 |
| czcI | cobalt-zinc-cadmium resistance protein | Co/Zn/Cd | JJQ59_31515 |
| czcA | efflux RND transporter permease subunit | Co/Zn/Cd | JJQ59_27715 |
| czcB | efflux RND transporter periplasmic adaptor subunit | Co/Zn/Cd | JJQ59_27720 |
| czcC | TolC family protein | Co/Zn/Cd | JJQ59_27725 |
| czcI | cobalt-zinc-cadmium resistance protein | Co/Zn/Cd | JJQ59_27730 |
| czcC | TolC family protein | Co/Zn/Cd | JJQ59_20825 |
| czcB | efflux RND transporter periplasmic adaptor subunit | Co/Zn/Cd | JJQ59_20830 |
| czcA | efflux RND transporter permease subunit | Co/Zn/Cd | JJQ59_20835 |
| czcC | TolC family protein | Co/Zn/Cd | JJQ59_35985 |
| czcA | efflux RND transporter permease subunit | Co/Zn/Cd | JJQ59_35990 |
| czcB | efflux RND transporter periplasmic adaptor subunit | Co/Zn/Cd | JJQ59_35995 |
| qseC | sensor histidine kinase N-terminal domain-containing protein | JJQ59_36000 | |
| qseB | winged helix-turn-helix domain-containing protein | JJQ59_36005 | |
| efflux RND transporter periplasmic adaptor subunit | JJQ59_24830 | ||
| efflux RND transporter permease subunit | JJQ59_24835 | ||
| efflux RND transporter periplasmic adaptor subunit | JJQ59_31600 | ||
| dmeF | CDF family Co(II)/Ni(II) efflux transporter DmeF | JJQ59_01450 | |
| Cobalt-zinc-cadmium resistance protein | Co/Zn/Cd | JJQ59_17920 | |
| zntA | heavy metal translocating P-type ATPase | Zn/Cd | JJQ59_26665 |
| zntR | Cd(II)/Pb(II)-responsive transcriptional regulator | Zn/Cd | JJQ59_18160 |
| mgtA | magnesium-translocating P-type ATPase | Mg | JJQ59_37705 |
| corA | magnesium/cobalt transporter(uptake system) | Mg/Co | JJQ59_16020 |
| dmeF | CDF family Co(II)/Ni(II) efflux transporter DmeF | Ni/Co | JJQ59_01450 |
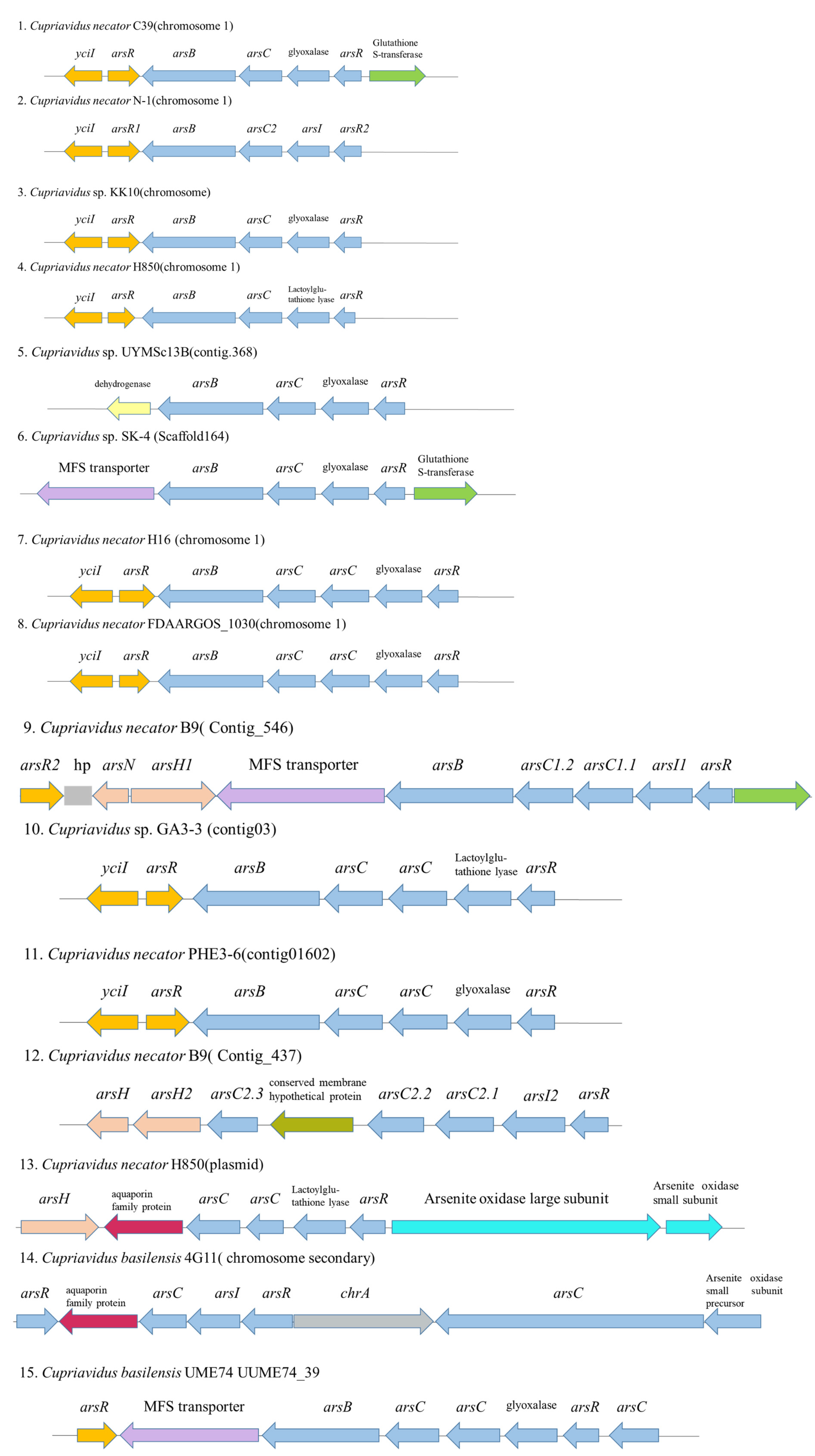
| arsC | arsenate reductase (glutaredoxin) | As | JJQ59_18050 |
| arsenic transporter | As | JJQ59_29185 | |
| arsB | arsenite efflux pump | As | JJQ59_29190 |
| arsB | ACR3 family arsenite efflux transporter | As | JJQ59_10970 |
| arsC | arsenate reductase | As | JJQ59_10975 |
