
Harmful and Harmless Soil-Dwelling Fungi Indicate Microhabitat Suitability for Off-Host Ixodid Ticks


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Tick Maintenance
2.2. Propagation of Fungi and Collection of Conidia
2.3. Preparation of Experimental Substrates
2.4. General Experimental Design
2.5. Specific Experiments
2.5.1. Effect of Bb-Inoculated Substrate on Behavioral Responses of Diverse Tick Taxa
2.5.2. Effect of Bb-Incubation Period on Behavioral Responses of Ticks
2.5.3. Olfactory or Contact-Chemoreceptive Recognition of Bb (or Its Metabolites) by Ticks
2.5.4. Effect of Cellulose or Its Fungal Metabolites on Behavioral Responses of Ticks
2.5.5. Effect of Various Soil-Dwelling Fungi on Behavioral Responses of Ticks
2.5.6. Effect of Fungus-Derived Volatiles on Attraction of Ticks
2.6. Statistical Analysis
3. Results
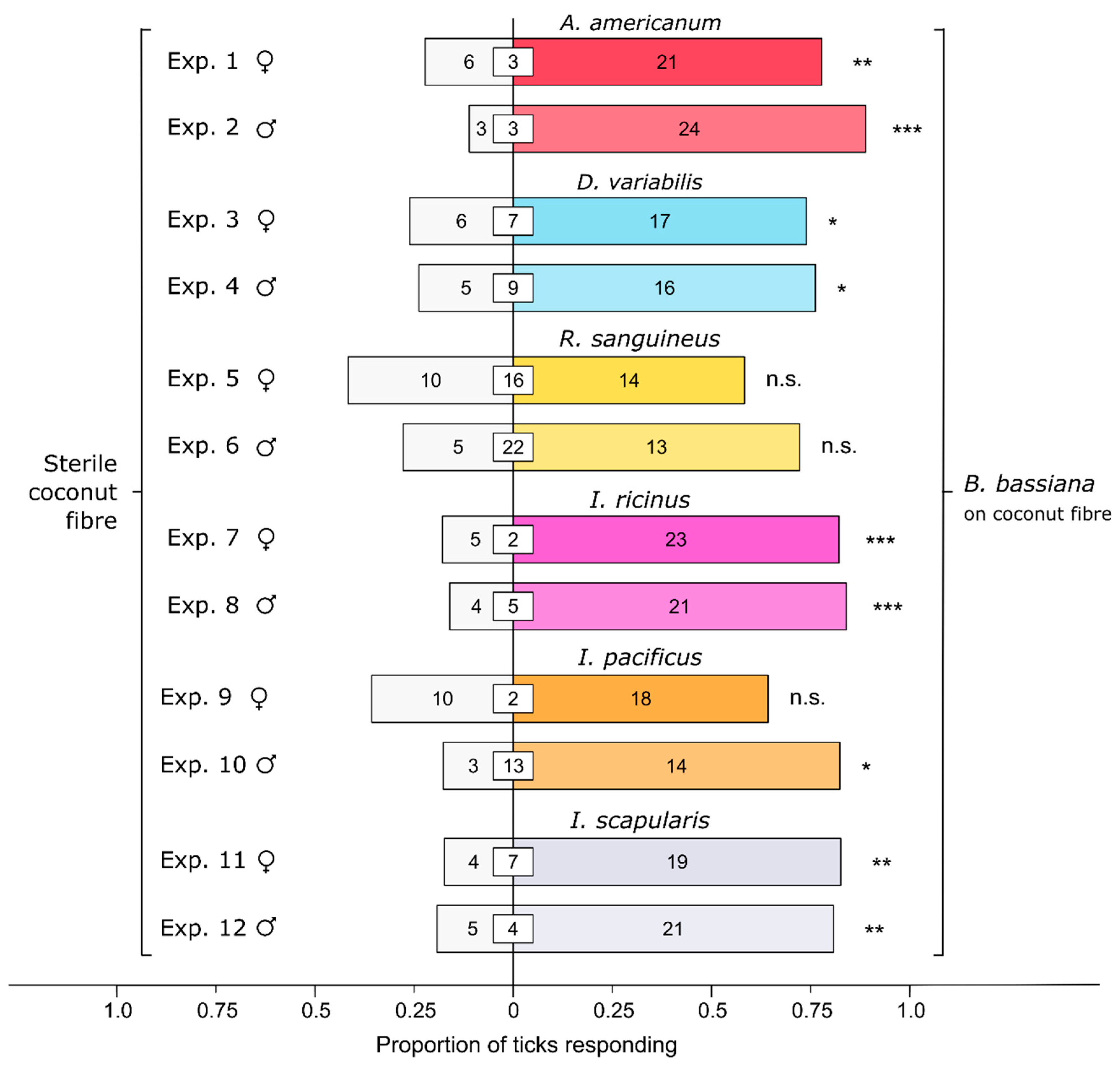
3.1. Effect of Bb-Inoculated Substrate on Behavioral Responses of Ticks
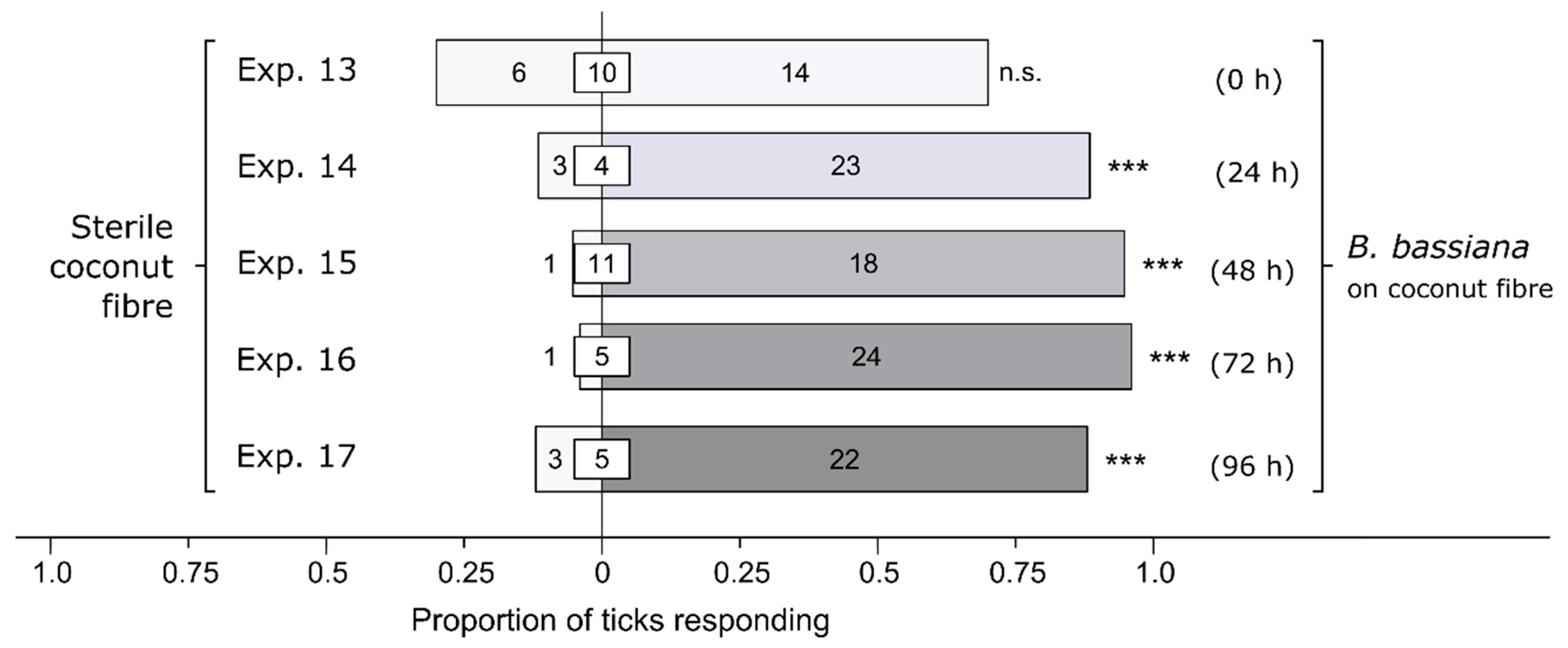
3.2. Effect of Bb-Incubation Period on Behavioral Responses of Ticks
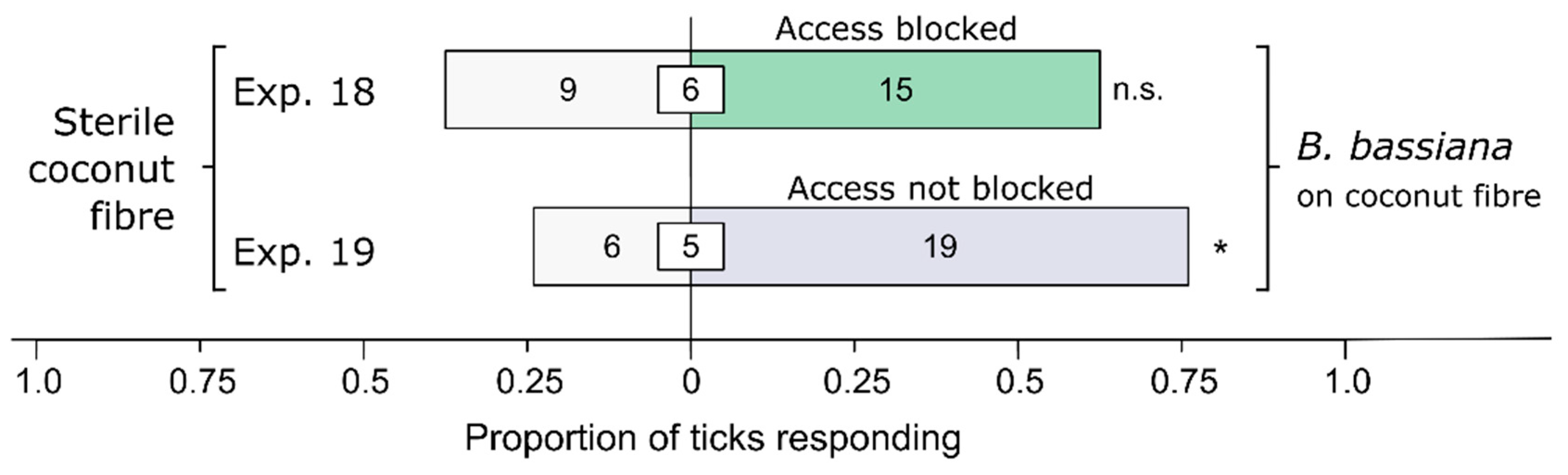
3.3. Olfactory or Contact-Chemoreceptive Recognition of Bb (or Its Metabolites) by Ticks
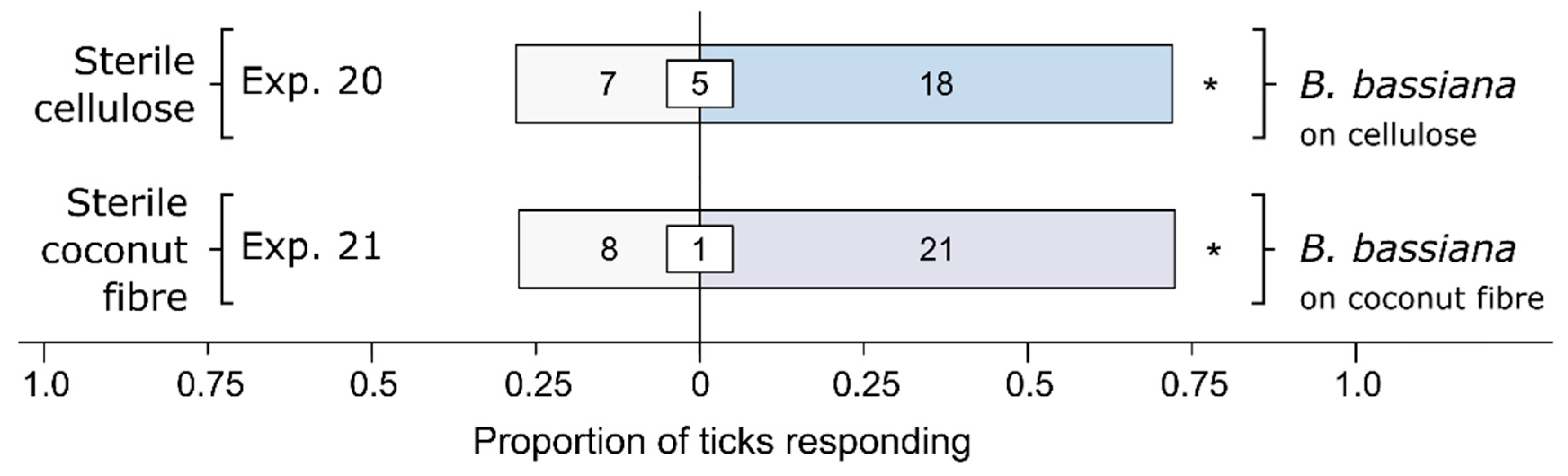
3.4. Effect of Cellulose or Its Fungal Metabolites on Behavioral Responses of Ticks
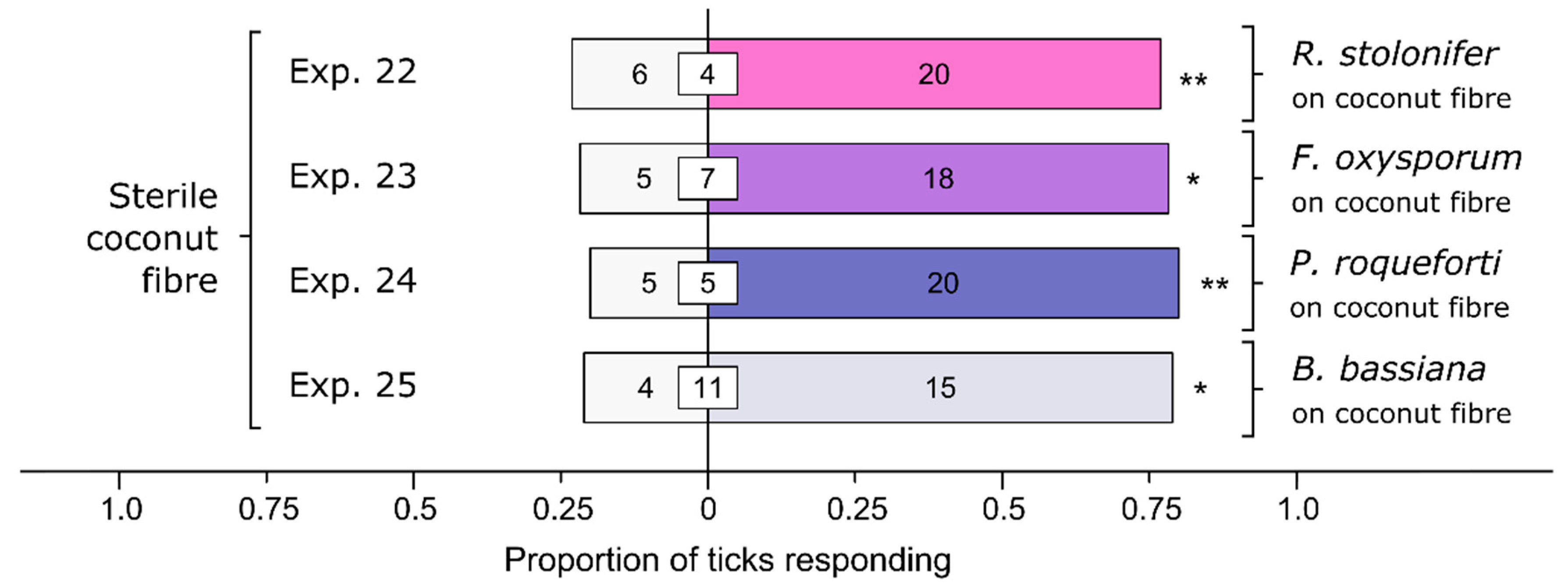
3.5. Effect of Various Soil-Dwelling Fungi on Behavioral Responses of Ticks
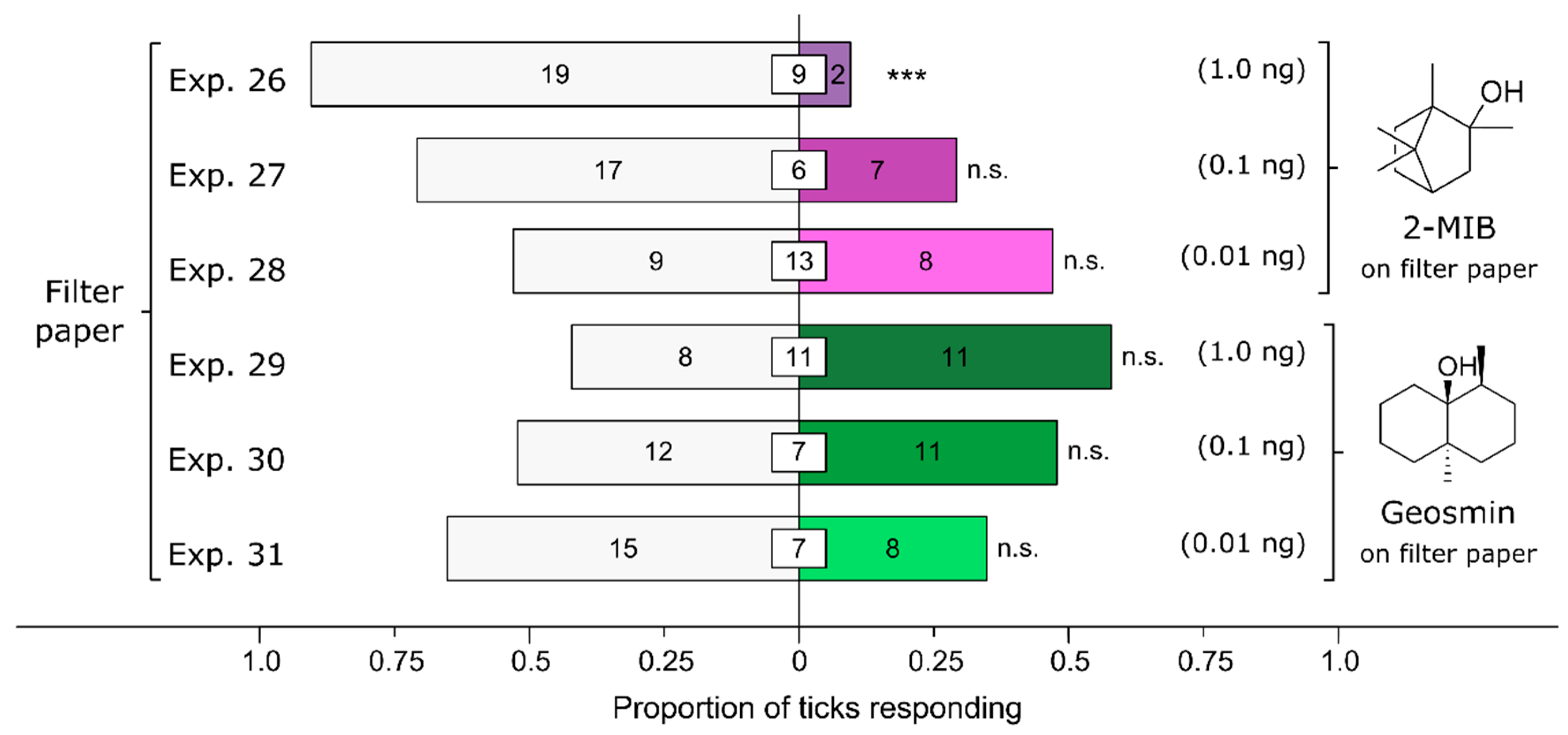
3.6. Effect of Fungus-Derived Volatiles on Attraction of Ticks
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wikel, S.K. Tick-Host Interactions. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2, pp. 88–128. [Google Scholar]
