Abstract
With the practice of a successful livestock industry using antibiotics, which has continued for more than five decades, researchers have long been interested in finding alternatives to antibiotics for poultry production. Probiotics can potentially reduce enteric diseases in livestock and enhance their productivity. The aim of this study was to isolate putative probiotics from camel milk and test them against Salmonella infection as well as host immune development. Thirteen different isolates were obtained from six different camel milk samples from dairy farms in Saudi Arabia. Three of the six isolates (PM1, PM2, PM3, PM4, PM5, and PM6) that showed Gram-positive characters reacted negatively to catalase and hemolytic assays. PM1, PM5, and PM6 showed significant nonpolar surface properties (>51% hydrophobic) and potent antimicrobial activities against avian pathogens, namely S. enterica, S. typhi, S. aureus, and E. coli. PM5 exhibited substantial probiotic traits; therefore, further focus was given to it. PM5 was identified as Bacillus subtilis OQ913924 by the 16S rRNA sequencing method and showed similarity matrix > 99%. An in vivo chicken model was used to access the health benefits of probiotics. After salmonella infection, the mucosal immune response was significantly increased (p < 0.01), and none of the challenge protocols caused mortality or clinical symptoms after infection in intestinal contents. S. enterica organ infiltration in the spleen, thymus, and small intestine was significantly reduced in the B. subtilis PM5-fed chickens. The S. enterica load in chicken feces was reduced from CFU 7.2 to 5.2 in oral-fed B. subtilis PM5-fed chickens. Probiotic-fed chickens showed buffered intestinal content and positively regulated the level of butyric acid (p < 0.05), and intestinal interleukin 1 beta (IL1-β), C-reactive protein (CRP), and interferon gamma (IFN-γ) levels were reduced (p < 0.05). In addition, B. subtilis PM5 showed significant binding to peritoneal macrophages cells and inhibited S. enterica surface adhesion, indicating co-aggregation of B. subtilis PM5 in macrophage cells. It could be concluded that supplementation with probiotics can improve the growth performance of broilers and the quality of broiler chickens against enteric pathogens. The introduction of this probiotic into the commercial poultry feed market in the near future may assist in narrowing the gap that now exists between chicken breeding and consumer demand.
1. Introduction
Camel milk has garnered a lot of interest across the globe because of its medicinal and nutritional benefits [1]. Camel milk has been tested for its microflora, and the results confirm the existence of high levels of microbial diversity, with probiotic bacteria being among the common isolates [2]. Probiotics are defined as live microorganisms that, when administered in sufficient quantities, can provide health benefits to the host [3]. A wide array of probiotic bacterial strains has been isolated from various ecological niches, including camel milk. Examples of this are Pediococcus pentosaceus, Enterococcus faecium, and Enterococcus durans from raw camel milk in Morocco [4], Bacillus subtilis in Iran [5], and Lactiplantibacillus plantarum in Tunisia [6]. A large body of evidence confirmed the significant beneficial impacts of probiotics on humans [7,8], animals [9], and plants [10].
Throughout the world, people rely on poultry as an inexpensive and accessible source of animal protein. In the poultry industry, microbial pathogen infection is a major threat that might compromise food safety. O’Bryan et al. (2022) estimate that 1.35 million people become infected, 26,500 people are hospitalized, and 420 people die per year in the United States due to Salmonella-related foodborne diseases [11]. There are about 2500 different serovars of the enteropathogenic bacterial species S. enterica, which is fairly prevalent. More than 2500 different strains of Salmonella enterica exist, making it one of the most diverse bacterial species. Virulence and antibiotic resistance in Salmonella strains contribute to a higher incidence of disease and death. S. enterica is a common bacterium that causes diarrhea, dehydration, and growth retardation in chickens. In commercial flocks, S. enterica may be transmitted by polluted environments, tainted feeds, and diseased rodents. Furthermore, there are health concerns because S. enterica can easily be transmitted to humans after they have consumed contaminated food. Using antibiotics to treat bacterial illnesses in chickens has the potential to harm human and poultry health and the environment. In addition to presenting serious threats to public health and food security, the widespread misuse of antibiotics is a major contributor to the emergence of infections that may be resistant to the antibiotics now in use. This means that antibiotics may no longer be effective in treating infections in chickens, which can lead to increased mortality rates and decreased productivity. Additionally, the use of antibiotics can disrupt the gut microbiome of chickens, which can lead to digestive problems and other health issues. Antibiotic-resistant bacteria are responsible for an estimated $35 billion in lost productivity in the United States and 4.95 million deaths worldwide by 2022 [12]. In addition to preventing gastrointestinal disorders, lactobacilli has been shown to increase body weight and egg weight in chicken models [13,14]. In addition, bacterial feed (lactobacilli and S. enteritidis) protected against S. enterica infection when used as a mixture outside the chicken host [15].
Bacillus species are well known worldwide and generally recognized as safe (GRAS) bacteria, despite the fact that some of their members are pathogenic [16]. They are widespread, showing up in places including water, plants, soil, food, humans, and animals. There are 105 species with validly published and correct names of Gram-positive, aerobic, and facultative anaerobic rod-shaped cells in the genus Bacillus. The website (https://lpsn.dsmz.de/search?word=Bacillus, accessed on accessed on 15 April 2023) provides a comprehensive list of Bacillus species. Species of Bacillus displayed remarkable positive impacts, including enhancing T-cell responses in chickens [17], alleviating colitis [7], and attenuating neurodegenerative symptoms in mouse models [8].
Probiotics have been reported as a viable antibiotic-sparing method for preventing Salmonella sp. Infections in chicken and enhancing their protein content [18]. Camel’s milk could be a fantastic source for discovering new probiotic bacterium species because it is an ideal growth medium for many microorganisms. This work was designed to isolate potential novel probiotic bacteria from camel milk and test their ability to reduce the incidence of Salmonella infection and enhance general parameters of health in chickens including physical condition.
2. Materials and Methods
2.1. Sample Collection and Bacterial Isolation
Six milk samples (each 200 mL) were obtained from local lactating healthy camels (Camelus dromedaries) from local camel farms in Al-Ahsa, Saudi Arabia. Before sampling, the udder was washed with 50% isopropanol and sterile water and dried with a single-use towel. The first three streams of milk were flushed away. The milk samples were collected in a sterilized 50 mL falcon tube and stored in an icebox. Samples were transported immediately to the laboratory for further analysis. The samples (500 µL) were spread-plated on de Man Rogosa Sharpe (MRS) agar (Himedia, India) after being diluted in sterile saline (0.85% w/v NaCl). After 24 h, morphologically distinct colonies were selected and re-streaked on the MRS agar plate to obtain pure isolates. The chosen colonies were tested again in a glucose yeast peptone (GYP) broth with a pH of 4.5. Successfully growing isolates were biochemically checked. The string test was used to measure Gram staining, the H2O2 drop test was used to check catalase activity, and Mueller-Hinton media (MH)-based blood agar was used to measure hemolysis. Only isolates that were Gram-positive, catalase-positive, and non-hemolytic characteristics were chosen for the subsequent probiotic features.
