Abstract
Multidrug-resistant (MDR) Pseudomonas aeruginosa infections represent a major public health concern and require comprehensive understanding of their genetic makeup. This study investigated the first occurrence of imipenemase (IMP)-carrying P. aeruginosa strains from Bulgaria. Whole genome sequencing identified a novel plasmid-mediated IMP-100 allele located in a a novel In4886 integron embedded in a putative Tn7700 transposon. Two other closely related chromosomal IMP variants, IMP-13 and IMP-84, were also detected. The IMP-producers were resistant to last-line drugs including cefiderocol (CFDC) (two out of three) and susceptible to colistin. The IMP-13/84 cassettes were situated in a In320 integron inserted in a Tn5051-like transposon as previously reported. Lastly, the p4782-IMP plasmid rendered the PA01 transformant resistant to CFDC, suggesting a transferable CFDC resistance. A variety of virulence factors associated with adhesion, antiphagocytosis, iron uptake, and quorum sensing, as well as secretion systems, toxins, and proteases, were confirmed, suggesting significant pathogenic potential consistent with the observed strong biofilm formation. The emergence of IMP-producing MDR P. aeruginosa is alarming as it remains unsusceptible even to last-generation drugs like CFDC. Newly detected IMP-100 was even located in a CFDC-resistant XDR strain.
1. Introduction
Pseudomonas aeruginosa is a member of the ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species) group of highly resistant pathogens and represents a formidable challenge in healthcare settings [1]. In the past decade, the emergence of carbapenem resistance among P. aeruginosa isolates has become a global concern as carbapenems are among the last options considered for treatment [2,3]. Carbapenem resistance is commonly associated with mutations in the outer-membrane porin OprD, overexpression of efflux pumps, and horizontal acquisition of carbapenemase genes, including both serine- and metallo-β-lactamases (MBLs) [3,4,5,6].
Beta-lactamases encoded by bla genes are grouped into four classes (A, B, C, D) according to the widely used Ambler classification. Carbapenemase activity in clinical strains of P. aeruginosa due to Klebsiella pneumoniae carbapenemase (KPC) and certain Guiana extended-spectrum β-lactamase (GES) variants (Ambler class A) are less frequently reported, similar to the oxacillinases (OXA) from Ambler class D with activity against carbapenems [7]. MBLs (Ambler class B) are characterized by their dependency on one or two zinc cations for enzymatic activity [4]. Various types of acquired MBLs have been identified in P. aeruginosa, including imipenemase (IMP), Verona integron metallo-beta-lactamase (VIM), São Paulo metallo-beta-lactamase (SPM), Germany imipenemase (GIM), New Delhi MBL (NDM), Florence imipenemase (FIM), and recently Belém imipenemase (BIM) [7,8]. Among these, IMP- and VIM-type enzymes are the most abundant and are of particular clinical importance, as they efficiently inactivate most β-lactam antibiotics, with the exception of monobactams [9,10]. The first identification of IMP-1 metallo-β-lactamase was reported in 1988 in a P. aeruginosa strain isolated from Japan [11]. Since then, the emergence and spread of IMP-carrying P. aeruginosa has been reported globally, often associated with international high-risk clones such as ST111, ST233, and ST235 [12]. The IMP-type enzymes represent a highly heterogeneous group forming at least seven phylogenetic clusters and frequently found as gene cassettes in class 1 integrons [4,13,14].
Integrons play a crucial role in capturing and disseminating antibiotic resistance genes including MBLs [4] as they are often associated with large transposon structures found on plasmids or chromosomes [13,15]. Transposable elements utilize specific transposase-mediated mechanisms for their insertion and excision within the bacterial genome. Composite transposons are flanked by insertion sequence (IS) elements. Unit transposons, on the other hand, encode an excision/integration-associated enzyme, recombinase or resolvase, along with accessory genes such as resistance genes, within a single genetic unit. Conjugative transposons, also referred to as integrative conjugative elements (ICEs), carry genes for excision, conjugative transfer, and integration, often accommodating a diverse repertoire of accessory genes, including antibiotic resistance genes [16]. Resistance plasmids could be directly studied from genomic data by either replicon or MOB-typing schemes based on the diversity of replicative and relaxase proteins, respectively. MOB-typing is more sensitive and amenable to plasmids in Pseudomonas spp. [17].
P. aeruginosa isolates in Bulgaria display a range of carbapenemases, of which VIM-type enzymes are commonly reported [18,19]. The co-occurrence of NDM-1 and GES-5 carbapenemases was recently identified [20]. The detection of OXA-50 carbapenemase has also been documented, although currently it is being considered intrinsic in P. aeruginosa [21].
In this study, we performed genomic analysis of three clinical P. aeruginosa isolates, revealing to our knowledge the first occurrence of IMP-carrying P. aeruginosa in Bulgaria. Furthermore, among the isolates, there was one that harbored a novel variant of the IMP carbapenemase, officially denoted as IMP-100 upon its submission in the NCBI GenBank database. This variant was situated on a multidrug-resistant plasmid which we termed p4782-IMP and coexisted with a chromosomally encoded blaVIM-4 gene.
2. Materials and Methods
2.1. Strains
The three strains (Paer3541, Paer3796A, and Paer4782MK) were initially isolated in Sofia, Bulgaria, between 2018 and 2022. Paer3541 was obtained from the throat swab of a hospitalized patient, Paer3796A was derived from the urine sample of an individual in an outpatient setting, and Paer4782MK was isolated from the blood culture of a patient with acute myeloid leukemia. Strains were sent to the National Reference Laboratory, Department of Microbiology, within the National Center of Infectious and Parasitic Diseases, Sofia, Bulgaria, for carbapenem resistance confirmation purposes (Supplementary Table S1). These three strains were the only IMP-type carbapenemase-carrying P. aeruginosa strains to date.
2.2. Phenotypic and Molecular Analysis
Strains were cultured on Columbia agar (Diachim AD, Sofia, Bulgaria) at 35 °C overnight. A single colony of each strain was identified via MALDI Biotyper (Bruker Daltonics GmbH & Co. KG, Bremen, Germany) with MALDI Reference 2022 Library v4.0.0. Antimicrobial Susceptibility Testing (AST) was performed using MICRONAUT-S Pseudomonas MIC and UMIC® Cefiderocol assay (Bruker Daltonics GmbH & Co. KG, Bremen, Germany). The interpretation of the AST results was in accordance with EUCAST clinical breakpoints v13.0.
Initially, the isolates were analyzed for carbapenemase activity by a modified CarbaNP test [22]. In parallel, an in-house carbapenemase gene detection multiplex PCR was performed with the PCR components, conditions, and protocols described in detail in Supplementary Table S1. Primer pairs for each gene were obtained from previously published sources, including SIM, SPM, OXA-48-like, GES, and KPC [23,24,25,26] along with additional primer pairs for IMP, VIM, and NDM [27]. QIAxcel Advanced high-resolution capillary electrophoresis system was used following the PCR (Qiagen, Hilden, Germany) with protocol 0M800 (3 kV for 800 s) for precise size estimation.
The antimicrobial resistance gene expression assay of AMR-associated genes mexA, mexC, mexE, mexX, ampC, and oprD was performed as previously described [5,28], and results interpreted according to [29]. Biofilm formation was quantified by the modified crystal violet (CV) assay [30].
2.3. Genomic and Plasmid DNA Extraction and Whole Genome Sequencing (WGS)
Total genomic DNA extraction was performed with PureLink™ Genomic DNA Mini Kit (Thermo Fisher Scientific, Missouri, TX, USA) according to the manufacturer’s instructions except that all homogenization steps were carried out by pipetting. Plasmid DNA was acquired using NucleoSpin Plasmid Mini kit for plasmid DNA (Macherey-Nagel, Düren, Germany) following the “low-copy plasmid” protocol. Short-read next-generation sequencing (NGS) was performed using Illumina DNA Prep kit for sequencing libraries preparation and MiSeq V3 (2 × 300 bp) for strains Paer3541 and Paer3796A, whereas Paer4782MK was sequenced on NextSeq 550 with V2.5 (2 × 150 bp) mid output flow cell (Illumina, Inc., San Diego, CA, USA). The same DNA extract without additional size-selection was used for long-read sequencing on a MinION Mk1C with the Rapid Barcoding Kit 96 (SQK-RBK110.96) and FLO-MIN106D (R9.4.1) (Oxford Nanopore Technologies, Oxford, UK). The final purification step of the library pool was performed with 0.4x SPRI magnetic particles as recently suggested for removal of DNA fragments <1.5 kb [31].
2.4. Cloning, Transformation, and Conjugation/Mating Experiments
An 1150 bp fragment amplified with primers IMP-new_Clon_F and R comprising the complete IMP-100 (OR004774) open reading frame (ORF), its promotor region, and part of the IntI gene was cloned into the pET28a T7pCONS TIR-2 sfGFP vector (cat. No 154464, Addgene, Inc., Watertown, MA, USA) using the FastCloning protocol [32]. The insert amplification and vector linearization PCR are described in detail in Supplementary Table S2. A schematic representation of the cloning design is available in Supplementary Figure S1. The PCR products mixed at 1:1 ratio were digested with DpnI for 2 h at 37 °C.