- Gray, J.S.; Estrada-Peña, A.; Vial, L. Ecology of Nidicolous Ticks. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2, pp. 3–38. [Google Scholar]
- Zając, Z.; Bartosik, K.; Woźniak, A. Monitoring Dermacentor reticulatus host-seeking activity in natural conditions. Insects 2020, 11, 264. [Google Scholar] [CrossRef]
- Suzin, A.; Vogliotti, A.; Nunes, P.H.; Barbieri, A.R.M.; Labruna, M.B.; Szabó, M.P.J. Free-living ticks (Acari: Ixodidae) in the Iguaçu National Park, Brazil: Temporal dynamics and questing behavior on vegetation. Ticks Tick. Borne. Dis. 2020, 11, 101471. [Google Scholar] [CrossRef]
- Loye, J.E.; Lane, R.S. Questing behavior of Ixodes pacificus (Acari: Ixodidae) in relation to meteorological and seasonal factors. J. Med. Entomol. 1988, 25, 391–398. [Google Scholar] [CrossRef]
- Arsnoe, I.M.; Hickling, G.J.; Ginsberg, H.S.; McElreath, R.; Tsao, J.I. Different populations of blacklegged tick nymphs exhibit differences in questing behavior that have implications for human Lyme disease risk. PloS ONE 2015, 10, e0127450. [Google Scholar] [CrossRef]
- Mejlon, H.A.; Jaenson, T.G.T. Questing behaviour of Ixodes ricinus ticks (Acari: Ixodidae). Exp. Appl. Acarol. 1997, 21, 747–754. [Google Scholar] [CrossRef]
- Waladde, S.M.; Rice, M.J. The sensory basis of tick feeding behaviour. In Physiology of Ticks; Obenchain, F.D., Galun, R., Eds.; Pergamon Press: Oxford, UK, 1982; pp. 71–118. [Google Scholar]
- Crooks, E.; Randolph, S.E. Walking by Ixodes ricinus ticks: Intrinsic and extrinsic factors determine the attraction of moisture or host odour. J. Exp. Biol. 2006, 209, 2138–2142. [Google Scholar] [CrossRef]