2.2. Probiotic In Vitro Characterization of Milk Isolates
The measurement of the capacity of isolates to tolerate challenging gastrointestinal conditions such as low gastric pH and bile salt was carried out in accordance with the method mentioned earlier [18]. After 18 h of incubation at 37 °C, isolates were cultured in MRS broth to achieve the necessary cell number (109 log CFU/mL). In brief, the isolates were plated at the appropriate concentration on a modified GYP broth medium that had either pH 3.0 adjusted or contained 0.3% bile salt (Oxgall). CFU/mL were determined in acid/bile-tolerant inoculation isolates by plating 100 μL on MRS agar.
2.3. Cell Surface Hydrophobicity
According to Hairul et al. [19], the hydrophobicity of the isolates was determined by their capacity to adhere to a nonpolar mucosal environment (heptane, Merck Co., Whitehouse Station, NJ, USA). In brief, the isolates of choice were cultured in MRS broth (1 mL), and then heptane (1 mL) was added and vortexed. The polar phase’s optical density was measured at the absorbance (A) of 640 nm using a microplate reader (Biotek, Salem, MA, USA). The numbers show the percentage of hydrophobicity on the cell surface. H% = ([A0 − A]/A0 100), where A is the absorbance of the probiotic suspension after mixing with heptane, and A0 is the absorbance of the probiotic suspension before mixing with heptane. Hydrophobic isolates were defined as those with an adherence value greater than 70%.
2.4. Assessment of Antimicrobial Activity on Avian Pathogens
The probiotic characteristics of isolates were assessed for their antibacterial activities against Salmonella enterica, Salmonella typhimurium, Staphylococcus aureus, and Escherichia coli following the method outlined earlier [19]. Pathogenic isolates were procured from the College of Medicine’s Department of Clinical Microbiology, King Faisal University in Al-Ahsa, Saudi Arabia. Overnight cultures of the potential probiotic isolates were harvested by centrifugation (6000× g for 10 min at 4 °C). Exactly, 150 μL of the cell-free lysate (pH 7) (Sigma-Aldrich, Darmstadt, Germany) was added to cell pellets. Standard antibiotic disc (Std) ciproflaxacin (5 µg/disc) was used to compare the probiotic lysates. After incubating the cell-free lysate (150 μL/well) of the screened isolates for 18 h at 37 °C, the diameter of the inhibition zone was measured to determine antimicrobial activity.
2.5. Molecular Identification Using 16S rRNA Gene Sequencing
Molecular identification of the probiotic bacterial isolates was carried out using 16S rRNA gene sequences. The chosen isolates’ total DNA was isolated with the use of a DNA purification kit (Qiagen, Madison, WI, USA). The primers used were forward 5′-GAGTTTGATCCTGGCTAG-3′ and the reverse 5′-AGAAAGGAGGTGATCCAGCC-3′, and extracted DNA template (25 ng) was combined with PCR master mix (Ampliqon, Odense, Denmark). Electrophoresis on 1% agarose gel was used to examine PCR results for quality control. Clean-up kit (Millipore, Fisher Scientific, Loughborough, UK) instructions were followed to isolate amplicons of the anticipated size. PCR products were sent to the “Macrogen” for sequencing.
2.6. Phylogenetic Analyses
The neighbor-joining approach [20] was used to infer the evolutionary history of the probiotic isolates and other related reference strains. The best tree has a total branch length of 0.20093730. Fourteen different DNA sequences were analyzed. The whole dataset included 1350 unique locations. MEGA7 was used to perform the evolutionary studies [21]. Using the Basic Local Alignment Search Tool (Blast-Bioedit tool), the PM5 16S rRNA gene sequence was analyzed and submitted to the GenBank NCBI database.
2.7. Bacterial Strain Preparation
B. subtilis PM5 was grown in a broth (modified GYP) with a basal chicken diet for 24 h, which yielded the optimum value of chicken feed. After 18 h of aerobic culture in LB, S. enterica was given intraorally using an oral gavage catheter.
2.8. Chickens’ Maintenance
The Animal Ethics approval was obtained from the committee of the Deanship of Scientific Research at King Faisal University in Al-Ahsa, Saudi Arabia for all preclinical chicken feed tests (KFU-REC-2022-AUG-Ethics101). In the city of Al-Ahsa, Saudi Arabia, 21 white leghorn chickens were purchased from a commercial hatchery at the age of 7 weeks. The procured chickens were excluded for infectious and vaccination history. Cages made of wire (100 × 80 × 50 cm) were used to house the chickens. The temperature was controlled at 28 ± 2 °C, and the self-light 12 h period was used in the laboratory. The birds were fed a basic meal and given access to water at will (ad libitum).
2.9. Experimental Design
Twenty-one (n = 21) chickens were randomly allocated into three experimental groups. Group 1 (n = 7, Control) chickens were given a baseline diet. Group 2 (n = 7) aimed to assess the effects of orally gavaged sublethal dose S. enterica (107 CFU /0.1 mL) on growth, immunological organ index, and inflammatory marker indices. Group 3 (n = 7) was studied for the effects of chosen probiotic supplements (B. subtilis PM5) (109 CFU /0.2 mL) on S. enterica-challenged chickens. A basal meal was provided to seven chickens in the control group (Figure 1).
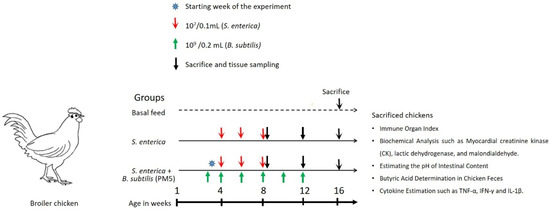
Figure 1.
Experimental design to show the effectiveness of B. subtilis PM5 as a potential probiotic strain using an in vivo chicken model.
For four weeks, the treatment group followed a regular diet supplemented with probiotics. The death rates of infected and probiotics-administered chickens were compared after 4 weeks of survival. After 12 weeks of age, the clinical samples were taken for additional investigations including bursal, feces, spleen, and thymus tissues.