Plasmid DNA from the Paer4782MK strain was transformed into P. aeruginosa PA01RifR. The transformation protocols for both the cloned IMP-100 and the whole p4782-IMP plasmid preparation were identical [33]. Briefly, E. coli NEB-10 (New England Biolabs, Inc., Ipswich, MA, USA) or P. aeruginosa PA01RifR recipients were incubated on Columbia agar at 35 °C overnight. A few colonies were inoculated into 10 mL BHI broth and agitated at 200 rpm at 35 °C until log phase (0.6 ± 0.05 OD) was achieved. Then, 1.4 mL suspension was aliquoted into 1.5 mL tubes and centrifuged at 8000× g for 2 min and the supernatant was discarded. Cell pellets were washed with 1 mL RT molecular-grade water twice by gentle resuspension. Cell pellets were resuspended in 80 µL of RT molecular-grade water. The digested vector and insert mix (20 µL) or plasmid DNA were added, gently mixed, and immediately transferred into 2 mm gap electroporation cuvettes (Avantor, Inc., Radnor, PA, USA). The electroporation was performed at 2500 mV on an Eporator® (Eppendorf, Hamburg, Germany). Immediately, 1 mL of BHI broth (RT) was added, gently mixed, and incubated at 35 °C for 2 h. Fresh BHI agar plates containing 50 mg/L kanamycin and 8 mg/L ceftazidime were streaked with 50 µL of the transformants.
The spontaneous transferability of plasmids carrying resistance determinants was investigated by filter mating [34] and the combined mating technique [35]. In an attempt to increase the conjugation efficiency in Pseudomonas, additional steps were implemented in the protocol as previously described [36]. Rifampicin-resistant E. coli C600 and a spontaneous mutant of P. aeruginosa PA01RifR were used as recipient strains. Transconjugants were selectively cultured on MacConkey agar supplemented with ceftazidime (30 mg/L) and rifampicin (200 mg/L) (Sigma Chemical Co., St Louis, MO, USA).
2.5. Bioinformatic Analysis
A quality check of both long and short reads was performed with FastQC v0.11.9 (https://www.bioinformatics.babraham.ac.uk/projects/fastqc, accessed on 8 June 2023). Quality trimming and filtering of the raw reads was conducted with fastp v0.23.2 [37] and filtlong v0.2.1 (https://github.com/rrwick/Filtlong, accessed on 8 June 2023) for short and long reads, respectively. First, long-read-only assemblies were produced with Flye v0.2.1 [38] and when no circular genome was achieved, hybrid assembly was attempted with Unicycler v0.4.8 [39]. The resulting assemblies were subsequently polished with tools such as Polypolish v0.5.0 [40], POLCA [41] and MEDAKA v1.7.3 (ONT, https://github.com/nanoporetech/medaka, accessed on 15 June 2023). Assembly quality was assessed by multiple tools such as Quast v 5.0.2 [42], BUSCO v5.4.6 [43], and CheckM v1.2.1 [44]. After genome quality evaluation, strain identification was performed with rMLST [45] and KmerFinder v 3.0.2 [46] with a database version from 11 July 2022. Then, genomes were annotated with Bakta v 1.7 [47] with database v5.0-full. Antibacterial resistance, phenotypic prediction, and virulence determinants were identified with AMRFinderPlus v3.11.4 [48] and ResFinder v4.3.1 [49] with database version 2022-05-24 and VFanalyzer (database version from 5 January 2023) [50], respectively. Mob-suite v 3.1.4 [17] was used for plasmid analysis. Analysis of ICEs was carried out with ICEfinder [51]. MLST profiles were inferred from sequence data using mlst v2.23.0 (Seemann T, mlst Github https://github.com/tseemann/mlst, accessed on 8 June 2023). In silico serotyping was performed using the tool PAst 1.0 distinguishing 11 serogroups by BLAST analysis of the O-specific antigen cluster. [52]. The functionality of the outer-membrane porin oprD was analyzed in silico using the recently described PorinPredict tool [53].
2.6. Data Availability
The complete genomes of the three strains were deposited in the European Nucleotide Archive (ENA) under project accession PRJEB62425. The genomes were assigned the following accession numbers: Paer4782MK—ERZ18545754, Paer3541—ERZ18545673, Paer3796A—ERZ18545648. In the case of Paer4782MK, a novel variant of IMP-100, a subclass B1 metallo-beta-lactamase, was detected, validated, and submitted to GenBank with accession number OR004774. The putative novel transposon was registered in The Transposon Registry (https://transposon.lstmed.ac.uk/, accessed on 28 August 2023) under Tn7700.
3. Results
3.1. AMR and Biofilm Formation
Antimicrobial susceptibility testing (AST) showed diverse resistance patterns among the tested strains. Paer4782MK exhibited a high level of resistance to multiple classes of antibiotics, including all beta-lactams except aztreonam, aminoglycosides, and fluoroquinolones, consistent with an XDR phenotype. Paer3541 demonstrated resistance to cephalosporins and fluroquinolones but remained susceptible to carbapenems and aminoglycosides (except tobramycin). Paer3796A displayed resistance to all tested cephalosporins, carbapenems (except imipenem/relebactam), fluoroquinolones (except levofloxacin), and aminoglycosides (except amikacin). However, all three strains remained susceptible to colistin despite resistance to novel cephalosporin/inhibitor combinations like ceftazidime/avibactam and ceftolozane/tazobactam. Notably, both the Paer4782MK and Paer3796A strains exhibited resistance to CFDC, a novel siderophore cephalosporin only recently approved for treatment of infections from MDR Gram-negatives [54]. The AST profiles of the three P. aeruginosa isolates are available in Table 1.

Table 1.
Minimal inhibitory concentrations (MIC) of P. aeruginosa strains in this study.
The modified CarbaNP test confirmed the presence of carbapenemases in all three isolates. Initially, the PCR results for IMP genes were negative. However, by employing alternative PCR primers [27], we successfully detected the presence of blaIMP in all three isolates and blaVIM in Paer4782MK.
We observed a positive correlation between the notable upregulation of the mexXY multidrug efflux operon in Paer3541 and the elevated MICs for levofloxacin and ciprofloxacin, and also between low to moderate mexXY expression and lower fluoroquinolone MIC values in Paer3796A. Despite Paer4782MK being characterized by the moderate expression of the mexCD, its association with the resistance profile remained uncertain as the strain already carried resistance genes for the majority of mexCD targets as seen from the WGS results. Interestingly, all three strains exhibited negative expression of the outer membrane porin oprD. The gene expression levels are available in Supplementary Table S3. Lastly, all strains exhibited strong biofilm production.
3.2. Genome Quality Assessment and Features
All three genomes achieved a completeness score close to 100%, indicating that the majority of the expected conserved genes were present. A high level of genome integrity without duplications was evidenced by the 99.2% single-copy genes detected. No fragmented or missing conserved genes were identified. In line with these findings, CheckM revealed relatively low foreign DNA contamination levels ranging from 0.63% to 0.7%. Detailed information on assembly statistics and genome quality assessment is available in Supplementary Table S4.
Two plasmids, p4782-IMP (61.5 Kbp) and p4782_002 (290.8 Kbp), were identified in Paer4782MK. The plasmid p4782-IMP (OX638703.1) was related to the MOBF and MPFT plasmid pMOS94-like family. Notably, this plasmid carried the novel blaIMP-100 (OR004774) allele, in addition to other genes associated with AMR. The plasmid p4782_002 (OX638702.1) was untypable and lacked both resistance and virulence determinants; thus it was excluded from further analysis. The two plasmids detected in Paer3541 were p3541_1 (179.3 Kbp, OX638611.1) and p3541_2 (41.5 Kbp, OX638612.1). Similar to p4782-IMP, the plasmid p3541_2 was MOBF and MPFT and was found to harbor a single aac(6′)-29 gene, encoding an aminoglycoside acetyltransferase. The plasmid p3541_1 had unknown MOB and MPF types, similar to p4782_002. For Paer3796A, a single plasmid p3796A (178.5 Kbp, OX638565.1) with unknown MOB and MPF types was identified and did not harbor any resistance or virulence determinants. Additional information regarding plasmid analysis is available in Supplementary Table S5.
Paer3541 and Paer3796A were assigned to ST621, a clone previously associated with IMP carriage [55], while Paer4782MK belonged to the international high-risk clone ST233. In silico serotyping revealed that Paer3541 and Paer3796A were serogroup O4, while Paer4782MK was O11.