- Sonenshine, D.E. Tick pheromones and their use in tick control. Annu. Rev. Entomol. 2006, 51, 557–580. [Google Scholar] [CrossRef]
- Randolph, S.E. Ecology of Non-Nidicolous Ticks. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2. [Google Scholar]
- Vail, S.G.; Smith, G. Vertical movement and posture of blacklegged tick (Acari: Ixodidae) nymphs as a function of temperature and relative humidity in laboratory experiments. J. Med. Entomol. 2002, 39, 842–846. [Google Scholar] [CrossRef]
- Perret, J.L.; Guerin, P.M.; Diehl, P.A.; Vlimant, M.; Gern, L. Darkness induces mobility, and saturation deficit limits questing duration, in the tick Ixodes ricinus. J. Exp. Biol. 2003, 206, 1809–1815. [Google Scholar] [CrossRef]
- Lubelczyk, C.B.; Elias, S.P.; Rand, P.W.; Holman, M.S.; Lacombe, E.H.; Smith, R.P. Habitat associations of Ixodes scapularis (Acari: Ixodidae) in Maine. Community Ecosyst. Ecol. 2004, 33, 900–906. [Google Scholar] [CrossRef]
- Bertrand, M.R.; Wilson, M.L. Microclimate-dependent survival of unfed adult Ixodes scapularis (Acari: Ixodidae) in nature: Life cycle and study design implications. J. Med. Entomol. 1996, 33, 619–627. [Google Scholar] [CrossRef]
- Goddard, J. Ticks and tick ecology in Mississippi: Implications for human disease transmission. J. Mississippi Acad. Sci. 2001, 46, 100–101. [Google Scholar]
- Burtis, J.C.; Yavitt, J.B.; Fahey, T.J.; Ostfeld, R.S. Ticks as soil-dwelling arthropods: An intersection between disease and soil ecology. J. Med. Entomol. 2019, 56, 1555–1564. [Google Scholar] [CrossRef]
- Linske, M.A.; Stafford, K.C.; Williams, S.C.; Lubelczyk, C.B.; Welch, M.; Henderson, E.F. Impacts of deciduous leaf litter and snow presence on nymphal Ixodes scapularis (Acari: Ixodidae) overwintering survival in coastal New England, USA. Insects 2019, 10, 227. [Google Scholar] [CrossRef]