2.10. Immune Organ Index and Growth Performance
On days 4, 7, 14, and 21, the chickens were weighed individually after an overnight fast, and then the data were used to calculate the average body weights on days 4, 7, 14, and 21. Furthermore, at the age of 4 and 8 weeks, the tissues including the bursal, spleen, and thymus gland were separated and weighed after euthanizing each chicken. Then, the immune organ index (immune organ weight, mg/body weight in grams) was calculated.
2.11. Biochemical Analysis
On the 29th day of the trial, the control diet group, a chosen probiotic-supplemented diet, and S. enterica-infected chickens were sacrificed. Blood and liver tissue samples were collected. Myocardial creatinine kinase (CK), lactic dehydrogenase (LDH, A020-2-2), and malondialdehyde (MDA, A003-1-2) activities were assessed. In all biochemical tests, total protein was measured and utilized as an internal control. An Invitrogen avian ELISA kit was used to quantify interferon-gamma (IFN-γ) levels in the blood. The feces of chickens were collected and suspended in 0.1 M PBS. The microbial load of S. enterica was quantified from the clear supernatant on the MRS agar plate.
2.12. Estimating the pH of Intestinal Content
The contents of the probiotic-treated and Salmonella-infected chicken gizzard and ileum were aspirated using sterile PBS, and the pH of the contents from the chicken group was determined using a digital pH meter (Mettler Toledo, Greifensee, Switzerland) [22].
2.13. Butyric Acid Determination in Chicken Feces
Fecal short-chain fatty acids were identified according to the method previously reported [23] with minor alterations. Briefly, butyrate, the microbial response factor, was measured in fecal samples that were collected, mixed with PBS (four equal volumes), and centrifuged at 12,000× g for 15 min. An amount of 1 mM 2-ethylbutyric acid was used as the internal standard. A colorimetric method (Bio-Rad plate reader) was used to quantify the content of butyrate in the fecal samples.
2.14. Cytokine Estimation
The cell-free supernatant of chicken serum was used for the estimation of cytokines such as TNF-α, IFN-γ, and IL-1β) [7], TNF-α, IFN-γ, and IL-1β. The concentrations in S. enterica-infected and PM5-administered chickens were estimated using enzyme-linked immunosorbent assay (ELISA) kits, and values are expressed as pg/mL (Genway, San Diego, CA, USA).
2.15. Assay for Cellular Toxicity and Adhesion
Injecting thioglucolate into mice allowed us to harvest their peritoneal macrophages. Cell counter (Thermo Scientific, Waltham, MA, USA) was used to assess macrophage purity. Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal calf serum was used to cultivate macrophages. In a 24-well plate, 2105 cells were seeded and incubated for 48 h at 37 °C. After 4 h of induction, cells were infected with S. enterica and then washed out with full DMEM. At a multiplicity of infection of 100, S. enterica-infected intestinal cells were injected into overnight fresh cultures of PO2 suspended in fresh serum-free DMEM without antibiotics [19]. The survival of macrophage cells, which S. enterica invaded via intercellular spaces, was measured under these culture conditions.
2.16. Statistical Analysis
The statistical significance of the various treatments was determined using one-way ANOVA on the experimental data at p < 0.05. A student’s t-test was used to compare the statistical analysis of the probiotic diet group to that of the Salmonella group. Microsoft Excel and the SPSS (Statistical Package for the Social Sciences) for Windows (SPSS, Version 10.0, SPSS, Chicago, IL, USA) were used for all statistical analyses.
3. Results
3.1. Characterization of Camel’s Milk Isolates
Totally, ten different colony morphological isolates were isolated from six camel milk. Only six out of ten isolates reacted positively to Gram staining and negatively to catalase reactions and hemolytic assay (Table 1). These isolates were given the designations PM1, PM2, PM3, PM4, PM5, and PM6 and were chosen for further probiotic characterization.

Table 1.
Morphological and physiological characteristics of isolates from camel milk.
3.2. Impacts of Simulated Bile and Gastric Juice on Isolated Probiotics
Figure 2A,B show the survival of the isolates in the presence of MRS at 0.45% bile salts and pH 3.0. Four isolates, PM1, PM4, PM5, and PM6 resisted the stress of bile salt, and the count of viable cells was greater than 2.0 CFU/mL after 180 min in the simulated environment. The other two isolates, on the other hand, did not withstand such conditions. Additionally, the Oxgall resistance of the isolates was proven, and they were able to live after 3 h. At pH 3.0, four isolates had considerably higher viability. After 3 h of incubation, PM1, PM4, PM5, and PM6 showed a significant growth increase. The selected four isolates all had viable cell counts that were more than 30% higher than the set baseline levels (Figure 2A,B).
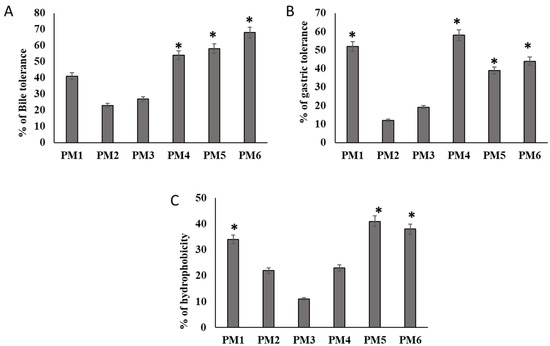
Figure 2.
Characteristics of in vitro probiotic activity shown by isolates obtained from camel milk. (A) Bile salt 0.6% supplemented in MRS agar medium. Log phase probiotic characteristic isolates (M1, M2, M3, M4, M5, and M6) were inoculated in Oxgall MRS media for 3 h and counted the viable cells as log × CFU. (B) The effect of gastric juice tolerance of selected isolates was evaluated in pH 3 in MRS broth. (C) Evaluation of several probiotic isolates with regard to the h numbers indicates means ± SD for triplicate observations. Hydrophobic characteristics of their cell surfaces employing the heptane polarity shift technique. Numbers indicate means ± SD for triplicate findings (significant at * p ≤ 0.05).
3.3. Hydrophobicity of the Cell Surface
The non-polar characteristics of the cell surfaces of considered isolates, as determined by in vitro microbial adherence to heptane droplets, are shown in Figure 2C. The greatest hydrophobic values (>51% hydrophobic nature) were found in PM1, PM5, and PM6 (Figure 2C). The isolates PM1, PM5, and PM6 were evaluated for further clinical antibacterial clearance and in vivo assessment based on these three physiological features.