3.3. Detection of AMR Determinants
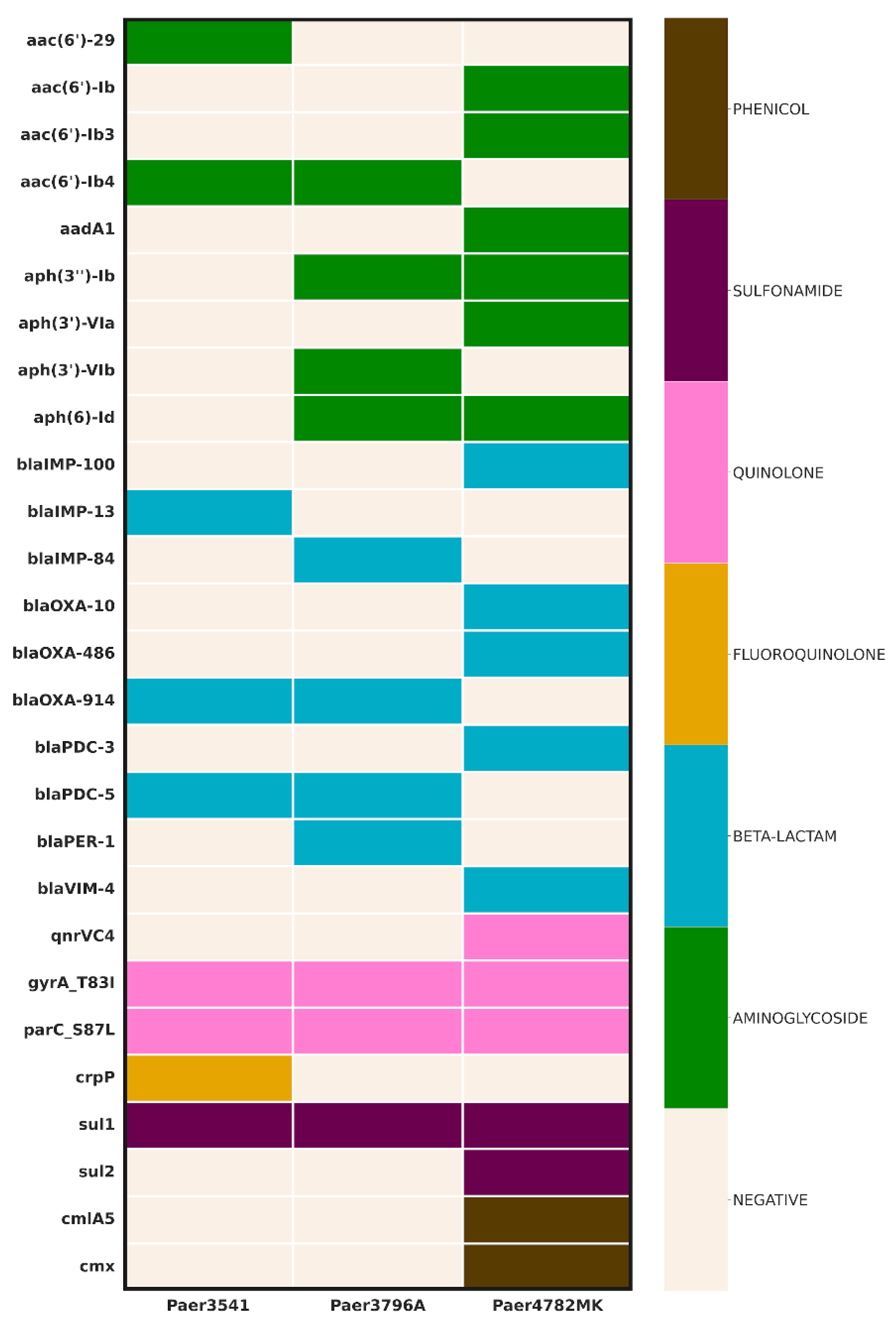
The AMR screening uncovered a diverse array of genes (Figure 1). All three isolates demonstrated the presence of an IMP carbapenemase, with the unique IMP-100 allele identified in a pMOS94-like plasmid in Paer4782MK. Additionally, strain Paer4782MK also harbored blaVIM-4. Two other closely related chromosomal IMP variants, IMP-13 and IMP-84, were detected in Paer3541 and Paer3796A, respectively. Additional acquired extended-spectrum beta-lactamases (ESBL) such as PER-1 and multiple genetic determinants associated with resistance to most non-beta-lactam antimicrobial groups were present as well.
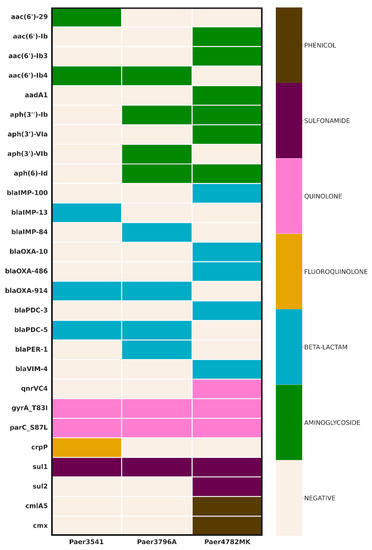
Figure 1.
Genomic inference of resistance determinants. Columns represent strains, whereas AMR genes are in rows. The color scheme represents the affected antimicrobial class (right) in relation to the resistance genes (left).
In silico analysis of the oprD gene revealed that both Paer3541 and Paer3796A harbor an intact and functional oprD porin. Despite the identification of a missense mutation (S325F) in the oprD gene of Paer3796A, the gene appeared intact. Conversely, a truncated oprD protein was evident in Paer4782MK, potentially compromising its function (Supplementary Table S6). No known mutations related to efflux overexpression were detected.
3.4. Detection of Virulence Determinants
All three isolates exhibited virulence factors related to adherence and motility, including flagella, type IV pili biosynthesis, and twitching motility. Alginate biosynthesis genes known for promoting bacterial resilience against harsh conditions and host immune responses [56] were detected in all isolates. However, certain antimicrobial-related factors (phzC2, phzD2, phzE2, phzF2, and phzG2) were absent from all strains compared to the reference strain PA01. Phospholipases C and the hemolytic plcH [57] were identified in all strains, while phospholidase D was absent from Paer4782MK. Complete siderophore biosynthesis operons were present in all strains, with the exception of the pyoverdine production gene pvdD, which was absent from Paer4782MK, indicating impaired pyoverdine synthesis [58]. Additional virulence factors, including alkaline protease (AprA), elastase (LasA and LasB), protease IV (PrpL), and exotoxin A (ToxA), as well as quorum-sensing components such as acylhomoserine lactone synthase (HdtS), transcription factors (LasI, RhlI), and receptors (LasR, RhlR), as well as the GacS/GacA two-component system and type VI secretion system (H-T6SS) components, were found in all three isolates. The rhamnolipid biosynthesis operon rhl associated with host cell infiltration and biofilm formation was also present in the studied strains [59,60]. T3SS effectors, including exoS, exoT, and exoY, involved in manipulating host cell signaling pathways and immune responses, along with T3SS genes such as pcrV, popB, popD, and pscF, were common across all our strains indicating a functional T3SS machinery capable of delivering effector molecules into host cells. All isolates harbored the hydrogen cyanide operon (HcnABC), a potent toxic compound known to disrupt cellular processes and impact aerobic respiration [61]. Furthermore, katG, involved in oxidative stress protection [62], was found exclusively on the p4782-IMP in Paer4782MK. A summary of the virulence screening results can be found in Supplementary Table S7.
3.5. Plasmid Analysis
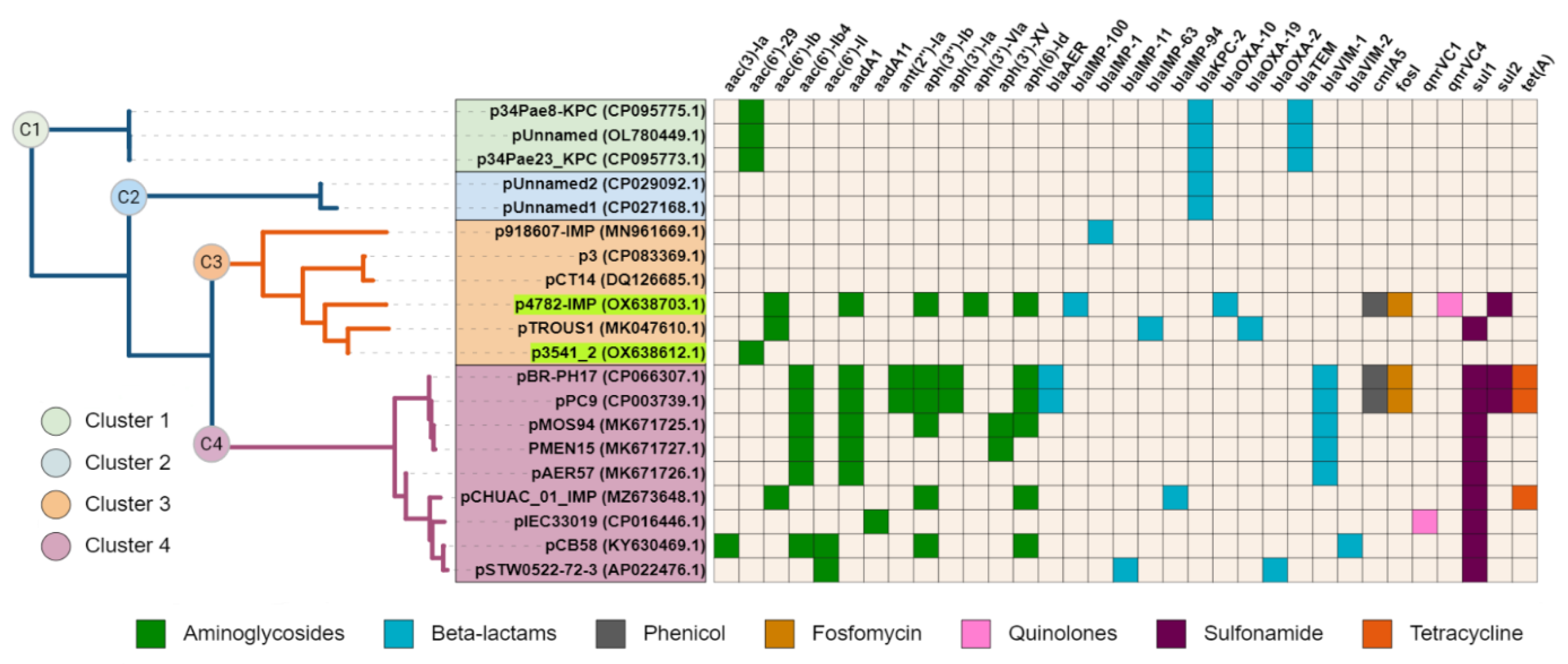
The complete sequence of plasmid p4782-IMP was subjected to a BLAST search against the nr/nt database of NCBI, and a neighbor-joining tree was constructed with the most similar plasmids (n = 19). Interestingly, plasmid p3541_2 was identified as closely related to p4782-IMP. Additionally, all plasmids were screened for AMR genes and the results were illustrated as a heatmap and linked to the phylogenetic tree (Figure 2).
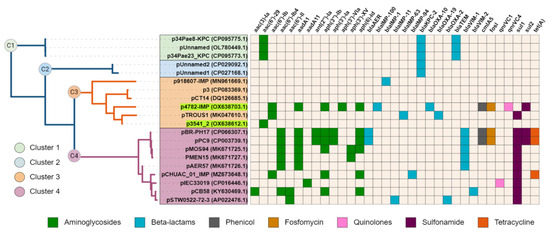
Figure 2.
Phylogenetic tree and resistance heatmap. Plasmids from the current study are highlighted in green. The different tree clusters (C1 ÷ C4) are distinguished by varying colors. The heatmap legend below provides information on the antibiotic class of the detected genes. The tree was generated with neighbor-joining after the NCBI blast with Max Seq Difference score ≥0.75. The heatmap and the tree were visualized with iToL 6.8 (https://itol.embl.de/about.cgi, accessed on 25 July 2023).
