


A Severe Acute Respiratory Syndrome Coronavirus 2 Anti-Spike Immunoglobulin G Assay: A Robust Method for Evaluation of Vaccine Immunogenicity Using an Established Correlate of Protection
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Assay Procedure
2.2. Samples
2.3. Validation Assays
2.3.1. Precision
2.3.2. Selectivity
2.3.3. Specificity
2.3.4. Matrix Effects
2.3.5. Linearity
2.3.6. Sensitivity
2.3.7. Assay Robustness (Incubation Time and Plate Coating Time)
2.3.8. Sample Stability
2.4. Variant Assays
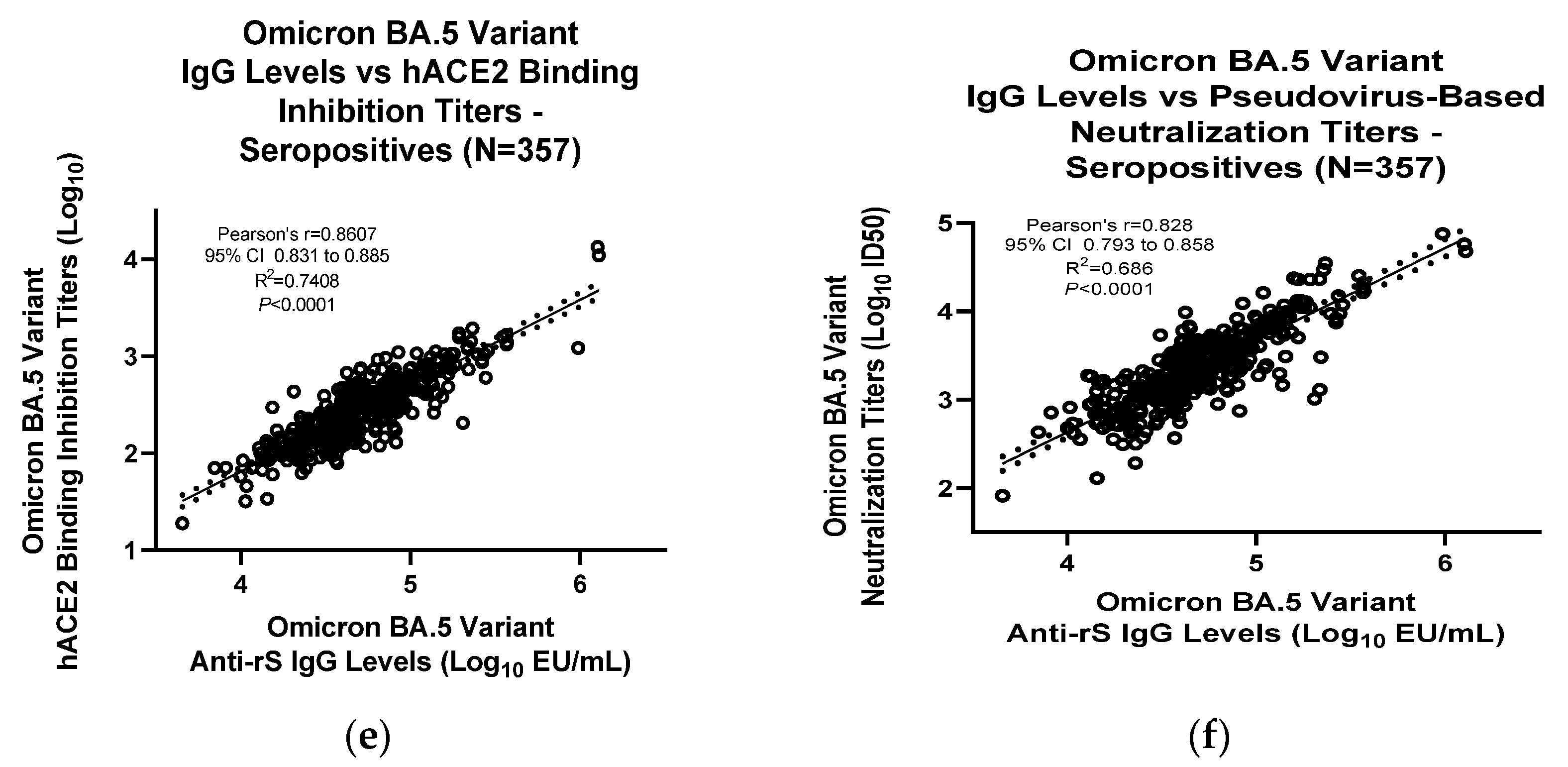
2.5. Correlation Analyses
2.6. Conversion to WHO International Units
3. Results
3.1. Ancestral Strain Assay Validation Parameters
3.2. Variant Strain Assay Validation
3.3. Assay Correlation with Other Markers
3.4. Conversion to WHO International Standard Units (BAU/mL)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. David J. Spencer CDC Museum—COVID-19 Timeline. Available online: https://www.cdc.gov/museum/timeline/covid19.html (accessed on 3 February 2023).