- Leal, B.; Zamora, E.; Fuentes, A.; Thomas, D.B.; Dearth, R.K. Questing by tick larvae (Acari: Ixodidae): A review of the influences that affect off-host survival. Ann. Entomol. Soc. Am. 2020, 113, 425–438. [Google Scholar] [CrossRef]
- Cusumano, A.; Lievens, B. Microbe-mediated alterations in floral nectar: Consequences for insect parasitoids. Curr. Opin. Insect Sci. 2023, 60, 101116. [Google Scholar] [CrossRef]
- Peach, D.A.H.; Carroll, C.; Meraj, S.; Gomes, S.; Galloway, E.; Balcita, A.; Coatsworth, H.; Young, N.; Uriel, Y.; Gries, R.; et al. Nectar-dwelling microbes of common tansy are attractive to its mosquito pollinator, Culex pipiens L. BMC Ecol. Evol. 2021, 21, 29. [Google Scholar] [CrossRef]
- Hubbard, C.J.; Li, B.; McMinn, R.; Brock, M.T.; Maignien, L.; Ewers, B.E.; Kliebenstein, D.; Weinig, C. The effect of rhizosphere microbes outweighs host plant genetics in reducing insect herbivory. Mol. Ecol. 2019, 28, 1801–1811. [Google Scholar] [CrossRef]
- Tao, L.; Hunter, M.D.; de Roode, J.C. Microbial root mutualists affect the predators and pathogens of herbivores above ground: Mechanisms, magnitudes, and missing links. Front. Ecol. Evol. 2017, 5, 160. [Google Scholar] [CrossRef]
- Romero, A.; Broce, A.; Zurek, L. Role of bacteria in the oviposition behaviour and larval development of stable flies. Med. Vet. Entomol. 2006, 20, 115–121. [Google Scholar] [CrossRef]
- Melo, N.; Wolff, G.H.; Costa-da-Silva, A.L.; Arribas, R.; Triana, M.F.; Gugger, M.; Riffell, J.A.; DeGennaro, M.; Stensmyr, M.C. Geosmin attracts Aedes aegypti mosquitoes to oviposition sites. Curr. Biol. 2020, 30, 127–134.e5. [Google Scholar] [CrossRef]