Upon observation, cluster C1 was found to encompass three plasmids with unknown MOB type that encoded KPC enzymes. The remaining clusters C2–C4 comprised plasmids from the MOBF and MPFT families. C2 was similar to C3 plasmids, but unlike them carried only the KPC enzyme. C3 and C4 both exhibited relatedness to the pMOS94-like plasmids [63]. Interestingly, C3 plasmids had lower density of AMR-associated genes compared to C4. Despite the high abundance of AMR genes found on plasmid p4782-IMP similar to C4 plasmids, it was grouped within the lower-resistance cluster C3 due to the larger degree of similarity of the backbone genes to C3.
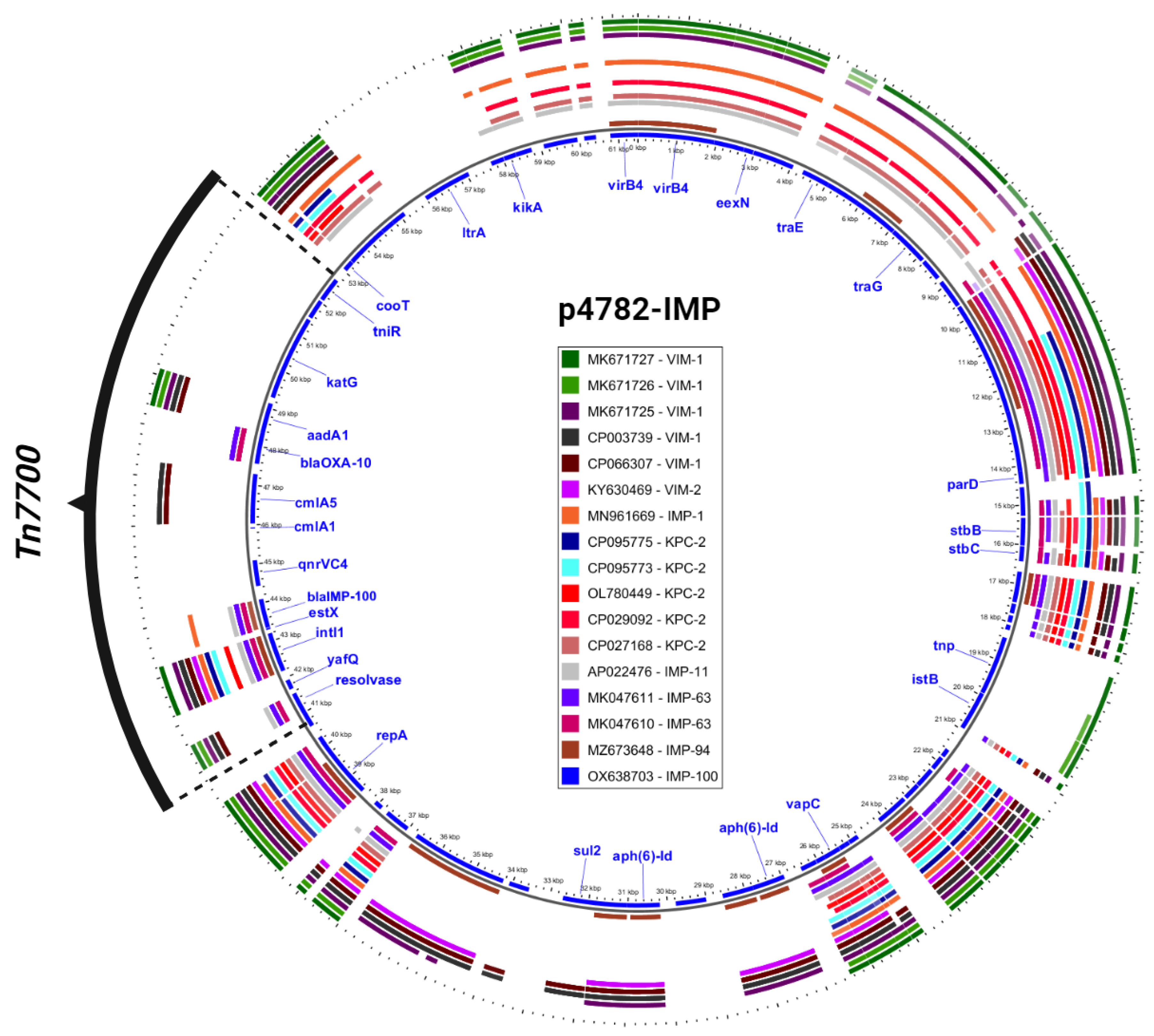
Additionally, an in-depth comparison was conducted between the most phylogenetically related carbapenemase-harboring plasmids and p4782-IMP using the Gview server’s BLAST Atlas. The analysis unveiled a distinctive transposon insertion within the MOBF/MPFT family plasmid backbone, as illustrated in Figure 3. This novel transposon carried the novel blaIMP-100 allele, as well as multiple AMR determinants, which accounted for the atypical abundance of resistance genes observed in p4782-IMP.
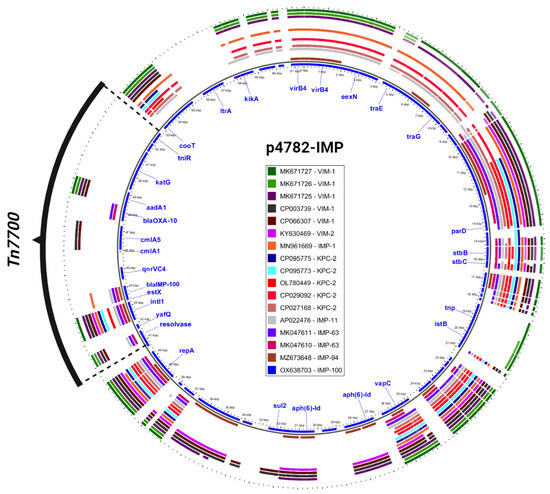
Figure 3.
Closest plasmids with carbapenemases. The plasmid accession numbers are listed in the legend and colored differently. The most inner circle (blue) is p4782-IMP which serves as a reference for comparison. Only important gene annotations were visualized in blue text. The novel transposon (Tn7700) is shown in black. The figure was produced with the Gview Atlas server (https://server.gview.ca, accessed on 25 July 2023).
3.6. Cloning, Transformation, and Conjugation
The transformants (PA01-p4782-IMP and E. coli NEB10-IMP-100) were successfully obtained. AST was conducted on both the recipients (PA01RifR and E. coli NEB10) and the transformants. The results are provided in Table 2. Firstly, the introduction of the whole p4782-IMP into PA01 increased the MIC for imipenem at least eight-fold, and for meropenem at least 128-fold. In comparison, the IMP-100 gene alone in E. coli NEB10-IMP-100 increased the MIC for imipenem at least two-fold, and for meropenem at least sixteen-fold. Although in silico analysis with mob-suite suggested the transferability of the MDR plasmid, all mating experiments were unsuccessful for both E. coli NEB10 and PA01 despite numerous attempts and variations.
Table 2.
AST of P. aeruginosa and E. coli transformants.
3.7. Phylogeny and Genetic Environment of blaIMP-100 in Paer4782MK
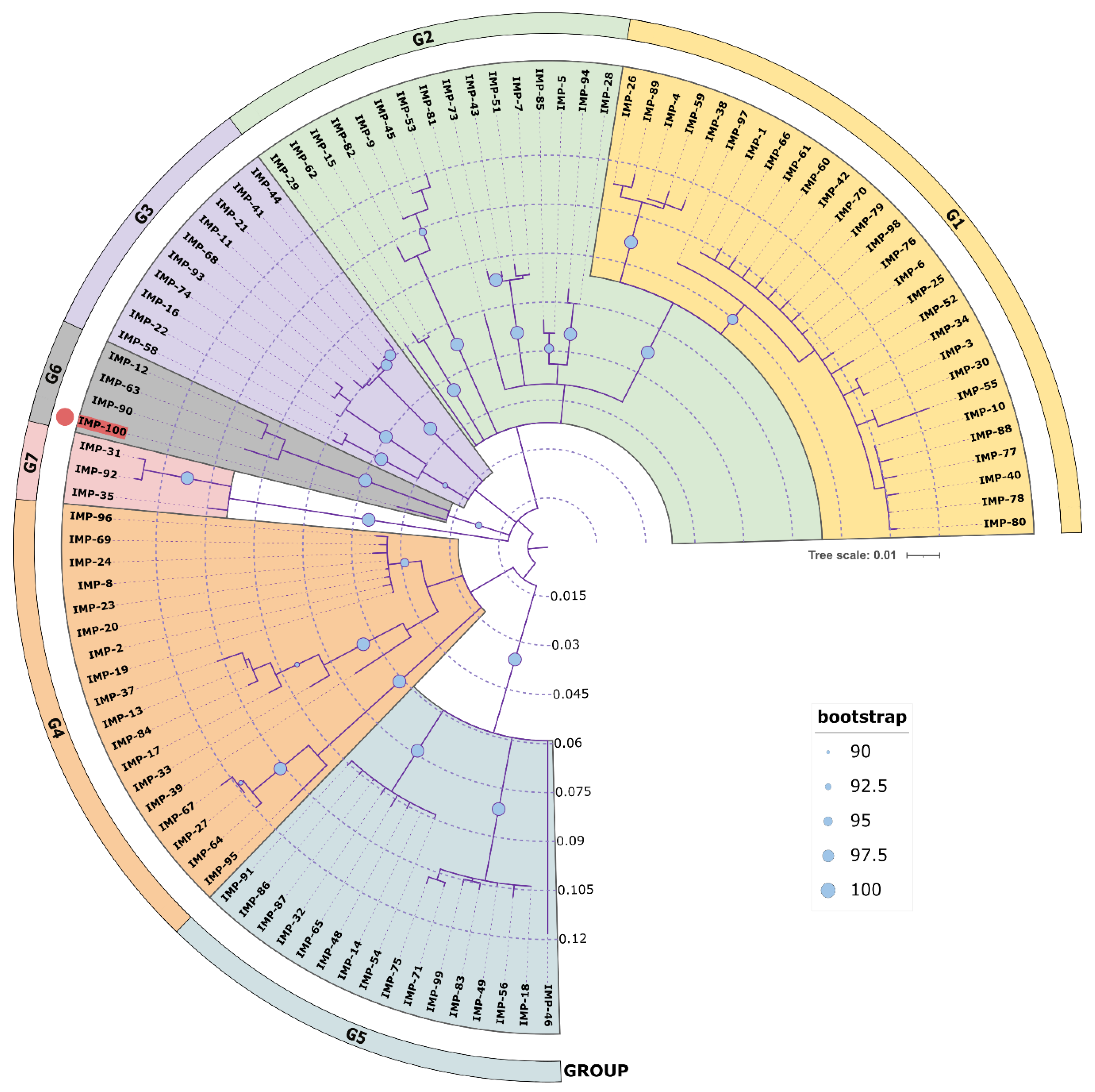
A phylogenetic analysis of all IMP variants was conducted through protein sequence alignment and subsequent construction of a phylogenetic tree (Figure 4). The clustering of IMP alleles and the labeling of the resulting groups were completed according to a clustering scheme suggested previously [14], thus maintaining consistency. Currently, there are 95 alleles including IMP-100. The novel allele IMP-100 falls into the G6 cluster revealing closest similarity to IMP-63, IMP-12, and IMP-90. The blaIMP-100 gene was positioned as the first cassette under a weak PcW+P2 promotor combination [64] in a novel In1300-like Tn402-type integron platform referred to as In4886 and followed by qnrVC4, cmlA5, and blaOXA-10 cassettes (Figure 5).