3.4. Assessment of Antimicrobial Activity against Salmonella spp.
The antimicrobial activity of probiotic isolates was tested using avian disease-causing bacteria. The bactericidal action of secretory metabolic products from probiotics was tested against avian pathogens. The isolates PM1, PM5, and PM6 were shown inhibitory activity against S. typhi, S. enterica and E. coli. PM5 showed significant inhibition against all pathogens compared to other tested probiotics. S. aureus was not sensitive to probiotic products. The inhibitory zone was substantially larger in PM5, showing that it had robust antibiotic action against Salmonella spp. tested. Lysates of PM1, PM5, and PM6 were examined for their anti-Salmonella activities to establish if the possible bioactive chemicals that allow antibacterial activity are intracellular or extracellular. PM5 showed significant pathogenic inhibin against both Salmonella and E. coli strains (Figure 3A,B).
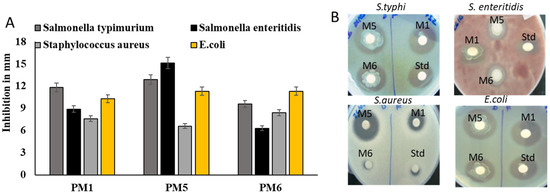
Figure 3.
Assessment of selected probiotics on antimicrobial properties against chicken-specific pathogens E. coli, S. aureus, S. enteritidis, and S. typhi. In order to assess the antibacterial activity of isolated bacterial whole lysate against avian infections, plates of MHA were used. (A) Antimicrobial activity of selected probiotics against avian pathogens and values expressed in mm in diameter (B) MHA plate was used for antimicrobial activity. Std: positive control (Ciprofloxacin 5 µg/disc). The zone of inhibition was calculated in the scale bar and expressed as mm in diameter. Numbers indicate means ± SD for triplicate findings.
3.5. 16S Identification of the Active Isolate
The isolates were characterized based on being Gram-positive, catalase-negative rods and then identified based on the 16S rRNA genes. The obtained sequences showed similarity with those of known species available in the NCBI database (Figure 4). 16S rDNA sequences were submitted to GenBank, and the accession number for PM5 (B. subtilis) (OQ913924) was 99% similar to that for B. subtilis (ON668232). The phylogenetic tree of the neighbor-joining method showed that B. subtilis was 0.001 bootstrap closer to Bacillus species.
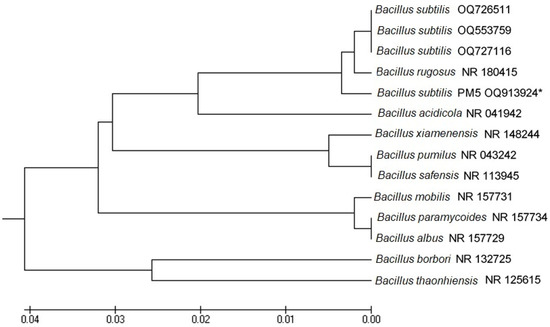
Figure 4.
UPGMA similarity relations of the potential probiotic bacterial strains, Bacillus subtilis OQ913924 with other closely related strains retrieved from NCBI GenBank. Horizontal bars denote the similarity branch length between the isolates. The scale bar was expressed in 0.01 substitutions per nucleotide position. *: indicates our bacterial isolate used in this study.
3.6. Bacillus Supplementation in the Diet Promotes Chicken Growth Performance
Using a chicken model, the probiotic isolate (B. subtilis PM5) was tested for in vivo probiotic properties on chicken immune health. The results showed that the effects of B. subtilis PM5 on chicken development performance, oxidative stress, and inflammatory cytokines are presented in Figure 5 and Figure 6. When PM5 was supplemented with a chicken feed containing 109 CFU/kg, the mortality of the infected chicken was at a higher rate in S. enterica challenged groups than B. subtilis PM5-fed chicken (p < 0.05). Chickens fed with B. subtilis PM5 gained weight, increasing from 427 g to 477 g, and had lower death rates (17 to 34%) (Figure 5). After 4 weeks, significant macroscopic differences in liver tissue color and morphology were observed between S. enterica-infected chickens fed with B. subtilis PM5 and the control group.
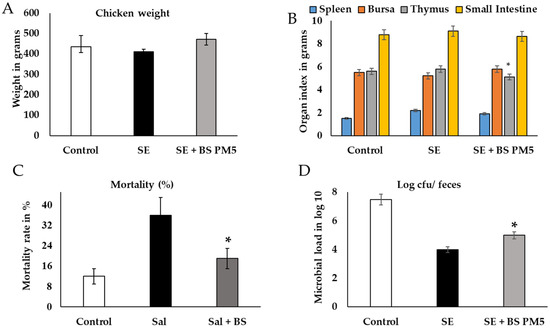
Figure 5.
Macroscopic and microbiota analysis of PM5 was evaluated in Salmonella-challenged chicken. (A) Chicken physiological parameters were evaluated after 28 days of challenge and PM6 administration. The weight of chicken is expressed in grams. (B) The organ index indicates the infection rate and recovery level. (C) The mortality rate in %. (D) Microbial load in fecal samples of chicken and samples were collected and pooled in different intervals, microbial load expressed in log × 107. Data are presented as the average of three independent measurements ± and expressed in respective units. Similar letter group means are insignificantly different (p > 0.05), whereas distinct letter group means are significantly different at * p ≤ 0.05.
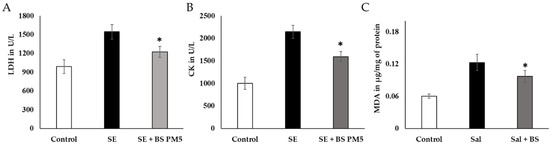
Figure 6.
The impact of a PM6-supplemented chicken feed on the immune system’s reaction to S. enterica challenge in chickens exposed to extreme strain. (A) LDH of liver tissue oxidative stress-related enzymes was evaluated after 4 weeks of acute stress (n = 6). (B). Serum creatinine kinase levels were analyzed to assess cardiac injury following 4 weeks of PM6 supplementation in chicken (n = 6). (C) Neutrophil infiltration was measured in hens that were challenged with Salmonella by measuring the activity of malondialdehyde (MDA). In each plot, the values reflect the mean and standard deviation. For repeated measurements, we used one-way ANOVA and then Tukey’s post hoc testing. * shows p < 0.05.