- Girard, M.; Martin, E.; Vallon, L.; Raquin, V.; Bellet, C.; Rozier, Y.; Desouhant, E.; Hay, A.E.; Luis, P.; Moro, C.V.; et al. Microorganisms associated with mosquito oviposition sites: Implications for habitat selection and insect life histories. Microorganisms 2021, 9, 1589. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Crippen, T.L.; Holmes, L.; Singh, B.; Pimsler, M.L.; Benbow, M.E.; Tarone, A.M.; Dowd, S.; Yu, Z.; Vanlaerhoven, S.L.; et al. Bacteria mediate oviposition by the black soldier fly, Hermetia illucens (L.), (Diptera: Stratiomyidae). Sci. Rep. 2013, 3, srep02563. [Google Scholar] [CrossRef]
- Long, J.; Maskell, K.; Gries, R.; Nayani, S.; Gooding, C.; Gries, G. Synergistic attraction of western black-legged ticks, Ixodes pacificus, to CO2 and odorant emissions from deer-associated microbes. R. Soc. Open Sci. 2023, 10, 230084. [Google Scholar] [CrossRef]
- Becher, P.G.; Verschut, V.; Bibb, M.J.; Bush, M.J.; Molnár, B.P.; Barane, E.; Al-Bassam, M.M.; Chandra, G.; Song, L.; Challis, G.L.; et al. Developmentally regulated volatiles geosmin and 2-methylisoborneol attract a soil arthropod to Streptomyces bacteria promoting spore dispersal. Nat. Microbiol. 2020, 5, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ren, L.; Li, H.; Schmidt, A.; Gershenzon, J.; Lu, Y.; Cheng, D. The nesting preference of an invasive ant is associated with the cues produced by actinobacteria in soil. PloS Pathog. 2020, 16, e1008800. [Google Scholar] [CrossRef] [PubMed]
- Tabachek, J.-A.L.; Yurkowski, M. Isolation and identification of blue-green algae producing muddy odor metabolites, geosmin, and 2-methylisoborneol, in saline lakes in Manitoba. J. Fish. Res. Board Canada 1976, 33, 25–35. [Google Scholar] [CrossRef]