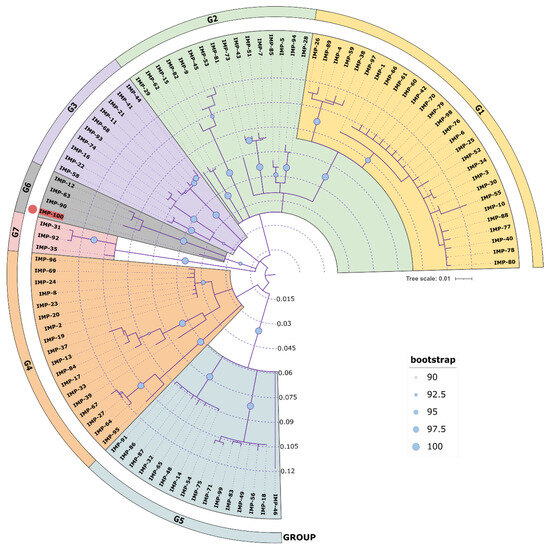
Figure 4.
Phylogenetic tree of all available IMP variants. All NCBI available protein sequences were downloaded and aligned with Muscle alignment 3.8.425 in Geneious Prime 2022 and the phylogenetic tree was constructed with Geneious Tree builder with the Jukes–Cantor genetic distance model and the neighbor-joining build method with bootstrap 1000. The tree was visualized in iToL 6.8 (https://itol.embl.de/about.cgi, accessed on 25 July 2023). Cluster groups G1 ÷ G7 were colored differently in accordance to [14], and the new IMP-100 allele is colored in red with a red dot at the tip of the leaf. Bootstrap indices are represented as circles placed at each node ranging in size from 90 (small circle) to 100 (large circle) to show the degree of cluster consistency.
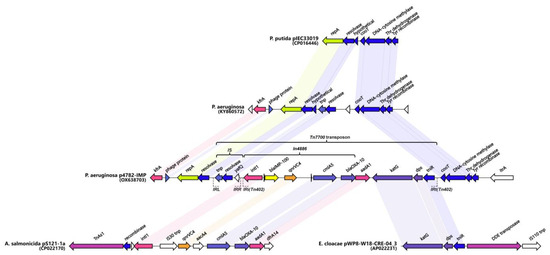
Figure 5.
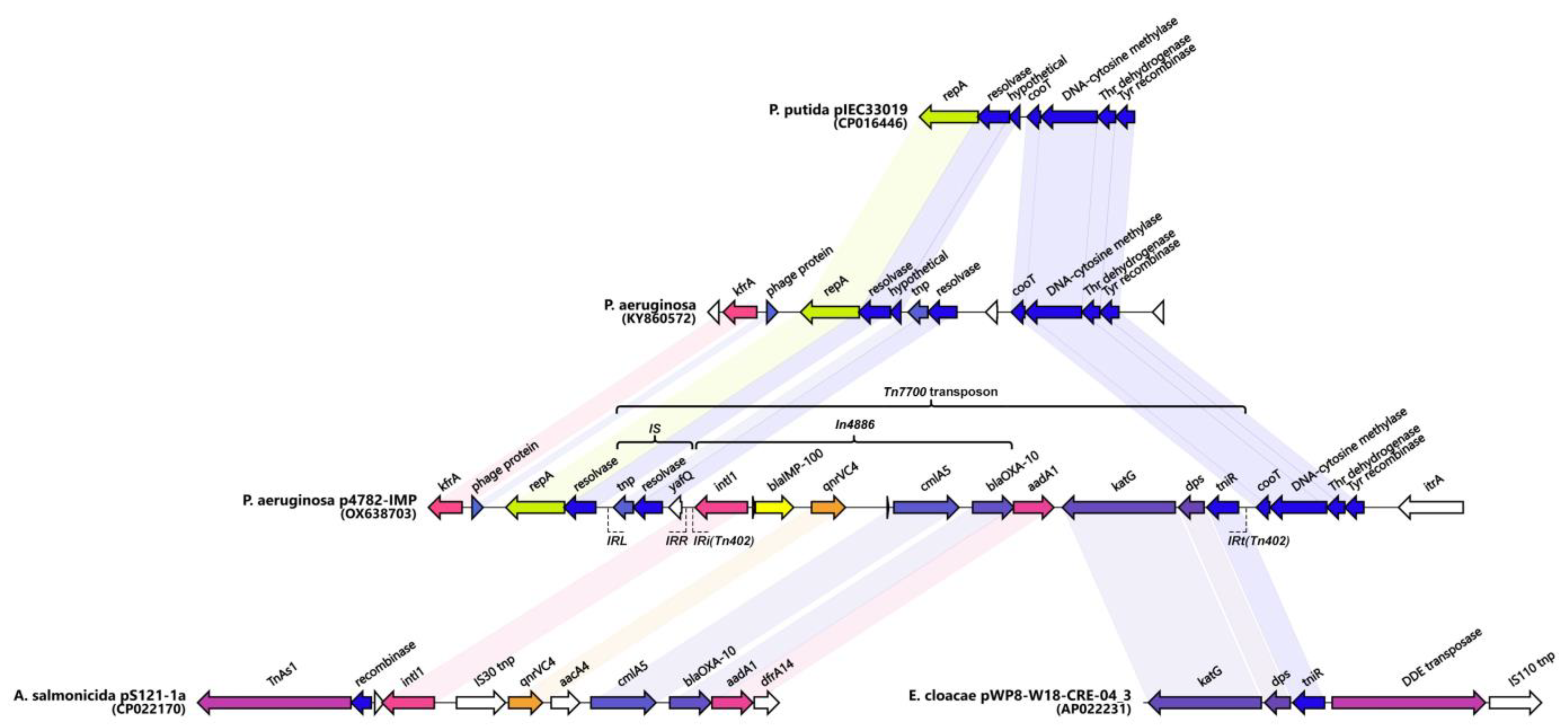
Genetic environment of blaIMP-100. A hypothesized origin of the novel transposon Tn7700 is depicted. Matching genes from the different sources are connected and identically colored. Non-matching genes (either hypothetical proteins or not) are shown in white. Unlabeled genes are hypothetical. The figure was created with Clinker v0.0.28 (https://github.com/gamcil/clinker, accessed on 25 July 2023).
A similar cassette array (In1300) was previously reported in the Aeromonas salmonicida MDR plasmid pS121-1a (CP022170). The In4886 was preceded by a novel putative IS, hereby referred to as IS4782 flanked by imperfect 25bp inverted repeats IRL (TGTCATTTTCAGAAGGCGACTGCAC) and IRR (TGTCATTTTCAGAAGACGACTGCAC) closely resembling those of ISPa17 and Tn402. The IRR was also found on the far 3′ end of the Tn402 integron as well (Figure 4). The IS4782 harbored three genes: the yafQ toxin gene, putative resolvase (188AA), and short transposase (128AA). This putative IS was lacking any significant homologs within the ISFinder or TnCentral databases [65,66]. The BLAST search was able to find eight identical hits (all plasmid-borne and one being the p3541_2 plasmid reported here) only matching the resolvase and transposase genes and not the whole IS. It was hypothesized that this element is capable of horizontal dissemination either alone or with the Tn402 integron due to sharing the same inverted repeats. Next, we managed to identify and map related mobile elements in order to detect a hypothetical origin of the putative novel transposon, registered as Tn7700 (Figure 5). The 5′ end most closely resembled the tnp/resolvase combination found in a P. aeruginosa transposon from the Czech Republic (KY860572) [67] followed by the In4886 carrying IMP-100 that was highly similar to the one from the Aeromonas salmonicida MDR plasmid pS121-1a (Figure 5). The 3′ end of the transposon resembled a remnant from another one found in the Enterobacter cloaceae plasmid (AP022231) containing katG, dps, and tniR. Similar structures have been reported in most pMOS94-like plasmids where ISPa17 preceded carbapenemase carrying Tn402 integrons [63]. Recently it was hypothesized that the ISPa17 transposase might be capable of the mobilization of adjacent Tn402 integrons resulting in distinct transposon elements [68]. The Tn7700 transposon had 5bp direct repeats (AAAAC) located up to 26bp apart from each IR, which although unexpected might be suggestive of past transposition events. Finally, we also discovered a probable ancestral insertion site of the whole Tn7700 located in a P. putida plasmid from Brazil (CP016446) [8].