A B. subtilis PM5-supplemented diet significantly reduced the infection rate of mass in tissues of the spleen and other organs of immunity (thymus) revealing considerably lower S. enterica total count ((p < 0.05). Levels of CFU in the bursa and thymus tissues were considerably higher in the chicken group that was only exposed to S. enterica (p < 0.05). After the treatment period, CFU in the thymus organ elevated substantially (p < 0.01) in the 109 CFU/kg B. subtilis PM5 group (Figure 5A,B). The small intestine of S. enterica-challenged group reduced the CFU of Salmonella in a nonsignificant manner. The fecal load of S. enterica in orally treated S. enterica and B. subtilis PM5 was decreased from CFU 6.8 to 3.9/g. Thus, B. subtilis PM5-supplemented feed reduced S. enterica colonization in the digestive tract significantly (p < 0.01) (Figure 5D). Based on the foregoing findings, B. subtilis PM5-fed hens were given a meal containing 109 CFU/kg for the following pre-clinical tests.
3.7. Bacillus Subtilis Inhibits Oxidative Stress in Chicken
Blood levels of inflammatory and oxidative stress indicators LDH, CK, and MDA were significantly higher in the S. enterica-infected group (p ≤ 0.05; Figure 6A–C), especially 4 weeks after being exposed to the challenge.
There was a significant decrease in serum LDH and CK levels. (p < 0.05) in the B. subtilis PM5-fed group compared to the disease group (Figure 6A,B). MDA, a neutrophil infiltration marker, was found to be considerably higher in the liver tissues of the S. enterica-infected group. In contrast, with S. enterica administration, B. subtilis PM5-fed hens displayed increased oxidative stress activities (p < 0.05) and lower MDA levels (p < 0.05) in the illness group (Figure 6C).
The physiological buffer system improves metabolic activity and hormonal regulations. The pH of different parts of food content in S. enterica-challenged group and B. subtilis-supplemented group showed significant variation. The gizzard food content was not significant, whereas the ileum food content was significantly increased. The range changes from 5.92 ± 0.2 to 6.13 ± 0.2 The short-chain fatty acids improved the gut–brain metabolic interactions and improve the immune system and hormonal regulation (Figure 7A). Butyric acid levels in B. subtilis PM5-fed chickens were higher (p < 0.05) than in the control and post-S. enterica- infected chicken groups. Butyric acid levels in the B. subtilis PM5-fed chicks were significantly increased (p ≤ 0.05) (Figure 7B). Probiotic supplementation has a considerable effect on inflammatory stress indicators. In the pro-inflammatory marker comparison research group, B. subtilis PM5-fed chickens had considerably lower (p < 0.05) intestinal IL-1 and CRP levels, whereas S. enterica-infected birds had lower (p < 0.05) pro-inflammatory cytokine CRP, IL-1, and IFN- levels (Figure 7C–E).
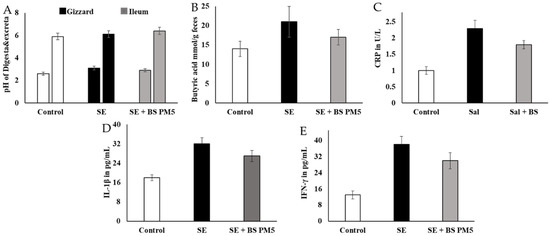
Figure 7.
The influence of PM6 supplementation on the inflammatory response of chickens challenged with S. enterica. (A) pH estimation of different intestinal content (Gizzard and ileum) food content at 20th days of the experiment. (B) Estimation of butyric acid levels in the mucus of S. enterica-challenged chicken colon and PM5 treatment groups. (C) Lymphocyte degradation marker CRP was quantified in the serum samples. (D) IL-1β inflammatory cytokine concentration in S. enterica-challenged chicken serum and PM5 treatment groups. (E) Determination of the IFN-γ content in the serum of S. enterica-challenged chickens and PM5-treated groups as an infection stimulation marker cytokine.
The in vitro findings conclude the interaction and co-aggregation of probiotics with pathogens in host immune cells. The cellular model revealed that both the B. subtilis PM5 and the non-mutual S. enterica displayed considerable binding to peritoneal macrophages in vitro, which was adequate to reduce S. typhimurium colonization (Figure 7). The cytotoxicity of peritoneal cell lines was gradually reduced in B. subtilis PM5-supplemented group. The peritoneal cell viability was increased from 61% to 72% (Figure 8A). S. enterica invasion to peritoneal cells was significantly reduced in the presence of Bacillus PM5 (p < 0.05). Invasion in co-culture groups including S. enterica and B. subtilis PM5 were reported as 29.2 to 13.1 % log CFU/mL, respectively (Figure 8B). The intracellular load of Salmonella was determined, and B. subtilis PM5-fed cell groups had a substantial decrease in invasive Salmonella. These findings showed that B. subtilis PM5 showed potent probiotic activity in overcoming infections by S. enterica (Figure 8A,B). The septic marker TNF-α was used to assess recovery levels. TNF-α levels in S. enterica-infected peritoneal cells were substantially higher than in the infected and control groups. When compared to Salmonella-infected cells, B. subtilis PM5 strains secrete significantly reduced cytokine levels (Figure 8C).
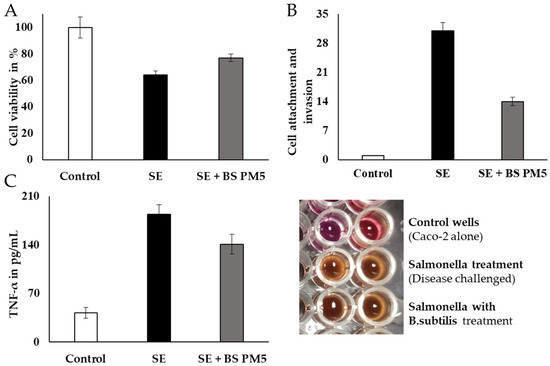
Figure 8.
Peritoneal macrophages (PM5) infected with S. enterica have been studied for their capacity for survival, invasion, and adhesion. (A) The MTT assay was used to measure the viability of B. subtilis infected with Salmonella enterica. (B) The impact of PM6’s adhesion and invasion capabilities on the overall invasion of S. enterica in lysed cell content. TNF in S. enterica-induced pM5 cells (C): PM6’s impact. Mean standard error of the mean (SEM) for triplicate measurements.
4. Discussion
With the continuous use of antibiotics in the livestock business for more than five decades, researchers have long been interested in developing antibiotic alternatives for poultry production. Probiotics are living microorganisms that are added to animal feed as supplements for boosting health and productivity. In this study, we screened raw camel milk for potential probiotic bacteria strains that could improve chicken resistance against Salmonella infections. Out of 13 morphologically distinct isolates, strain PM5 has been selected as a potential probiotic bacterial strain as evidenced by positive Gram staining, negative to catalase and hemolytic tests, the greatest hydrophobic values (>51% hydrophobic nature), and potent antimicrobial activity against avian pathogens, namely S. enterica, S. typhi, S. aureus, and E. coli. Furthermore, PM5 displayed a substantial enhancement in chicken health and their resistance against infections with Salmonella spp. PM5 has been identified as B. subtilis based on the comparative analysis of the 16S rRNA gene sequences.