| arsI | glyoxalase/bleomycin resistance/dioxygenase family protein | As | JJQ59_10980 |
| arsR | metalloregulator ArsR/SmtB family | As | JJQ59_10985 |
| arsR1 | metalloregulator ArsR/SmtB family | As | JJQ59_10965 |
| phnB | YciI family protein; putative C-As lyase | As | JJQ59_10960 |
| GST | glutathione S-transferase family protein | As | JJQ59_10990 |
| Compound Name | Gene Symbol | K Number | EC Number | Definition | Annotated Genes |
|---|---|---|---|---|---|
| benzoate | benA | K05549 | EC:1.14.12.10 | benzoate 1,2-dioxygenase alpha subunit | JJQ59_09715 (Chr 1) |
| benB | K05550 | EC:1.14.12.10 | benzoate 1,2-dioxygenase beta subunit | JJQ59_09710 (Chr 1) | |
| benD | K05783 | EC:1.3.1.25 | 1,6-dihydroxycyclohexa-2,4-diene-1-carboxylate dehydrogenase | JJQ59_09700 (Chr 1) | |
| 3-hydroxybenzoate | nagX | K22270 | EC:1.14.13.24 | 3-hydroxybenzoate 6-monooxygenase | JJQ59_22370 (Chr 2) |
| nagI | K00450 | EC:1.13.11.4 | gentisate 1,2-dioxygenase | JJQ59_22355 (Chr 2) | |
| nagL/maiA | K01801 | EC:5.2.1.2 | maleylpyruvate isomerase/maleylacetoacetate isomerase | JJQ59_22365 (Chr 2) | |
| nagK/NA | K16165 | EC:3.7.1.20 | fumarylpyruvate hydrolase/fumarylacetoacetate hydrolase | JJQ59_22365 (Chr 2) | |
| 4-hydroxybenzoate | pobA | K00481 | EC:1.14.13.2 | 4-hydroxybenzoate 3-monooxygenase | JJQ59_30915 (Chr 2) |
| pcaG | K00448 | EC:1.13.11.3 | protocatechuate 3,4-dioxygenase, alpha subunit | JJQ59_30935 (Chr 2) | |
| pcaH | K00449 | EC:1.13.11.3 | protocatechuate 3,4-dioxygenase, beta subunit | JJQ59_30940 (Chr 2) | |
| pcaB | K01857 | EC:5.5.1.2 | 3-carboxy-cis,cis-muconate cycloisomerase | JJQ59_30930 (Chr 2) | |
| pcaC | K01607 | EC:4.1.1.44 | carboxymuconolactone decarboxylase family | JJQ59_25245 (Chr 2) | |
| pcaD | K14727 | EC:3.1.1.24 | 3-oxoadipate enol-lactonase | JJQ59_09740 (Chr 2), JJQ59_26420 (Chr 2) | |
| catechol | catA | K03381 | EC:1.13.11.1 | catechol 1,2-dioxygenase | JJQ59_09720 (Chr 1) |