For the remaining two strains we found that IMP-13 (Paer3541) and IMP-84 (Paer3796A) were located chromosomally in class 1 integrons embedded in Tn5051-like transposons, whereas VIM-4 (in Paer4782MK) was found in the In237-like integron. Importantly, all these MBLs were components of self-transmissible T4SS-type ICEs meaning they are capable of dissemination by horizontal gene transfer (Supplementary Figure S2. Lastly, all putative novel mobile genetic elements were submitted to the respective databases (ISFinder, TnRegistry, and INTEGRALL) [65,69,70].
4. Discussion
Between 2018 and 2022, three epidemiologically unrelated MDR P. aeruginosa strains were obtained from diverse clinical sources. Surprisingly, the modified CarbaNP test and the PCR for carbapenemase detection yielded conflicting results for strains Paer3541 and Paer3796A with the PCR being negative. Furthermore, WGS analysis revealed the presence of blaIMP-13 in Paer3541 and blaIMP-84 in Paer3796A. These findings were suggestive that the primers used to target blaIMP genes were ineffective and had to be replaced. Paer4782MK was the most recent isolate of the three and the blaIMP was therefore successfully detected with the updated PCR assay [27]. It is important to note that the availability of several alternative methods for carbapenemase detection may prove useful as well as regular updates of the methodologies being required to correctly detect the presence of rare gene variants.
Among the carbapenemases detected in Bulgaria, VIM-type enzymes are frequently observed [18,19], but occurrences of the NDM-1, GES-5, and OXA-50 carbapenemases have also been documented [20,21]. To our knowledge, this study represents the first report of blaIMP detection in three clinical MDR isolates from Bulgaria. Additionally, we identified a novel allele of the blaIMP gene, designated blaIMP-100, located on a MDR plasmid (p4782-IMP) within Paer4782MK. This strain also carried a chromosomally-encoded blaVIM-4 carbapenemase and was categorized as ST233, a globally prevalent multidrug-resistant clone [71], often associated with blaVIM-2 production [72]. Paer3541 (IMP-13) and Paer3796A (IMP-84) were classified as ST621, an epidemic clone known for its association with blaIMP production and a higher prevalence of pldA, a trans-kingdom phospholipase T6SS effector [73]. This effector is associated with the H2 Type VI secretion system (H2-T6SS) and is involved in bacterial endocytosis [74].
In line with previous research demonstrating a correlation between ST233 and the exoS+ (exoU-) genotype [72], our virulence analysis revealed that Paer4782MK (ST233) exhibited this specific genotype. ExoU and ExoS are mutually exclusive T3SS effectors, with ExoS leading to delayed apoptotic cell death, while ExoU induces rapid host cell lysis [75]. Regarding serotypes, our findings align with existing literature [76], demonstrating a strong association between serotypes O4 and O11 and MDR phenotypes. Typically, the O4 serotype is associated with the exoU-negative genotype, while the O11 serotype is linked to the exoU-positive genotype. Interestingly, Paer4782MK tested negative for exoU and exhibited the exoS genotype. Furthermore, despite recent studies suggesting an association between ST233 and serotype O6 [77], our classification of Paer4782MK as ST233:O11 deviates from this anticipated relationship.
Upon analyzing the WGS data, it was determined that the observed negative expression of oprD in the expression assay was due to an identical 10 amino acid indel, which prevented primer binding across all three isolates, thereby rendering the results invalid. Additionally, the PorinPredict tool confirmed the presence of an intact and functional oprD porin in Paer3541, which likely contributed to its susceptibility to and the lowest MIC values of meropenem, imipenem, meropenem/vaborbactam, and imipenem/relebactam for all three strains. Despite the occurrence of a missense mutation S325F, the oprD integrity in Paer3796A remained unaffected. In fact, the presence of this mutation has been associated with a potential increase in the MIC of imipenem [78]. However, further investigations are necessary to determine its precise impact. Lastly, both our observation and PorinPredict concluded that the oprD gene in Paer4782MK is truncated, and therefore its functionality is compromised, consistent with the higher reported MIC values for meropenem, imipenem, meropenem/vaborbactam, and imipenem/relebactam in this particular strain.
Phylogenetic analysis demonstrated a close genetic association between the plasmid p4782-IMP, harboring the novel blaIMP-100 allele, and the pMOS94-like plasmid family. The pMOS94 plasmid family was recently recognized as an emerging lineage involved in the dissemination of MBL genes among Pseudomonas species, also commonly carrying blaVIM and blaIMP genes, and recently blaBIM, and some of them were found to display disruptions in the transfer module (trw), thus impeding conjugation [63]. In the genetic environment of the trw transfer module of p4782-IMP, we could not identify any disruption, so the observed unsuccessful mating experiments could be due to other reasons. Next, the comparison with the nearest carbapenemase-harboring plasmids revealed an insertion of the unique Tn7700 transposon in p4782-IMP, carrying multiple AMR determinants. The transformation of PA01 with the p4782-IMP plasmid resulted in a resistance profile similar to that of the Paer4782MK strain, demonstrating the impact of p4782-IMP in reducing susceptibility to important antibiotic classes and conferring multiple drug resistance. Furthermore, the transfer of the entire plasmid into the PA01 conferred resistance to CFDC. On the other hand, in the case of the E. coli NEB10 transformant carrying only the blaIMP-100 gene, there was no significant increase in the CFDC MIC suggesting that the CFDC resistance was a result of the complex interplay between multiple genes present on the plasmid, in addition to blaIMP-100.
5. Conclusions
Our study is the first to our knowledge to document the blaIMP in clinical P. aeruginosa from Bulgaria. We also discovered a novel blaIMP-100 allele located on a MDR plasmid (p4782-IMP). The plasmid in turn conferred resistance to multiple antibiotics including CFDC, as shown by the similar MDR pattern revealed in the PA01 transformant. However, by examining transformants carrying only the blaIMP-100 gene, we observed that the CFDC MIC values did not show a substantial increase, further implying the involvement of other plasmid genes in the development of resistance to this novel agent. Lastly, our study emphasizes the importance of employing multiple methods for carbapenemase screening, given that conventional methods, apart from WGS, might show reduced sensitivity and neglect carbapenemase activity or genes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms11092270/s1, Table S1. Carba mPCR; Table S2. Cloning protocol; Figure S1. Cloning schematic; Table S3. Expression analysis; Table S4. Genome Quality; Table S5. MOB Suite results; Table S6. oprD analysis; Table S7. Vir. factors & BF; Figure S2. ICEs.
Author Contributions
All authors contributed in different aspects. Conceptualization, methodology, and validation—I.N.I., R.H., S.S. and I.S; software, formal analysis, and investigation—I.S., D.D., I.N.I., D.T. and E.D.; resources—S.S., E.D. and R.H.; data curation—D.D., D.T. and S.S.; writing—original draft preparation—I.S., I.N.I. and D.D.; writing—review and editing—I.S., I.N.I., S.S., D.T. and E.D.; visualization—D.D. and I.S.; supervision, project administration, and funding acquisition—I.N.I. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by The Bulgarian National Science Fund under Grant KП-06-H23/5 and the European Regional Development Fund through the Operational Program Science and Education for Smart Growth 2014–2020; Grant BG05M2OP001-1.002-0001-C04 “Fundamental Translational and Clinical Research in Infection and Immunity”.
