
Deconstructing the Dimensions of Mycobiome Fingerprints in Luohandu Cave, Guilin, Southern China

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cave Description and Sample Collection
2.2. DNA Extraction and Mycobiome Sequencing
2.3. Bioinformatics Analysis
3. Results
3.1. Physicochemical Properties of Weathered Rocks and Sediments in Luohandu Cave
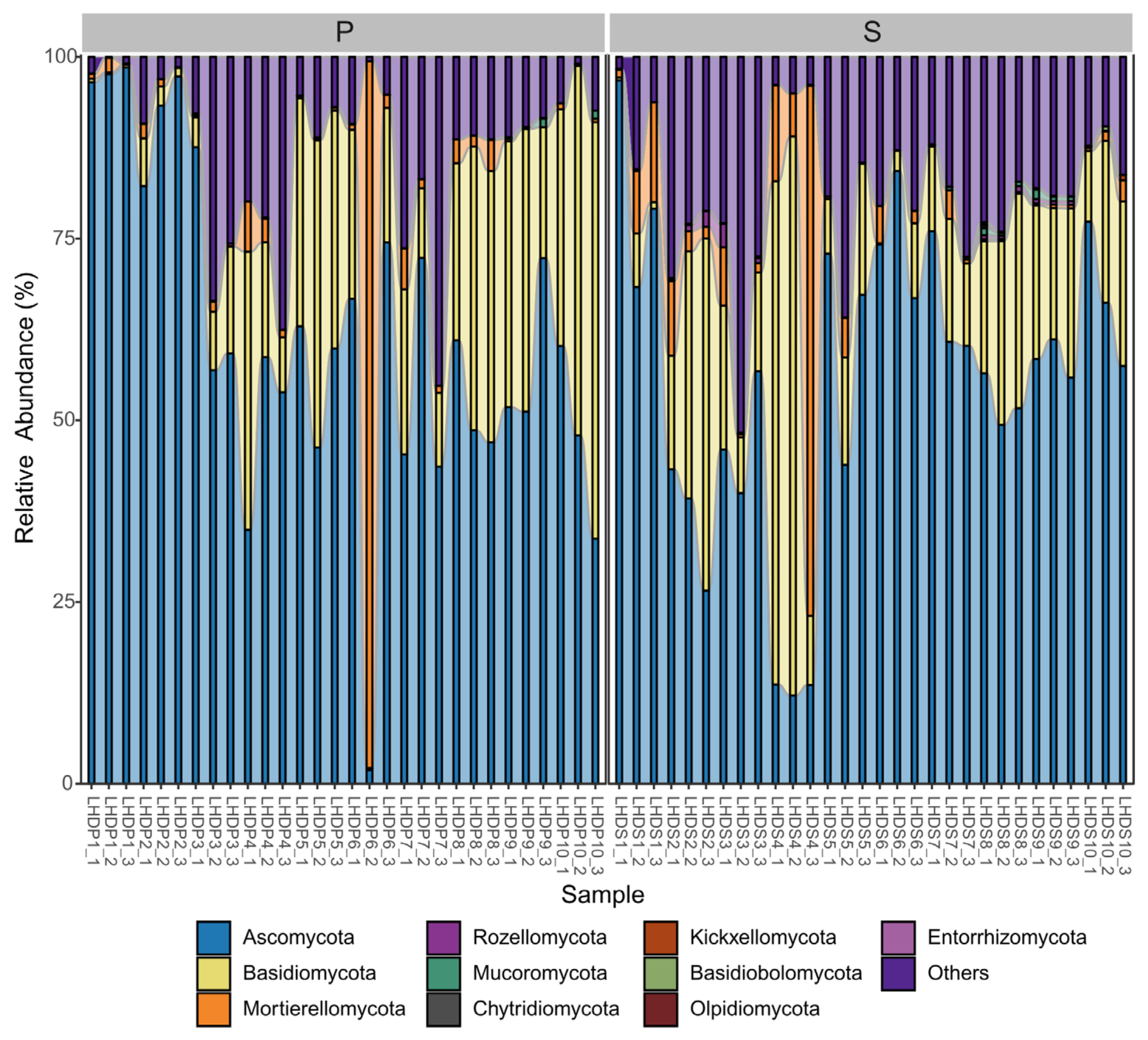
3.2. Mycobiome Community Structure and Interaction with the Environmental Conditions
3.3. Biodiversity Assessment of the Mycobiomes in Weathered Rocks and Sediments
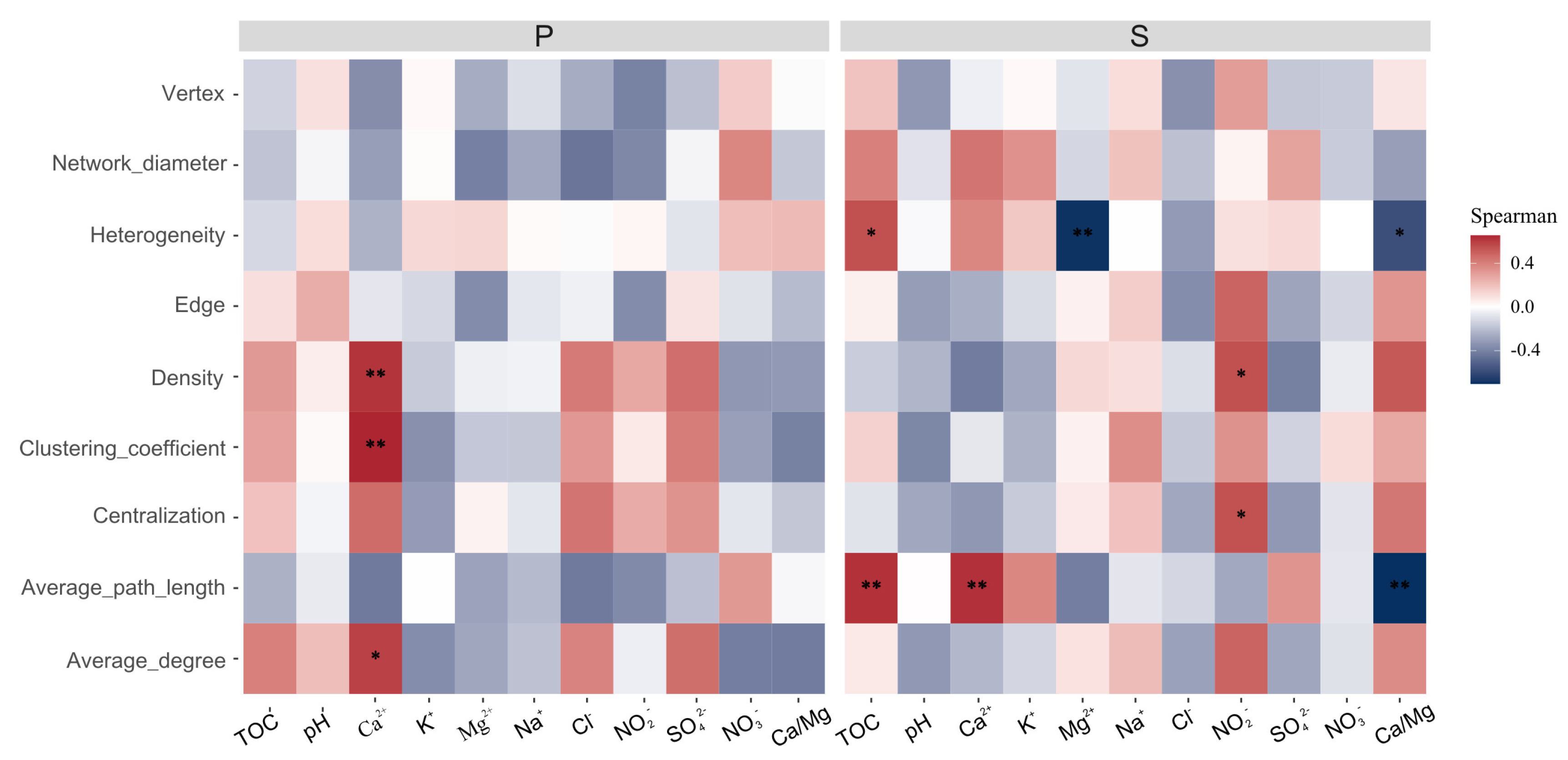
3.4. Mycobiome Interactions in Weathered Rocks and Sediments
4. Discussion
4.1. Disclosure of Luohandu Cave’s Potential “Hidden Diversity” in Mycobiomes
4.2. The Ca/Mg Ratio Significantly Affects the Mycobiome Community Structure
4.3. Ascomycota Dominate the Mycobiome Networks
4.4. Limitations and Future Investigations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Meiser, A.; Bálint, M.; Schmitt, I. Meta-analysis of deep-sequenced fungal communities indicates limited taxon sharing between studies and the presence of biogeographic patterns. New Phytol. 2013, 201, 623–635. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1–5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Tomczyk-Żak, K.; Zielenkiewicz, U. Microbial diversity in caves. Geomicrobiol. J. 2016, 33, 20–38. [Google Scholar] [CrossRef]
- Man, B.; Wang, H.; Yun, Y.; Xiang, X.; Wang, R.; Duan, Y.; Cheng, X. Diversity of fungal communities in Heshang Cave of Central China revealed by mycobiome-sequencing. Front. Microbiol. 2018, 9, 1400. [Google Scholar] [CrossRef] [PubMed]
- Bontemps, Z.; Prigent-Combaret, C.; Guillmot, A.; Hugoni, M.; Moënne-Loccoz, Y. Dark-zone alterations expand throughout Paleolithic Lascaux Cave despite spatial heterogeneity of the cave microbiome. Environ. Microbiome 2023, 18, 31. [Google Scholar] [CrossRef]
- Vanderwolf, K.; Malloch, D.; Mcalpine, D.; Forbes, G. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Man, B.; Wang, H.; Xiang, X.; Wang, R.; Yun, Y.; Gong, L. Phylogenetic diversity of culturable fungi in the Heshang Cave, central China. Front. Microbiol. 2015, 6, 1158. [Google Scholar] [CrossRef]
- Suetrong, S.; Preedanon, S.; Kobmoo, N.; Srihom, C.; Somrithipol, S.; Saengkaewsuk, S.; Srikitikulchai, P.; Klaysuban, A.; Nuankaew, S.; Chuaseeharonnachai, C. Unravelling the hidden diversity of cave mycobiota in Thailand’s Satun Geopark. Sci. Rep. 2023, 13, 19162. [Google Scholar] [CrossRef]