- Fendler, A.; de Vries, E.G.E.; GeurtsvanKessel, C.H.; Haanen, J.B.; Wörmann, B.; Turajlic, S.; von Lilienfeld-Toal, M. COVID-19 vaccines in patients with cancer: Immunogenicity, efficacy and safety. Nat. Rev. Clin. Oncol. 2022, 19, 385–401. [Google Scholar] [CrossRef] [PubMed]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: A narrative review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rocklöv, J. The effective reproductive number of the Omicron variant of SARS-CoV-2 is several times relative to Delta. J. Travel Med. 2022, 29, taac037. [Google Scholar] [CrossRef] [PubMed]
- Van Tilbeurgh, M.; Lemdani, K.; Beignon, A.-S.; Chapon, C.; Tchitchek, N.; Cheraitia, L.; Marcos Lopez, E.; Pascal, Q.; Le Grand, R.; Maisonnasse, P.; et al. Predictive Markers of Immunogenicity and Efficacy for Human Vaccines. Vaccines 2021, 9, 579. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.; Huang, Y.; Benkeser, D.; Carpp, L.N.; Áñez, G.; Woo, W.; McGarry, A.; Dunkle, L.M.; Cho, I.; Houchens, C.R.; et al. Immune Correlates Analysis of the PREVENT-19 COVID-19 Vaccine Efficacy Clinical Trial. medRxiv 2022, 14, 1581. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Pang, N.Y.-L.; Pang, A.S.-R.; Chow, V.T.; Wang, D.-Y. Understanding neutralising antibodies against SARS-CoV-2 and their implications in clinical practice. Mil. Med. Res. 2021, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.H.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.D.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef] [PubMed]
- Bentley, E.M.; Atkinson, E.; Rigsby, P.; Elsley, W.; Bernasconi, V.; Kristiansen, P.; Harvala, H.; Turtle, L.C.; Dobson, S.; Wendel, S.; et al. Establishment of the 2nd WHO International Standard for Anti-SARS-CoV-2 Immunoglobulin and Reference Panel for Antibodies; WHO/BS/2022.2427. 2022. Available online: https://www.who.int/publications/m/item/who-bs-2022.2427 (accessed on 3 February 2023).