- Safferman, R.S.; Rosen, A.A.; Mashni, C.I.; Morris, M.E. Earthy-smelling substance from a blue-green alga. Environ. Sci. Technol. 1967, 1, 429–430. [Google Scholar] [CrossRef]
- Klausen, C.; Nicolaisen, M.H.; Strobel, B.W.; Warnecke, F.; Nielsen, J.L.; Jørgensen, N.O.G. Abundance of actinobacteria and production of geosmin and 2-methylisoborneol in Danish streams and fish ponds. FEMS Microbiol. Ecol. 2005, 52, 265–278. [Google Scholar] [CrossRef]
- La Guerche, S.; Dauphin, B.; Pons, M.; Blancard, D.; Darriet, P. Characterization of some mushroom and earthy off-odors microbially induced by the development of rot on grapes. J. Agric. Food Chem. 2006, 54, 9193–9200. [Google Scholar] [CrossRef]
- Fravel, D.R.; Connick, W.J.; Grimm, C.C.; Lloyd, S.W. Volatile compounds emitted by sclerotia of Sclerotinia minor, Sclerotinia sclerotiorum, and Sclerotium rolfsii. J. Agric. Food Chem. 2002, 50, 3761–3764. [Google Scholar] [CrossRef]
- Mattheis, J.P.; Roberts, R.G. Identification of geosmin as a volatile metabolite of Penicillium expansum. Appl. Environ. Microbiol. 1992, 58, 3170–3172. [Google Scholar] [CrossRef]
- Gooding, C.E.; Pinard, C.; Gries, R.; Devireddy, A.; Gries, G. Blacklegged ticks, Ixodes scapularis, reduce predation risk by eavesdropping on communication signals of Formica oreas thatching ants. R. Soc. Open Sci. 2024, 11, 231355. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Tan, K.; Qu, Y.; Nieh, J.C. Giant Asian honeybees use olfactory eavesdropping to detect and avoid ant predators. Anim. Behav. 2014, 97, 69–76. [Google Scholar] [CrossRef]