| catB | K01856 | EC:5.5.1.1 | muconate cycloisomerase | JJQ59_20615 (Chr 2) | |
| catC | K03464 | EC:5.3.3.4 | muconolactone delta-isomerase | JJQ59_09735 (Chr 1) | |
| pcaD | K01055 | EC:3.1.1.24 | 3-oxoadipate enol-lactonase | JJQ59_30925 (Chr 2) | |
| phenol | dmpK | K16249 | EC:1.14.13.244 | phenol hydroxylase P0 protein | JJQ59_20635 (Chr 2) |
| dmpL | K16243 | EC:1.14.13.244 | phenol hydroxylase P1 protein | JJQ59_20640 (Chr 2) | |
| dmpM | K16244 | EC:1.14.13.244 | phenol hydroxylase P2 protein | JJQ59_20645 (Chr 2) | |
| dmpN | K16242 | EC:1.14.13.244 | phenol hydroxylase P3 protein | JJQ59_20650 (Chr 2) | |
| dmpQ | K16245 | EC:1.14.13.244 | phenol hydroxylase P4 protein | JJQ59_20655 (Chr 2) | |
| dmpP | K16246 | EC:1.14.13.244 | phenol hydroxylase P5 protein | JJQ59_20660 (Chr 2) | |
| benzonitrile | NA | K01501 | EC: 3.5.5.1 | nitrilase | JJQ59_09680 (Chr 1) |
| benzamide | amiE | K01426 | EC:3.5.1.4 | amidase | JJQ59_09300 (Chr 1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Z.; Wang, D.; Ben Fekih, I.; Yu, Y.; Li, Y.; Alwathnani, H.; Herzberg, M.; Rensing, C. Whole Genome Sequence Analysis of Cupriavidus necator C39, a Multiple Heavy Metal(loid) and Antibiotic Resistant Bacterium Isolated from a Gold/Copper Mine. Microorganisms 2023, 11, 1518. https://doi.org/10.3390/microorganisms11061518
Xie Z, Wang D, Ben Fekih I, Yu Y, Li Y, Alwathnani H, Herzberg M, Rensing C. Whole Genome Sequence Analysis of Cupriavidus necator C39, a Multiple Heavy Metal(loid) and Antibiotic Resistant Bacterium Isolated from a Gold/Copper Mine. Microorganisms. 2023; 11(6):1518. https://doi.org/10.3390/microorganisms11061518
Chicago/Turabian StyleXie, Zhenchen, Dan Wang, Ibtissem Ben Fekih, Yanshuang Yu, Yuanping Li, Hend Alwathnani, Martin Herzberg, and Christopher Rensing. 2023. "Whole Genome Sequence Analysis of Cupriavidus necator C39, a Multiple Heavy Metal(loid) and Antibiotic Resistant Bacterium Isolated from a Gold/Copper Mine" Microorganisms 11, no. 6: 1518. https://doi.org/10.3390/microorganisms11061518