In order to classify one microbe as a probiotic, a number of attributes should be evaluated at the in vitro and in vivo levels including tolerance to acid and bile salts, hydrophobicity of the cell surface for adhesion, antimicrobial activity, and antibiotic susceptibility [24].
The catalase test for PM5 was negative, indicating that this strain does not generate the catalase enzyme necessary to convert hydrogen peroxide to water and oxygen. This finding also suggests that PM5 can survive on very low levels of oxygen. These results were similar to the study by Saroha et al., who reported Gram-positive, catalase-negative Limosilactobacillus walteri sp. as a novel probiotic antimicrobial lipopeptide-producing bacterium [25]. Hemolysis is responsible for the destruction of host cells, and screening bacteria for these products is critical for ensuring the safety of a single isolate [26]. PM5 had a negative hemolytic response, indicating that it is more likely to be a safe isolate with no risk on the host and appropriate for use as probiotics. This finding is consistent with previous reports of Bacillus subtilis isolated from camel milk as a probiotic candidate [5].
The acid in the stomach and bile salts in the intestine is the basic bio-barrier that a probiotic strain must pass to reach its target [27]. Low-pH conditions may limit metabolic activity and impair probiotic development and survival. Many studies have shown bacterial viability has been demonstrated to decrease after being exposed to stomach acid at pH 2 for 3 h. [28]. Thus, probiotic strains’ survival and growth in the gastrointestinal tract depend on their tolerance to acid, gastric juice, and bile. In our study, PM5 resisted bile salt, acidic pH, and artificial gastric juice. These results are similar to the probiotic Bacillus strain from a previous study [5].
Cell surface hydrophobicity, auto-aggregation, and epithelial cell adherence are other attributes that a probiotic bacterial strain should possess. PM5 showed the highest hydrophobic nature among all other strains. The bacterial film generated by probiotics while sticking to epithelium decreases pathogen–host cell contact [29]. The peritoneal cell line was used to determine specific adherence. In our investigation, the isolate B. subtilis PM5 showed strong adhesion and anti-inflammatory action against S. enterica. The chosen strain demonstrated colonization with only a few infectious S. enterica, adhesion to cells of the epithelium, and development of biofilms according to the hydrophobicity of the surface and auto-aggregation [19,30]. Bacterial adhesion is a complicated mechanism that requires bacterial cell membranes to connect interacting surfaces, giving probiotic bacteria a competitive advantage in undertaking their beneficial action. Although cell surface hydrophobicity has been used as an indicator of in vivo adhesion, a better measure is adhesion assays using cell lines such as Caco2 as a model of the intestinal epithelial barrier. Adhesion experiments that use cell lines such as Caco2 are a better way to measure adhesion in living organisms because they assess the ability of cells to adhere to specific receptors on host cells [31]. The type of cell line (Caco2 or HT29) and bacterial strain are factors that affect the level of bacteria adherence.
PM5 displayed noticeable antimicrobial activities against common avian pathogens, namely S. typhi, S. enterica and E. coli, as evidenced by the wide zone of inhibition. Comparable results have been obtained with Bacillus spp. with probiotic traits [32,33]. Additionally, probiotics have been proven in several in vivo and in vitro investigations to reduce the development of Salmonella [9] and Shigella dysenteriae [33]. The biocidal action of PM5 can be attributed to the bioactive compounds that the PM5 lysate contained, which negatively affect the pathogen’s metabolic activity, growth, and reproduction. DNA replication, protein synthesis, cell wall integrity, biofilm formation, cell signaling, and membrane integrity are among the potential targets of the bio-compounds [34].
Based on the above-mentioned characteristics, PM5 was confirmed as a potential probiotic bacterial strain obtained from fresh camel’s milk. 16S rDNA sequencing identified PM5 as B. subtilis (OQ913924) and was 99.1% similar to that for B. subtilis. Furthermore, PM5 was clearly clustered within the B. subtilis clade based on the phylogenetic tree of 16S rRNA genes from Bacillus spp. confirming the identity of the strain. This result also provides further evidence about the effectiveness of the 16S rRNA gene sequencing as a powerful taxonomic tool for bacterial identification at the genus and species levels [35].
The effectiveness of B. subtilis PM5 as a potential probiotic strain was further confirmed using an in vivo chicken model. PM5 supplementation in the diet promotes chicken growth performance, boosts the immune response, and enhances resistance against Salmonella infections. Similar studies have reported that probiotic strain administration for livestock promotes health and productivity [36]. Peng et al. discovered that a feed enriched with L. plantarum increased weight gain in broiler chickens. B. licheniformis feeding may enhance body mass and average daily gain (ADG) [37,38]. S. enterica, a virulent bacterium, is tightly linked to the chicken gut and causes foodborne diseases that weaken chicken immune systems, resulting in high poultry industry losses [39]. MP5 Supplementation into the diets of chickens infected with S. enterica was an effective approach to preventing S. enterica from evading the immune system and spreading illness. Genes involved in inflammation and certain cytokines that respond to illness, such as interleukin-8 (IL-8) and interferon-gamma (IFN-γ), are among those whose expression is altered by probiotic bacteria. These findings suggest that improving livestock breeding management is the key to ensuring disease-free livestock. Furthermore, adequate additives for feed may assist in the prevention of bacterial infections such as E. coli and S. enterica [23,40]. Earlier research has revealed that L. plantarum supplementation might enhance intestinal health and reduce mortality in chickens infected with E. coli [23]. Furthermore, Bacillus subtilis-based probiotic has been found to improve skeletal health and immunity in broiler chickens exposed to heat stress [41] and alleviate neuroinflammation in the hippocampus via the gut microbiota-brain axis in heat-stressed chickens [42]. The recent findings indicate that PM5 has defensive action for combating S. enterica by lowering the colonization of Salmonella. Butyric acid is produced mainly due to microbial metabolic activities in the intestine and inhibits the growth of pathogenic bacteria [43]. In our investigation, higher amounts of butyric acid and lower detection of Salmonella in the ileum were linked to decreased liver cirrhosis in chickens fed with B. subtilis PM5. In animal husbandry and poultry research, potential probiotics have emerged as an important, safe, potentially efficient, and sustainable host enhancement approach. These findings showed that B. subtilis PM5 could minimize infections caused by bacteria and enhance the quality of broiler chicken. Furthermore, it is possible that the introduction of this probiotic into the commercial poultry feed market in the near future may assist in narrowing the gap that now exists between chicken breeding and consumer demand.