- Storm, J.J.; Lima, S.L. Predator-naïve fall field crickets respond to the chemical cues of wolf spiders. Can. J. Zool. 2008, 86, 1259–1263. [Google Scholar] [CrossRef]
- Sidhu, C.S.; Wilson Rankin, E.E. Honey bees avoiding ant harassment at flowers using scent cues. Environ. Entomol. 2016, 45, 420–426. [Google Scholar] [CrossRef]
- Kempraj, V.; Park, S.J.; Taylor, P.W. Forewarned is forearmed: Queensland fruit flies detect olfactory cues from predators and respond with predator-specific behaviour. Sci. Rep. 2020, 10, 7297. [Google Scholar] [CrossRef]
- Mestre, L.; Bucher, R.; Entling, M.H. Trait-mediated effects between predators: Ant chemical cues induce spider dispersal. J. Zool. 2014, 293, 119–125. [Google Scholar] [CrossRef]
- Bucher, R.; Binz, H.; Menzel, F.; Entling, M.H. Effects of spider chemotactile cues on arthropod behavior. J. Insect Behav. 2014, 27, 567–580. [Google Scholar] [CrossRef]
- Bucher, R.; Menzel, F.; Entling, M.H. Risk of spider predation alters food web structure and reduces local herbivory in the field. Oecologia 2015, 178, 571–577. [Google Scholar] [CrossRef]
- Ormond, E.L.; Thomas, A.P.M.; Pugh, P.J.A.; Pell, J.K.; Roy, H.E. A fungal pathogen in time and space: The population dynamics of Beauveria bassiana in a conifer forest. FEMS Microbiol. Ecol. 2010, 74, 146–154. [Google Scholar] [CrossRef]
- Shimazu, M.; Sato, H.; Maehara, N. Density of the entomopathogenic fungus, Beauveria bassiana Vuillemin (Deuteromycotina: Hyphomycetes) in forest air and soil. J. Appl. Entomol. Zool. 2002, 37, 19–26. [Google Scholar] [CrossRef]
- Meyling, N.V.; Eilenberg, J. Ecology of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae in temperate agroecosystems: Potential for conservation biological control. Biol. Control 2007, 43, 145–155. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef]