- Wasti, I.G.; Khan, F.A.A.; Bernard, H.; Hassan, N.H.; Fayle, T.; Sathiya Seelan, J.S. Fungal communities in bat guano, speleothem surfaces, and cavern water in Madai cave, Northern Borneo (Malaysia). Mycology 2021, 12, 188–202. [Google Scholar] [CrossRef]
- Alonso, L.; Creuzé-des-Châtelliers, C.; Trabac, T.; Dubost, A.; Moënne-Loccoz, Y.; Pommier, T. Rock substrate rather than black stain alterations drives microbial community structure in the passage of Lascaux Cave. Microbiome 2018, 6, 216. [Google Scholar] [CrossRef]
- Alves, V.; Lira, R.; Lima, J.; Barbosa, R.; Bento, D.; Barbier, E.; Bernard, E.; Souza-Motta, C.; Bezerra, J. Unravelling the fungal darkness in a tropical cave: Richness and the description of one new genus and six new species. Fungal Syst. Evol. 2022, 10, 139–167. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Speruda, M.; Borzęcka, J.; Piecuch, A.; Cal, M. First speleomycological study on the occurrence of psychrophilic and psychrotolerant aeromycota in the brestovská cave (Western Tatras Mts., Slovakia) and first reports for some species at underground sites. Biology 2021, 10, 497. [Google Scholar] [CrossRef] [PubMed]
- Karunarathna, S.C.; Dong, Y.; Karasaki, S.; Tibpromma, S.; Hyde, K.D.; Lumyong, S.; Xu, J.; Sheng, J.; Mortimer, P.E. Discovery of novel fungal species and pathogens on bat carcasses in a cave in Yunnan Province, China. Emerg. Microbes Infect. 2020, 9, 1554–1566. [Google Scholar] [CrossRef] [PubMed]
- Preedanon, S.; Suetrong, S.; Shrihom, S.; Somrithipol, S.; Kobmoo, N.; Saengkaewsuk, S.; Srikitikulchai, P.; Klaysuban, A.; Nuankaew, S.; Chuaseeharonnachai, C. Eight novel cave fungi in Thailand’s Satun Geopark. Fungal Syst. Evol. 2024, 12, 1–30. [Google Scholar] [CrossRef]
- Pereira, M.L.; Carvalho, J.L.; Lima, J.M.; Barbier, E.; Bernard, E.; Bezerra, J.D.; Souza-Motta, C.M. Richness of Cladosporium in a tropical bat cave with the description of two new species. Mycol. Prog. 2022, 21, 345–357. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, S.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable mycobiota from Karst caves in China II, with descriptions of 33 new species. Fungal Divers. 2021, 106, 29–136. [Google Scholar] [CrossRef]
- Vaughan, M.J.; Nelson, W.; Soderlund, C.; Maier, R.M.; Pryor, B.M. Assessing fungal community structure from mineral surfaces in Kartchner Caverns using multiplexed 454 pyrosequencing. Microb. Ecol. 2015, 70, 175–187. [Google Scholar] [CrossRef]