5. Conclusions
Herein, we successfully obtained Bacillus subtilis strain PM5 from raw camel milk. PM5 was confirmed as a potential probiotic bacterial strain based on positive Gram staining, negative to catalase and hemolytic tests, the highest hydrophobic values, and the potent antimicrobial activity against chicken pathogens, S. enterica, S. typhi, S. aureus, and E. coli. Furthermore, PM5 showed a significant improvement in chicken health and resistance to Salmonella spp. infections in an in vivo study. These interesting findings open the door for further exploitation of PM5 as an effective and biocompatible antibiotic alternative for sustaining the poultry industry.
Author Contributions
A.K.: Conceptualization, Data Curation, writing—original draft preparation, review and editing, supervision, project administration and funding acquisition. A.S.; Conceptualization, Methodology and data curation. H.-I.M.I.; Conceptualization, methodology, data curation writing—original draft preparation, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deanship of Scientific Research, King Faisal University, Saudi Arabia for funding this research work through the project number (GRANT3736).
Institutional Review Board Statement
The study was conducted in accordance with code of ethics for the care and use of chicken of the deanship of scientific research ethical committee (KFU-REC-2022-AUG-Ethics101).
Data Availability Statement
Available from the corresponding author upon request.
Acknowledgments
The authors are thankful to the Deanship of scientific research, King Faisal University, Saudi Arabia for funding this research work through the project number (GRANT3736).
Conflicts of Interest
The author declares no conflict of interest.
References
- Konuspayeva, G.; Faye, B. Recent Advances in Camel Milk Processing. Animals 2021, 11, 1045. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Fan, H.; Kwok, L.-Y.; Guo, F.; Ji, R.; Ya, M.; Chen, Y. Analyses of physicochemical properties, bacterial microbiota, and lactic acid bacteria of fresh camel milk collected in Inner Mongolia. J. Dairy Sci. 2020, 103, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Moussaid, S.; El Alaoui, M.A.; Ounine, K.; Benali, A.; Bouhlal, O.; Rkhaila, A.; Hami, H.; El Maadoudi, E.H. In-vitro evaluation of the probiotic potential and the fermentation profile of Pediococcus and Enterococcus strains isolated from Moroccan camel milk. Arch. Microbiol. 2023, 205, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Daneshazari, R.; Khorasgani, M.R.; Hosseini-Abari, A.; Kim, J.-H. Bacillus subtilis isolates from camel milk as probiotic candidates. Sci. Rep. 2023, 13, 3. [Google Scholar] [CrossRef]
- Chouikhi, A.; Ktari, N.; Bardaa, S.; Hzami, A.; Ben Slima, S.; Trabelsi, I.; Asehraou, A.; Ben Salah, R. A novel probiotic strain, Lactiplantibacillus plantarum LC38, isolated from Tunisian camel milk promoting wound healing in Wistar diabetic rats. Arch. Microbiol. 2021, 204, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Khalifa, A.; Sheikh, A.; Ibrahim, H.I.M. Bacillus amyloliquefaciens Enriched Camel Milk Attenuated Colitis Symptoms in Mice Model. Nutrients 2022, 14, 1967. [Google Scholar] [CrossRef]
- Ibrahim, H.I.M.; Sheikh, A.; Khalil, H.E.; Khalifa, A. Bacillus amyloliquifaciens-Supplemented Camel Milk Suppresses Neuroinflammation of Autoimmune Encephalomyelitis in a Mouse Model by Regulating Inflammatory Markers. Nutrients 2023, 15, 550. [Google Scholar] [CrossRef]
- Khalifa, A.; Ibrahim, H.I.M. Enterococcus faecium from chicken feces improves chicken immune response and alleviates Salmonella infections: A pilot study. J. Anim. Sci. 2023, 101, skad016. [Google Scholar] [CrossRef]
- Al-Tammar, F.K.; Khalifa, A.Y.Z. Plant growth promoting bacteria drive food security. Braz. J. Biol. 2022, 82, 1944. [Google Scholar] [CrossRef]
- O’Bryan, C.A.; Ricke, S.C.; Marcy, J.A. Public health impact of Salmonella spp. on raw poultry: Current concepts and future prospects in the United States. Food Control. 2021, 132, 108539. [Google Scholar] [CrossRef]
- Murray, C.J.L.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Talebi, A.; Amirzadeh, B.; Mokhtari, B.; Gahri, H. Effects of a multi-strain probiotic (PrimaLac) on performance and antibody responses to Newcastle disease virus and infectious bursal disease virus vaccination in broiler chickens. Avian Pathol. 2008, 37, 509–512. [Google Scholar] [CrossRef]
- Taha-Abdelaziz, K.; Astill, J.; Kulkarni, R.R.; Read, L.R.; Najarian, A.; Farber, J.M.; Sharif, S. In vitro assessment of immunomodulatory and anti-Campylobacter activities of probiotic lactobacilli. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Juricova, H.; Matiasovicova, J.; Faldynova, M.; Sebkova, A.; Kubasova, T.; Prikrylova, H.; Karasova, D.; Crhanova, M.; Havlickova, H.; Rychlik, I. Probiotic Lactobacilli Do Not Protect Chickens against Salmonella Enteritidis Infection by Competitive Exclusion in the Intestinal Tract but in Feed, Outside the Chicken Host. Microorganisms 2022, 10, 219. [Google Scholar] [CrossRef]
- Gadotti, C. Control of Pathogenic Bacteria in Queso Fresco by Using Generally Recognized as Safe Ingredients. Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, 2011. [Google Scholar]
- Larsberg, F.; Sprechert, M.; Hesse, D.; Loh, G.; Brockmann, G.A.; Kreuzer-Redmer, S. Probiotic Bacillus Strains Enhance T Cell Responses in Chicken. Microorganisms 2023, 11, 269. [Google Scholar] [CrossRef]
- El Jeni, R.; Dittoe, D.K.; Olson, E.G.; Lourenco, J.; Corcionivoschi, N.; Ricke, S.C.; Callaway, T.R. Probiotics and potential applications for alternative poultry production systems. Poult. Sci. 2021, 100, 101156. [Google Scholar] [CrossRef]