- Mattiuzzo, G.; Bentley, E.M.; Hassall, M.; Routley, S.; Richardson, S.; Bernasconi, V.; Kristiansen, P.; Harvala, H.; Roberts, D.; Semple, M.G.; et al. Establishment of the WHO International Standard and Reference Panel for Anti-SARS-CoV-2 Antibody; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Shinde, V.; Cho, I.; Plested, J.S.; Agrawal, S.; Fiske, J.; Cai, R.; Zhou, H.; Pham, X.; Zhu, M.; Cloney-Clark, S.; et al. Comparison of the safety and immunogenicity of a novel Matrix-M-adjuvanted nanoparticle influenza vaccine with a quadrivalent seasonal influenza vaccine in older adults: A phase 3 randomised controlled trial. Lancet Infect. Dis. 2022, 22, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Formica, N.; Mallory, R.; Albert, G.; Robinson, M.; Plested, J.S.; Cho, I.; Robertson, A.; Dubovsky, F.; Glenn, G.M. Different dose regimens of a SARS-CoV-2 recombinant spike protein vaccine (NVX-CoV2373) in younger and older adults: A phase 2 randomized placebo-controlled trial. PLoS Med. 2021, 18, e1003769. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Borisov, O.; Kee, J.J.; Carpp, L.N.; Wrin, T.; Cai, S.; Sarzotti-Kelsoe, M.; McDanal, C.; Eaton, A.; Pajon, R.; et al. Calibration of two validated SARS-CoV-2 pseudovirus neutralization assays for COVID-19 vaccine evaluation. Sci. Rep. 2021, 11, 23921. [Google Scholar] [CrossRef] [PubMed]
- Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Massuda, E.; Patel, U.; Klindworth, A.; Massare, M.J.; Cai, R.; Fries, L.; Glenn, G.; et al. Severe Acute Respiratory Syndrome Coronavirus 2 Receptor (Human Angiotensin-Converting Enzyme 2) Binding Inhibition Assay: A Rapid, High-Throughput Assay Useful for Vaccine Immunogenicity Evaluation. Microorganisms 2023, 11, 368. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.; Klumpp-Thomas, C.; Kalish, H.; Shunmugavel, A.; Mehalko, J.; Denson, J.P.; Snead, K.R.; Drew, M.; Corbett, K.S.; Graham, B.S.; et al. Serologic Cross-Reactivity of SARS-CoV-2 and Endemic and Seasonal Betacoronaviruses. medRxiv 2021, 41, 906–913. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Anti-rS Protein Ab GMC (EU/mL) | Inter-Assay %GCV | Intra-Assay %GCV | Total %GCV |
|---|---|---|---|---|
| Overall 2 | N/A | 0.9 | 13.5 | 13.5 |
| 1 | 47,648 | 0 | 13.7 | 13.7 |
| 2 | 16,537 | 5.8 | 13.6 | 14.9 |
| 3 | 234 | 6.7 | 11.0 | 12.9 |
| 4 | 9303 | 7.2 | 17.2 | 18.7 |
| 5 | 180,688 | 0 | 7.8 | 7.8 |
| 6 | 202,618 | 6.4 | 8.8 | 10.9 |
| 7 | 56,315 | 2.5 | 7.7 | 8.1 |
| 8 | 2146 | 0 | 14.2 | 14.2 |
| 9 | 4890 | 0 | 14.1 | 14.1 |
| 10 | 1447 | 0 | 14.0 | 14.0 |
| 11 | 105,191 | 0 | 9.6 | 9.6 |
| 12 | 510 | 0 | 14.6 | 14.6 |
| 13 | 1067 | 3.3 | 12.3 | 12.7 |
| 14 | 187 | 1.9 | 9.4 | 9.6 |
| 15 | 4629 | 0 | 12.8 | 12.8 |
| 16 | 824 | 0 | 14.5 | 14.5 |
| 17 | 35,042 | 0 | 10.8 | 10.8 |
| 18 | 74,679 | 0 | 9.6 | 9.6 |
| 19 | 106,859 | 10.9 | 14.1 | 17.8 |
| 20 | 7075 | 0 | 13.5 | 13.5 |