Data Availability Statement
All used data are included in the main text and in the supplementary materials. Relevant links and/or references to other sources are included in the main text. The generated information and/or datasets analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Daikos, G.L.; da Cunha, C.A.; Rossolini, G.M.; Stone, G.G.; Baillon-Plot, N.; Tawadrous, M.; Irani, P. Review of Ceftazidime-Avibactam for the Treatment of Infections Caused by Pseudomonas aeruginosa. Antibiotics 2021, 10, 1126. [Google Scholar] [CrossRef]
- Toleman, M.A.; Biedenbach, D.; Bennett, D.M.C.; Jones, R.N.; Walsh, T.R. Italian metallo-β-lactamases: A national problem? Report from the SENTRY Antimicrobial Surveillance Programme. J. Antimicrob. Chemother. 2005, 55, 61–70. [Google Scholar] [CrossRef]
- Zavascki, A.P.; Barth, A.L.; Gonçalves, A.L.S.; Moro, A.L.D.; Fernandes, J.F.; Martins, A.F.; Ramos, F.; Goldani, L.Z. The influence of metallo- -lactamase production on mortality in nosocomial Pseudomonas aeruginosa infections. J. Antimicrob. Chemother. 2006, 58, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R.; Toleman, M.A.; Poirel, L.; Nordmann, P. Metallo-β-lactamases: The quiet before the storm? Clin. Microbiol. Rev. 2005, 18, 306–325. [Google Scholar] [CrossRef] [PubMed]
- Quale, J.; Bratu, S.; Gupta, J.; Landman, D. Interplay of Efflux System, ampC, and oprD Expression in Carbapenem Resistance of Pseudomonas aeruginosa Clinical Isolates. Antimicrob. Agents Chemother. 2006, 50, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, J.-M.; Poirel, L.; Nordmann, P. Molecular Epidemiology and Mechanisms of Carbapenem Resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2009, 53, 4783–4788. [Google Scholar] [CrossRef]
- Hong, D.J.; Bae, I.K.; Jang, I.-H.; Jeong, S.H.; Kang, H.-K.; Lee, K. Epidemiology and Characteristics of Metallo-β-Lactamase-Producing Pseudomonas aeruginosa. Infect. Chemother. 2015, 47, 81–97. [Google Scholar] [CrossRef]
- Souza, C.O.; Cayô, R.; Lima, K.V.B.; Brasiliense, D.M.; Streling, A.P.; Siqueira, A.V.; Alberto-Lei, F.; Leal, J.T.; Nodari, C.S.; Pérez-Chaparro, P.J.; et al. Genetic and biochemical characterization of BIM-1, a novel acquired subgroup B1 MBL found in a Pseudomonas sp. strain from the Brazilian Amazon region. J. Antimicrob. Chemother. 2023, 78, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V. Metallo beta lactamases in Pseudomonas aeruginosa and Acinetobacter species’. Expert Opin. Investig. Drugs 2008, 17, 131–143. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. β-Lactamases identified in clinical isolates of Pseudomonas aeruginosa’. Crit. Rev. Microbiol. 2010, 36, 245–258. [Google Scholar] [CrossRef]
- Watanabe, M.; Iyobe, S.; Inoue, M.; Mitsuhashi, S. Transferable imipenem resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1991, 35, 147–151. [Google Scholar] [CrossRef]
- Wang, M.-G.; Liu, Z.-Y.; Liao, X.-P.; Sun, R.-Y.; Li, R.-B.; Liu, Y.; Fang, L.-X.; Sun, J.; Liu, Y.-H.; Zhang, R.-M. Retrospective Data Insight into the Global Distribution of Carbapenemase-Producing Pseudomonas aeruginosa. Antibiotics 2021, 10, 548. [Google Scholar] [CrossRef]
- Zhao, W.-H.; Hu, Z.-Q. IMP-type metallo-β-lactamases in Gram-negative bacilli: Distribution, phylogeny, and association with integrons. Crit. Rev. Microbiol. 2011, 37, 214–226. [Google Scholar] [CrossRef]
- Li, X.; Mu, X.; Chen, F.; Lu, X.; He, J.; Zheng, Y.; Zhou, D.; Yin, Z.; Wang, P. Characterization of Three Novel IMP Metallo-β-Lactamases, IMP-89, IMP-91, and IMP-96, and Diverse bla IMP -Carrying Accessory Genetic Elements from Chinese Clinical Isolates. Microbiol. Spectr. 2023, 11, e0498622. [Google Scholar] [CrossRef]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Genet. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.P.; Chandler, M.; Courvalin, P.; Guédon, G.; Mullany, P.; Pembroke, T.; Rood, J.I.; Smith, C.J.; Summers, A.O.; Tsuda, M.; et al. Revised Nomenclature for Transposable Genetic Elements. Plasmid 2008, 60, 167–173. [Google Scholar] [CrossRef]
- Robertson, J.; Nash, J.H.E. MOB-suite: Software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microb. Genom. 2018, 4, e000206. [Google Scholar] [CrossRef]
- Schneider, I.; Keuleyan, E.; Rasshofer, R.; Markovska, R.; Queenan, A.M.; Bauernfeind, A. VIM-15 and VIM-16, Two New VIM-2-Like Metallo-β-Lactamases in Pseudomonas aeruginosa Isolates from Bulgaria and Germany. Antimicrob. Agents Chemother. 2008, 52, 2977–2979. [Google Scholar] [CrossRef]
- Strateva, T.; Setchanova, L.; Peykov, S. Characterization of a Bulgarian VIM-2 metallo-β-lactamase-producing Pseudomonas aeruginosa clinical isolate belonging to the high-risk sequence type 111. Infect. Dis. 2021, 53, 883–887. [Google Scholar] [CrossRef]
- Kostyanev, T.; Nguyen, M.; Markovska, R.; Stankova, P.; Xavier, B.; Lammens, C.; Marteva-Proevska, Y.; Velinov, T.; Cantón, R.; Goossens, H.; et al. Emergence of ST654 Pseudomonas aeruginosa co-harbouring blaNDM-1 and blaGES-5 in novel class I integron In1884 from Bulgaria. J. Glob. Antimicrob. Resist. 2020, 22, 672–673. [Google Scholar] [CrossRef]
- Petrova, A.; Feodorova, Y.; Miteva-Katrandzhieva, T.; Petrov, M.; Murdjeva, M. First detected OXA-50 carbapenem-resistant clinical isolates Pseudomonas aeruginosa from Bulgaria and interplay between the expression of main efflux pumps, OprD and intrinsic AmpC. J. Med. Microbiol. 2019, 68, 1723–1731. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Dortet, L. Rapid Detection of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 2012, 18, 1503–1507. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Naas, T.; Nordmann, P. Pyrosequencing as a rapid tool for identification of ges-type extended-spectrum β-lactamases. J. Clin. Microbiol. 2006, 44, 3008–3011. [Google Scholar] [CrossRef]
- Mendes, R.E.; Kiyota, K.A.; Monteiro, J.; Castanheira, M.; Andrade, S.S.; Gales, A.C.; Pignatari, A.C.C.; Tufik, S. Rapid Detection and Identification of Metallo-β-Lactamase-Encoding Genes by Multiplex Real-Time PCR Assay and Melt Curve Analysis. J. Clin. Microbiol. 2007, 45, 544–547. [Google Scholar] [CrossRef]
- Cole, J.M.; Schuetz, A.N.; Hill, C.E.; Nolte, F.S. Development and Evaluation of a Real-Time PCR Assay for Detection of Klebsiella pneumoniae Carbapenemase Genes. J. Clin. Microbiol. 2009, 47, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Gröbner, S.; Linke, D.; Schütz, W.; Fladerer, C.; Madlung, J.; Autenrieth, I.B.; Witte, W.; Pfeifer, Y. Emergence of carbapenem-non-susceptible extended-spectrum β-lactamase-producing Klebsiella pneumoniae isolates at the university hospital of Tübingen, Germany. J. Med. Microbiol. 2009, 58, 912–922. [Google Scholar] [CrossRef]
- Goudarzi, H.; Mirsamadi, E.S.; Ghalavand, Z.; Vala, M.H.; Mirjalali, H.; Hashemi, A. Rapid detection and molecular survey of blaVIM, blaIMP and blaNDM genes among clinical isolates of Acinetobacter baumannii using new multiplex real-time PCR and melting curve analysis. BMC Microbiol. 2019, 19, 122. [Google Scholar] [CrossRef]
- Wi, Y.M.; Greenwood-Quaintance, K.E.; Schuetz, A.N.; Ko, K.S.; Peck, K.R.; Song, J.-H.; Patel, R. Activity of Ceftolozane-Tazobactam against Carbapenem-Resistant, Non-Carbapenemase-Producing Pseudomonas aeruginosa and Associated Resistance Mechanisms. Antimicrob. Agents Chemother. 2018, 62, e01970-17. [Google Scholar] [CrossRef] [PubMed]
- Cabot, G.; Ocampo-Sosa, A.A.; Tubau, F.; Macia, M.D.; Rodríguez, C.; Moya, B.; Zamorano, L.; Suárez, C.; Peña, C.; Martínez-Martínez, L.; et al. Overexpression of AmpC and Efflux Pumps in Pseudomonas aeruginosa Isolates from Bloodstream Infections: Prevalence and Impact on Resistance in a Spanish Multicenter Study. Antimicrob. Agents Chemother. 2011, 55, 1906–1911. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Rao, T.S. An Improved Crystal Violet Assay for Biofilm Quantification in 96-Well Microtitre Plate. bioRxiv 2017, 100214. [Google Scholar] [CrossRef]
- Alvarez-Arevalo, M.; Sterndorff, E.B.; Faurdal, D.; Jørgensen, T.S.; Mourched, A.-S.; Vuksanovic, O.; Saha, S.; Weber, T. Extraction and Oxford Nanopore sequencing of genomic DNA from filamentous Actinobacteria. STAR Protoc. 2022, 4, 101955. [Google Scholar] [CrossRef]
- Li, C.; Wen, A.; Shen, B.; Lu, J.; Huang, Y.; Chang, Y. FastCloning: A highly simplified, purification-free, sequence- and ligation-independent PCR cloning method. BMC Biotechnol. 2011, 11, 92. [Google Scholar] [CrossRef]
- Tu, Q.; Yin, J.; Fu, J.; Herrmann, J.; Li, Y.; Yin, Y.; Stewart, A.F.; Müller, R.; Zhang, Y. Room temperature electrocompetent bacterial cells improve DNA transformation and recombineering efficiency. Sci. Rep. 2016, 6, 24648. [Google Scholar] [CrossRef]
- Livermore, D.M.; Jones, C.S. Characterization of NPS-1, a novel plasmid-mediated beta-lactamase, from two Pseudomonas aeruginosa isolates. Antimicrob. Agents Chemother. 1986, 29, 99–103. [Google Scholar] [CrossRef]
- Walter, M.V.; Porteous, A.; Seidler, R.J. Measuring genetic stability in bacteria of potential use in genetic engineering. Appl. Environ. Microbiol. 1987, 53, 105–109. [Google Scholar] [CrossRef]