- Ruibal, C.; Gueidan, C.; Selbmann, L.; Gorbushina, A.; Crous, P.; Groenewald, J.; Muggia, L.; Grube, M.; Isola, D.; Schoch, C. Phylogeny of rock-inhabiting fungi related to Dothideomycetes. Stud. Mycol. 2009, 64, 123–133. [Google Scholar] [CrossRef]
- Egidi, E.; De Hoog, G.; Isola, D.; Onofri, S.; Quaedvlieg, W.; De Vries, M.; Verkley, G.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and taxonomy of meristematic rock-inhabiting black fungi in the Dothideomycetes based on multi-locus phylogenies. Fungal Divers. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Cai, L. Substrate and spatial variables are major determinants of fungal community in karst caves in Southwest China. J. Biogeogr. 2019, 46, 1504–1518. [Google Scholar] [CrossRef]
- Piano, E.; Biagioli, F.; Nicolosi, G.; Coleine, C.; Poli, A.; Prigione, V.; Zanellati, A.; Addesso, R.; Varese, G.C.; Selbmann, L. Tourism affects microbial assemblages in show caves. Sci. Total Environ. 2023, 871, 162106. [Google Scholar] [CrossRef] [PubMed]
- Biagioli, F.; Coleine, C.; Delgado-Baquerizo, M.; Feng, Y.; Saiz-Jimenez, C.; Selbmann, L. Outdoor climate drives diversity patterns of dominant microbial taxa in caves worldwide. Sci. Total Environ. 2024, 906, 167674. [Google Scholar] [CrossRef]
- Yang, Z.; Cheng, X.; Wang, H.; Zeng, Z.; LIiu, X.; Cao, J.; Su, C.; Huang, Q. Environmental driving mechanisms and community assembly process of bacterial communities in the Luohandu cave, Guilin, Guangxi Province, China. Acta Microbiol. Sin. 2021, 61, 4118–4136. [Google Scholar]
- Cheng, X.; Liu, X.; Wang, H.; Su, C.; Zhao, R.; Bodelier, P.L.; Wang, W.; Ma, L.; Lu, X. USC γ dominated community composition and cooccurrence network of methanotrophs and bacteria in subterranean karst caves. Microbiol. Spectr. 2021, 9, e00820–e00821. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, H.; Wang, W.; Cheng, X.; Wang, Y.; Li, Q.; Li, L.; Ma, L.; Lu, X.; Tuovinen, O.H. Nitrate determines the bacterial habitat specialization and impacts microbial functions in a subsurface karst cave. Front. Microbiol. 2023, 14, 1115449. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, R.; Kõljalg, U. UNITE QIIME Release for Eukaryotes, Version 04.02.; UNITE Community: Adelaide, SA, Australia, 2020. [Google Scholar]
- Xiang, X.; Wang, H.; Man, B.; Xu, Y.; Gong, L.; Tian, W.; Yang, H. Diverse Bathyarchaeotal lineages dominate archaeal communities in the acidic Dajiuhu peatland, Central China. Microb. Ecol. 2023, 85, 557–571. [Google Scholar] [CrossRef]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJournal Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Torres, T.; Ortiz, J.; Cobo, R. Features of deep cave sediments: Their influence on fossil preservation. Estud. Geológicos 2003, 59, 195–204. [Google Scholar]