- Islam, V.I.H.; Saravanan, S.; Raj, J.P.P.; Paulraj, M.G.; Ignacimuthu, S. Myroides pelagicus from the Gut of Drosophila melanogaster Attenuates Inflammation on Dextran Sodium Sulfate-Induced Colitis. Dig. Dis. Sci. 2014, 59, 1121–1133. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Sozcu, A. Growth performance, pH value of gizzard, hepatic enzyme activity, immunologic indicators, intestinal histomorphology, and cecal microflora of broilers fed diets supplemented with processed lignocellulose. Poult. Sci. 2019, 98, 6880–6887. [Google Scholar] [CrossRef]
- Ding, S.; Wang, Y.; Yan, W.; Li, A.; Jiang, H.; Fang, J. Effects of Lactobacillus plantarum 15-1 and fructooligosaccharides on the response of broilers to pathogenic Escherichia coli O78 challenge. PLoS ONE 2019, 14, e0212079. [Google Scholar] [CrossRef]
- Ayyash, M.M.; Abdalla, A.K.; AlKalbani, N.S.; Baig, M.A.; Turner, M.S.; Liu, S.Q.; Shah, N.P. Invited Review: Characterization of New Probiotics from Dairy and Nondairy Products—Insights into Acid Tolerance, Bile Metabolism and Tolerance, and Adhesion Capability. J. Dairy Sci. 2021, 104, 8363–8379. [Google Scholar] [CrossRef] [PubMed]
- Saroha, T.; Sharma, S.; Choksket, S.; Korpole, S.; Patil, P.B. Limosilactobacillus walteri sp. nov., a novel probiotic antimicrobial lipopeptide-producing bacterium. FEMS Microbiol. Lett. 2023, 370, fnad004. [Google Scholar] [CrossRef] [PubMed]
- Haranahalli Nataraj, B.; Behare, P.V.; Yadav, H.; Srivastava, A.K. Emerging Pre-Clinical Safety Assessments for Potential Pro-biotic Strains: A Review. Crit. Rev. Food Sci. Nutr. 2023, 1–29. [Google Scholar] [CrossRef]
- Tremblay, A.; Auger, J.; Alyousif, Z.; Calero, S.E.C.; Mathieu, O.; Rivero-Mendoza, D.; Elmaoui, A.; Dahl, W.J.; Tompkins, T. Total Transit Time and Probiotic Persistence in Healthy Adults: A Pilot Study. J. Neurogastroenterol. Motil. 2023, 29, 218–228. [Google Scholar] [CrossRef]
- Broom, L.J.; Kogut, M.H. Gut immunity: Its development and reasons and opportunities for modulation in monogastric production animals. Anim. Health Res. Rev. 2018, 19, 46–52. [Google Scholar] [CrossRef]
- Noohi, N.; Papizadeh, M.; Rohani, M.; Talebi, M.; Pourshafie, M.R. Screening for probiotic characters in lactobacilli isolated from chickens revealed the intra-species diversity of Lactobacillus brevis. Anim. Nutr. 2021, 7, 119–126. [Google Scholar] [CrossRef]
- Mandal, A.; Mandal, R.K.; Yang, Y.; Khatri, B.; Kong, B.-W.; Kwon, Y.M. In vitro characterization of chicken gut bacterial isolates for probiotic potentials. Poult. Sci. 2021, 100, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Silva-Dias, A.; Miranda, I.; Branco, J.; Monteiro-Soares, M.; Pina-Vaz, C.; Rodrigues, A.G. Adhesion, biofilm formation, cell surface hydrophobicity, and antifungal planktonic susceptibility: Relationship among Candida spp. Front. Microbiol. 2015, 6, 205. [Google Scholar] [CrossRef]
- Simon, A.; Colom, J.; Mazhar, S.; Khokhlova, E.; Deaton, J.; Rea, K. Bacillus megaterium Renuspore® as a potential probiotic for gut health and detoxification of unwanted dietary contaminants. Front. Microbiol. 2023, 14, 1125616. [Google Scholar] [CrossRef]
- Tran, C.; Horyanto, D.; Stanley, D.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Properties of Bacillus Probiotics as Animal Growth Promoters. Antibiotics 2023, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, G.; Murali, M.R.; Devaraj, S.N. Protective role of lactobacilli in Shigella dysenteriae 1–induced diarrhea in rats. Nutrition 2007, 23, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Nakharuthai, C.; Boonanuntanasarn, S.; Kaewda, J.; Manassila, P. Isolation of Potential Probiotic Bacillus spp. from the Intestine of Nile Tilapia to Construct Recombinant Probiotic Expressing CC Chemokine and Its Effectiveness on Innate Immune Responses in Nile Tilapia. Animals 2023, 13, 986. [Google Scholar] [CrossRef]
- Markowiak, P.; Śliżewska, K. The role of probiotics, prebiotics and synbiotics in animal nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef]
- Peng, Q.; Zeng, X.F.; Zhu, J.L.; Wang, S.; Liu, X.T.; Hou, C.L.; Thacker, P.A.; Qiao, S.Y. Effects of dietary Lactobacillus plantarum B1 on growth performance, intestinal microbiota, and short chain fatty acid profiles in broiler chickens. Poult. Sci. 2016, 95, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Yu, Y.-H. Bacillus licheniformis–fermented products improve growth performance and the fecal microbiota community in broilers. Poult. Sci. 2019, 99, 1432–1443. [Google Scholar] [CrossRef]
- Rashid, S.; Tahir, S.; Akhtar, T.; Altaf, S.; Ashraf, R.; Qamar, W. Bacillus-Based Probiotics: An Antibiotic Alternative for the Treatment of Salmonellosis in Poultry. Pak. Vet. J. 2023, 43, 361–368. [Google Scholar]
- Forkus, B.; Ritter, S.; Vlysidis, M.; Geldart, K.; Kaznessis, Y.N. Antimicrobial Probiotics Reduce Salmonella enterica in Turkey Gastrointestinal Tracts. Sci. Rep. 2017, 7, 40695. [Google Scholar] [CrossRef]
- Jiang, S.; Yan, F.F.; Hu, J.Y.; Mohammed, A.; Cheng, H.W. Bacillus subtilis-Based Probiotic Improves Skeletal Health and Im-munity in Broiler Chickens Exposed to Heat Stress. Animals 2021, 11, 1494. [Google Scholar] [CrossRef]
- Fu, Y.; Hu, J.; Cheng, H.-W. Research Note: Probiotic, Bacillus subtilis, alleviates neuroinflammation in the hippocampus via the gut microbiota-brain axis in heat-stressed chickens. Poult. Sci. 2023, 102, 102635. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ishfaq, M.; Guo, Y.; Chen, C.; Li, J. Assessment of Probiotic Properties of Lactobacillus salivarius Isolated From Chickens as Feed Additives. Front. Veter- Sci. 2020, 7, 415. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).