- Sakuda, A.; Suzuki-Minakuchi, C.; Matsui, K.; Takahashi, Y.; Okada, K.; Yamane, H.; Shintani, M.; Nojiri, H. Divalent cations increase the conjugation efficiency of the incompatibility P-7 group plasmid pCAR1 among different Pseudomonas hosts. Microbiology 2018, 164, 20–27. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Wick, R.R.; Holt, K.E. Polypolish: Short-read polishing of long-read bacterial genome assemblies. PLoS Comput. Biol. 2022, 18, e1009802. [Google Scholar] [CrossRef]
- Zimin, A.V.; Salzberg, S.L. The genome polishing tool POLCA makes fast and accurate corrections in genome assemblies. PLoS Comput. Biol. 2020, 16, e1007981. [Google Scholar] [CrossRef] [PubMed]
- Mikheenko, A.; Prjibelski, A.; Saveliev, V.; Antipov, D.; Gurevich, A. Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 2018, 34, i142–i150. [Google Scholar] [CrossRef]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Simão, F.A.; Zdobnov, E.M. BUSCO Update: Novel and Streamlined Workflows along with Broader and Deeper Phylogenetic Coverage for Scoring of Eukaryotic, Prokaryotic, and Viral Genomes. Mol. Biol. Evol. 2021, 38, 4647–4654. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Bliss, C.M.; Bennett, J.S.; Bratcher, H.B.; Brehony, C.; Colles, F.M.; Wimalarathna, H.; Harrison, O.B.; Sheppard, S.K.; Cody, A.J.; et al. Ribosomal multilocus sequence typing: Universal characterization of bacteria from domain to strain. Microbiology 2012, 158 Pt 4, 1005–1015. [Google Scholar] [CrossRef]
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and precise alignment of raw reads against redundant databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef]
- Schwengers, O.; Jelonek, L.; Dieckmann, M.A.; Beyvers, S.; Blom, J.; Goesmann, A. Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 2021, 7, 000685. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci. Rep. 2021, 11, 307. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Zhou, S.; Chen, L.; Yang, J. VFDB 2022: A general classification scheme for bacterial virulence factors. Nucleic Acids Res. 2022, 50, D912–D917. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.-Y. ICEberg 2.0: An updated database of bacterial integrative and conjugative elements. Nucleic Acids Res. 2019, 47, D660–D665. [Google Scholar] [CrossRef]
- Thrane, S.W.; Taylor, V.L.; Lund, O.; Lam, J.S.; Jelsbak, L. Application of whole-genome sequencing data for o-specific antigen analysis and in silico serotyping of pseudomonas aeruginosa isolates. J. Clin. Microbiol. 2016, 54, 1782–1788. [Google Scholar] [CrossRef] [PubMed]
- Biggel, M.; Johler, S.; Roloff, T.; Tschudin-Sutter, S.; Bassetti, S.; Siegemund, M.; Egli, A.; Stephan, R.; Seth-Smith, H.M.B. PorinPredict: In Silico Identification of OprD Loss from WGS Data for Improved Genotype-Phenotype Predictions of P. aeruginosa Carbapenem Resistance. Microbiol. Spectr. 2023, 11, e0358822. [Google Scholar] [CrossRef]
- Syed, Y.Y. Cefiderocol: A Review in Serious Gram-Negative Bacterial Infections. Drugs 2021, 81, 1559–1571. [Google Scholar] [CrossRef] [PubMed]
- Giani, T.; Arena, F.; Pollini, S.; Di Pilato, V.; D’andrea, M.M.; De Angelis, L.H.; Bassetti, M.; Rossolini, G.M.; Vismara, C.; Luzzaro, F.; et al. Italian nationwide survey on Pseudomonas aeruginosa from invasive infections: Activity of ceftolozane/tazobactam and comparators, and molecular epidemiology of carbapenemase producers. J. Antimicrob. Chemother. 2018, 73, 664–671. [Google Scholar] [CrossRef]
- Franklin, M.J.; Nivens, D.E.; Weadge, J.T.; Howell, P.L. Biosynthesis of the Pseudomonas aeruginosa Extracellular Polysaccharides, Alginate, Pel, and Psl. Front. Microbiol. 2011, 2, 167. [Google Scholar] [CrossRef] [PubMed]
- Vasil, M.L.; Stonehouse, M.J.; Vasil, A.I.; Wadsworth, S.J.; Goldfine, H.; Bolcome, R.E.; Chan, J. A Complex Extracellular Sphingomyelinase of Pseudomonas aeruginosa Inhibits Angiogenesis by Selective Cytotoxicity to Endothelial Cells. PLoS Pathog. 2009, 5, e1000420. [Google Scholar] [CrossRef]
- Meldrum, A.J. Regulation of Pyoverdine Biosynthesis in Pseudomonas aeruginosa. Master’s Thesis, Queen’s University, Kingston, ON, Canada, 1999. Available online: https://www.nlc-bnc.ca/obj/s4/f2/dsk2/ftp01/MQ37969.pdf?is_thesis=1&oclc_number=1017515220 (accessed on 6 September 2023).
- McClure, C.D.; Schiller, N.L. Inhibition of macrophage phagocytosis by pseudomonas aeruginosa rhamnolipids in vitro and in vivo. Curr. Microbiol. 1996, 33, 109–117. [Google Scholar] [CrossRef]
- Zulianello, L.; Canard, C.; Köhler, T.; Caille, D.; Lacroix, J.-S.; Meda, P. Rhamnolipids Are Virulence Factors That Promote Early Infiltration of Primary Human Airway Epithelia by Pseudomonas aeruginosa. Infect. Immun. 2006, 74, 3134–3147. [Google Scholar] [CrossRef]
- Ryall, B.; Davies, J.C.; Wilson, R.; Shoemark, A.; Williams, H.D. Pseudomonas aeruginosa, cyanide accumulation and lung function in CF and non-CF bronchiectasis patients. Eur. Respir. J. 2008, 32, 740–747. [Google Scholar] [CrossRef]
- Manca, C.; Paul, S.; Barry, C.E.; Freedman, V.H.; Kaplan, G. Mycobacterium tuberculosis catalase and peroxidase activities and resistance to oxidative killing in human monocytes in vitro. Infect. Immun. 1999, 67, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Di Pilato, V.; Antonelli, A.; Giani, T.; De Angelis, L.H.; Rossolini, G.M.; Pollini, S. Identification of a novel plasmid lineage associated with the dissemination of metallo-β-lactamase genes among pseudomonads. Front. Microbiol. 2019, 10, 1504. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiang, X.; Li, M.; Chen, X.; Li, G.; Li, R.; Lu, Y. Transcription of integron-harboured gene cassette impacts integration efficiency in class 1 integron. Mol. Microbiol. 2011, 80, 1326–1336. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Ross, K.; Varani, A.M.; Snesrud, E.; Huang, H.; Alvarenga, D.O.; Zhang, J.; Wu, C.; McGann, P.; Chandler, M. TnCentral: A Prokaryotic Transposable Element Database and Web Portal for Transposon Analysis. Mbio 2021, 12, e0206021. [Google Scholar] [CrossRef] [PubMed]
- Papagiannitsis, C.C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J. Molecular Characterization of Carbapenemase-Producing Pseudomonas aeruginosa of Czech Origin and Evidence for Clonal Spread of Extensively Resistant Sequence Type 357 Expressing IMP-7 Metallo-β-Lactamase. Antimicrob. Agents Chemother. 2017, 61, e01811-17. [Google Scholar] [CrossRef] [PubMed]
- Papa-Ezdra, R.; Cordeiro, N.F.; Outeda, M.; Garcia-Fulgueiras, V.; Araújo, L.; Seija, V.; Ayala, J.A.; Bado, I.; Vignoli, R. Novel Resistance Regions Carrying TnaphA6, blaVIM-2, and blaPER-1, Embedded in an ISPa40-Derived Transposon from Two Multi-Resistant Pseudomonas aeruginosa Clinical Isolates. Antibiotics 2023, 12, 304. [Google Scholar] [CrossRef]
- Moura, A.; Soares, M.; Pereira, C.; Leitão, N.; Henriques, I.; Correia, A. INTEGRALL: A database and search engine for integrons, integrases and gene cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef]
- Tansirichaiya, S.; Rahman, M.A.; Roberts, A.P. The Transposon Registry. Mobile DNA 2019, 10, 40. [Google Scholar] [CrossRef]
- Aguilar-Rodea, P.; Zúñiga, G.; Rodríguez-Espino, B.A.; Cervantes, A.L.O.; Arroyo, A.E.G.; Moreno-Espinosa, S.; Zamboni, D.d.l.R.; Martínez, B.L.; Castellanos-Cruz, M.d.C.; Parra-Ortega, I.; et al. Identification of extensive drug resistant Pseudomonas aeruginosa strains: New clone ST1725 and high-risk clone ST233. PLoS ONE 2017, 12, e0172882. [Google Scholar] [CrossRef]
- del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired β-lactamases: 2020 update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef]
- Boulant, T.; Boudehen, Y.-M.; Filloux, A.; Plesiat, P.; Naas, T.; Dortet, L. Higher Prevalence of PldA, a Pseudomonas aeruginosa Trans-Kingdom H2-Type VI Secretion System Effector, in Clinical Isolates Responsible for Acute Infections and in Multidrug Resistant Strains. Front. Microbiol. 2018, 9, 2578. [Google Scholar] [CrossRef]
- Russell, A.B.; LeRoux, M.; Hathazi, K.; Agnello, D.M.; Ishikawa, T.; Wiggins, P.A.; Wai, S.N.; Mougous, J.D. Diverse type VI secretion phospholipases are functionally plastic antibacterial effectors. Nature 2013, 496, 508–512. [Google Scholar] [CrossRef]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Recio, R.; Mancheño, M.; Viedma, E.; Villa, J.; Orellana, M.; Lora-Tamayo, J.; Chaves, F. Predictors of Mortality in Bloodstream Infections Caused by Pseudomonas aeruginosa and Impact of Antimicrobial Resistance and Bacterial Virulence. Antimicrob. Agents Chemother. 2020, 64, e01759-19. [Google Scholar] [CrossRef]
- Pottier, M.; Gravey, F.; Castagnet, S.; Auzou, M.; Langlois, B.; Guérin, F.; Giard, J.-C.; Léon, A.; Le Hello, S. A 10-year microbiological study of Pseudomonas aeruginosa strains revealed the circulation of populations resistant to both carbapenems and quaternary ammonium compounds. Sci. Rep. 2023, 13, 2639. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, J.Y.; Maure, A.; Gingras, H.; Leprohon, P.; Ouellette, M. Chemogenomic Screen for Imipenem Resistance in Gram-Negative Bacteria. mSystems 2019, 4, e00465-19. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).