- Epure, L.; Meleg, I.N.; Munteanu, C.-M.; Roban, R.D.; Moldovan, O.T. Bacterial and fungal diversity of quaternary cave sediment deposits. Geomicrobiol. J. 2014, 31, 116–127. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhou, Z.; Ding, S.; Zhang, H.; Huang, J.; Gong, X.; Su, D. Spatiotemporal variation characteristics and influencing factors of karst cave microclimate environments: A case study in Shuanghe Cave, Guizhou Province, China. Atmosphere 2023, 14, 813. [Google Scholar] [CrossRef]
- Wu, Y.; Tan, L.; Liu, W.; Wang, B.; Wang, J.; Cai, Y.; Lin, X. Profiling bacterial diversity in a limestone cave of the western Loess Plateau of China. Front. Microbiol. 2015, 6, 244. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S.; Di Piazza, S.; Ziotti, M.; Badino, G.; Marco, I. Human-induced alterations of the mycobiota in an alpine show cave (Italy, SW-Alps). Acta Carsologica 2017, 46, 111–123. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lücking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, F.; Zhou, X.; Liu, X.; Liu, S.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia-Mol. Phylogeny Evol. Fungi 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Wang, Y.; Cheng, X.; Wang, H.; Zhou, J.; Liu, X.; Tuovinen, O.H. The characterization of microbiome and interactions on weathered rocks in a subsurface karst cave, Central China. Front. Microbiol. 2022, 13, 909494. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Moñino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne fungi in show caves from Southern Spain. Appl. Sci. 2021, 11, 5027. [Google Scholar] [CrossRef]
- Dzurendova, S.; Zimmermann, B.; Kohler, A.; Reitzel, K.; Nielsen, U.G.; Dupuy--Galet, B.X.; Leivers, S.; Horn, S.J.; Shapaval, V. Calcium affects polyphosphate and lipid accumulation in Mucoromycota fungi. J. Fungi 2021, 7, 300. [Google Scholar] [CrossRef]
- Roy, A.; Kumar, A.; Baruah, D.; Tamuli, R. Calcium signaling is involved in diverse cellular processes in fungi. Mycology 2021, 12, 10–24. [Google Scholar] [CrossRef]