| 21 | 101,122 | 0 | 16.6 | 16.6 |
| 22 | 26,750 | 9.4 | 10.6 | 14.2 |
| 23 | 12,125 | 0 | 14.1 | 14.1 |
| 24 | 3358 | 8.5 | 16.0 | 18.2 |
| 25 (HQC) | 30,518 | 0 | 14.1 | 14.1 |
| 26 (LQC) | 1671 | 0 | 18.3 | 18.3 |
| 27 (NC) 3 | 28 | 0 | 17.4 | 17.4 |
| Assay Specificity | ||||||
|---|---|---|---|---|---|---|
| Sample | Sample Source | Assay Buffer | Incubated with Homologous rS Protein, 4 µg/mL | Incubated with Homologous rS Protein, 2 µg/mL | ||
| Ab (EU/mL) | Ab (EU/mL) | % Inhibition | Ab (EU/mL) | % Inhibition | ||
| 1 | Phase 1 trial | 16,077 | 877 | 94.5 | 1699 | 89.4 |
| 2 | Phase 1 trial | 1219 | 139 | 88.6 | 224 | 81.6 |
| 3 | Phase 1 trial | 131,959 | 6791 | 94.9 | 12,057 | 90.9 |
| 4 | Phase 1 trial | 35,640 | 2161 | 93.9 | 2446 | 93.1 |
| 5 | Phase 1 trial | 6353 | 632 | 90.1 | 1015 | 84.0 |
| 6 | Phase 1 trial | 574 | 67 | 88.3 | 94 | 83.6 |
| Antibody Cross-reactivity | ||||||
| MERS-CoV rS Protein | ||||||
| Sample | Sample Source | Assay Buffer | Incubated with MERS-CoV rS Protein, 4 µg/mL | Incubated with MERS-CoV rS Protein, 2 µg/mL | ||
| Ab (EU/mL) | Ab (EU/mL) | % Inhibition | Ab (EU/mL) | % Inhibition | ||
| 1 | Phase 1 trial | 18,575 | 17,933 | 3.5 | 17,812 | 4.1 |
| 2 | Phase 1 trial | 1764 | 1448 | 17.9 | 1443 | 18.2 |
| 3 | Phase 1 trial | 132,825 | 109,291 | 17.7 | 110,509 | 16.8 |
| 4 | Phase 1 trial | 31,774 | 26,580 | 16.3 | 27,783 | 12.6 |
| 5 | Phase 1 trial | 7799 | 6404 | 17.9 | 6361 | 18.4 |
| 6 | Phase 1 trial | 646 | 484 | 25.1 | 586 | 9.3 |
| SARS-CoV-1 rS Protein | ||||||
| Sample | Sample Source | Assay Buffer | Incubated with SARS-CoV-1rS Protein, 4 µg/mL | Incubated with SARS-CoV-1rS Protein, 2 µg/mL | ||
| Ab (EU/mL) | Ab (EU/mL) | % Inhibition | Ab (EU/mL) | % Inhibition | ||
| 1 | Phase 1 trial | 19,392 | 16,789 | 13.4 | 17,769 | 8.4 |
| 2 | Phase 1 trial | 1765 | 1512 | 14.3 | 1551 | 12.1 |
| 3 | Phase 1 trial | 130,229 | 100,597 | 22.8 | 127,364 | 2.2 |
| 4 | Phase 1 trial | 29,804 | 30,897 | −3.7 | 28,603 | 4.0 |
| 5 | Phase 1 trial | 6384 | 5501 | 13.8 | 5290 | 17.1 |
| 6 | Phase 1 trial | 531 | 478 | 10.0 | 457 | 13.9 |
| Irrelevant Proteins (RSV F Protein, Influenza HA) | ||||||
| Sample | Sample Source | Assay Buffer | Incubated with RSV F Protein, 4 µg/mL | Influenza A/Kansas/14/2017 VLP, 4 HA Units | ||
| Ab (EU/mL) | Ab (EU/mL) | % Inhibition | Ab (EU/mL) | % Inhibition | ||
| 1 | Phase 1 trial | 20,879 | 18,936 | 9.3 | 17,321 | 17.0 |
| 2 | Phase 1 trial | 1936 | 1680 | 13.2 | 1725 | 10.9 |
| 3 | Phase 1 trial | 153,044 | 120,742 | 21.1 | 133,650 | 12.7 |
| 4 | Phase 1 trial | 34,951 | 31,896 | 8.7 | 33,289 | 4.8 |
| 5 | Phase 1 trial | 7791 | 7035 | 9.7 | 6893 | 11.5 |
| 6 | Phase 1 trial | 671 | 626 | 6.7 | 645 | 3.9 |
| Heme Matrix Effects | |||||
|---|---|---|---|---|---|
| Sample | Control | 50% Hemolyzed | 25% Hemolyzed | ||
| Ab (EU/mL) | Ab (EU/mL) | % Recovery | Ab (EU/mL) | % Recovery | |