- Aoki, K.; Yamamoto, K.; Ohkuma, M.; Sugita, T.; Tanaka, N.; Takashima, M. Hyphal growth in trichosporon asahii is accelerated by the addition of magnesium. Microbiol. Spectr. 2023, 11, e04242-22. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Moral, S.; Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Ontañon, R.; Saiz-Jimenez, C. Environment-driven control of fungi in subterranean ecosystems: The case of La Garma Cave (northern Spain). Int. Microbiol. 2021, 24, 573–591. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.; Guo, J.; Li, Y.; Zhong, X.; Lv, Y.; Li, J.; Yang, A. The diversity of microbes and prediction of their functions in karst caves under the influence of human tourism activities—A case study of Zhijin Cave in Southwest China. Environ. Sci. Pollut. Res. 2022, 29, 25858–25868. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Cardona, C.; Li, Y.; Shi, Y.; Xiang, X.; Shen, C.; Wang, H.; Gilbert, J.A.; Chu, H. Rhizosphere-associated bacterial network structure and spatial distribution differ significantly from bulk soil in wheat crop fields. Soil Biol. Biochem. 2017, 113, 275–284. [Google Scholar] [CrossRef]
- Fillol, M.; Auguet, J.-C.; Casamayor, E.O.; Borrego, C.M. Insights in the ecology and evolutionary history of the Miscellaneous Crenarchaeotic Group lineage. ISME J. 2016, 10, 665–677. [Google Scholar] [CrossRef]
- Trosvik, P.; de Muinck, E.J. Ecology of bacteria in the human gastrointestinal tract—Identification of keystone and foundation taxa. Microbiome 2015, 3, 44. [Google Scholar] [CrossRef]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Man, B.-Y.; Xiang, X.; Cheng, X.-Y.; Wang, H.-M.; Su, C.-T.; Huang, Q.-B.; Luo, Y.; Zhang, C.; Cheng, G.; Ni, Y.-Y.; et al. Deconstructing the Dimensions of Mycobiome Fingerprints in Luohandu Cave, Guilin, Southern China. Microorganisms 2024, 12, 211. https://doi.org/10.3390/microorganisms12010211
Man B-Y, Xiang X, Cheng X-Y, Wang H-M, Su C-T, Huang Q-B, Luo Y, Zhang C, Cheng G, Ni Y-Y, et al. Deconstructing the Dimensions of Mycobiome Fingerprints in Luohandu Cave, Guilin, Southern China. Microorganisms. 2024; 12(1):211. https://doi.org/10.3390/microorganisms12010211
Chicago/Turabian StyleMan, Bai-Ying, Xing Xiang, Xiao-Yu Cheng, Hong-Mei Wang, Chun-Tian Su, Qi-Bo Huang, Yang Luo, Chao Zhang, Gang Cheng, Yu-Yang Ni, and et al. 2024. "Deconstructing the Dimensions of Mycobiome Fingerprints in Luohandu Cave, Guilin, Southern China" Microorganisms 12, no. 1: 211. https://doi.org/10.3390/microorganisms12010211
APA StyleMan, B.-Y., Xiang, X., Cheng, X.-Y., Wang, H.-M., Su, C.-T., Huang, Q.-B., Luo, Y., Zhang, C., Cheng, G., Ni, Y.-Y., & Shao, X.-H. (2024). Deconstructing the Dimensions of Mycobiome Fingerprints in Luohandu Cave, Guilin, Southern China. Microorganisms, 12(1), 211. https://doi.org/10.3390/microorganisms12010211