| 1 | 2931 | 2821 | 96.2 | 2954 | 100.8 |
| 2 | 31,541 | 30,494 | 96.7 | 33,161 | 105.1 |
| 3 | 4101 | 3969 | 96.8 | 4137 | 100.9 |
| 4 | 61,022 | 63,570 | 104.2 | 63,406 | 103.9 |
| 5 | 342 | 324 | 94.7 | 351 | 102.6 |
| 6 | 40 (<200) | 33 (<200) | N/A | 27 (<200) | N/A |
| Lipid Matrix Effects | |||||
| Sample | Control | 5.0 mg/mL Triglycerides | 2.5 mg/mL Triglycerides | ||
| Ab (EU/mL) | Ab (EU/mL) | % Recovery | Ab (EU/mL) | % Recovery | |
| 1 | 2931 | 3173 | 108.3 | 2748 | 93.8 |
| 2 | 31,541 | 32,109 | 101.8 | 29,970 | 95.0 |
| 3 | 4101 | 4260 | 103.9 | 4339 | 105.8 |
| 4 | 61,022 | 72,766 | 119.2 | 73,059 | 119.7 |
| 5 | 342 | 351 | 102.6 | 315 | 92.1 |
| 6 | 40 (<200) | 25 (<200) | N/A | 25 (<200) | N/A |
| Sample | Parameter | Estimate | 95% LCL | 95% UCL |
|---|---|---|---|---|
| 1 | Slope | 1.034 | 1.022 | 1.046 |
| Intercept | −0.171 | −0.220 | −0.121 | |
| Residual Variability (% GSD) | 0.015 (3.4%) | N/A | ||
| R2 | 0.9998 | N/A | ||
| 2 | Slope | 1.011 | 0.999 | 1.022 |
| Intercept | −0.046 | −0.091 | −0.001 | |
| Residual Variability (% GSD) | 0.016 (3.8%) | N/A | ||
| R2 | 0.9998 | N/A | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, M.; Cloney-Clark, S.; Feng, S.-l.; Parekh, A.; Gorinson, D.; Silva, D.; Skonieczny, P.; Wilson, A.; Kalkeri, R.; Woo, W.; et al. A Severe Acute Respiratory Syndrome Coronavirus 2 Anti-Spike Immunoglobulin G Assay: A Robust Method for Evaluation of Vaccine Immunogenicity Using an Established Correlate of Protection. Microorganisms 2023, 11, 1789. https://doi.org/10.3390/microorganisms11071789
Zhu M, Cloney-Clark S, Feng S-l, Parekh A, Gorinson D, Silva D, Skonieczny P, Wilson A, Kalkeri R, Woo W, et al. A Severe Acute Respiratory Syndrome Coronavirus 2 Anti-Spike Immunoglobulin G Assay: A Robust Method for Evaluation of Vaccine Immunogenicity Using an Established Correlate of Protection. Microorganisms. 2023; 11(7):1789. https://doi.org/10.3390/microorganisms11071789
Chicago/Turabian StyleZhu, Mingzhu, Shane Cloney-Clark, Sheau-line Feng, Anand Parekh, Drew Gorinson, David Silva, Paul Skonieczny, Adjele Wilson, Raj Kalkeri, Wayne Woo, and et al. 2023. "A Severe Acute Respiratory Syndrome Coronavirus 2 Anti-Spike Immunoglobulin G Assay: A Robust Method for Evaluation of Vaccine Immunogenicity Using an Established Correlate of Protection" Microorganisms 11, no. 7: 1789. https://doi.org/10.3390/microorganisms11071789
APA StyleZhu, M., Cloney-Clark, S., Feng, S.-l., Parekh, A., Gorinson, D., Silva, D., Skonieczny, P., Wilson, A., Kalkeri, R., Woo, W., Cai, M. R., Fries, L., Glenn, G., & Plested, J. S. (2023). A Severe Acute Respiratory Syndrome Coronavirus 2 Anti-Spike Immunoglobulin G Assay: A Robust Method for Evaluation of Vaccine Immunogenicity Using an Established Correlate of Protection. Microorganisms, 11(7), 1789. https://doi.org/10.3390/microorganisms11071789