- Kirkland, B.H.; Westwood, G.S.; Keyhani, N.O. Pathogenicity of entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae to Ixodidae tick species Dermacentor variabilis, Rhipicephalus sanguineus, and Ixodes scapularis. J. Med. Entomol. 2004, 41, 705–711. [Google Scholar] [CrossRef]
- Sullivan, C.F.; Parker, B.L.; Skinner, M. A review of commercial Metarhizium and Beauveria based biopesticides for the biological control of ticks in the USA. Insects 2022, 13, 260. [Google Scholar] [CrossRef] [PubMed]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2004, 31, 162–171. [Google Scholar] [CrossRef]
- Ormond, E.L.; Thomas, A.P.M.; Pell, J.K.; Freeman, S.N.; Roy, H.E. Avoidance of a generalist entomopathogenic fungus by the ladybird, Coccinella septempunctata. FEMS Microbiol. Ecol. 2011, 77, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Mburu, D.M.; Ochola, L.; Maniania, N.K.; Njagi, P.G.N.; Gitonga, L.M.; Ndung’u, M.W.; Wanjoya, A.K.; Hassanali, A. Relationship between virulence and repellency of entomopathogenic isolates of Metarhizium anisopliae and Beauveria bassiana to the termite Macrotermes michaelseni. J. Insect Physiol. 2009, 55, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Alma, C.R.; Gillespie, D.R.; Roitberg, B.D.; Goettel, M.S. Threat of infection and threat-avoidance behavior in the predator Dicyphus esperus feeding on whitefly nymphs infected with an entomopathogen. J. Insect Behav. 2010, 23, 90–99. [Google Scholar] [CrossRef]
- Wu, S.; Xing, Z.; Sun, W.; Xu, X.; Meng, R.; Lei, Z. Effects of Beauveria bassiana on predation and behavior of the predatory mite Phytoseiulus persimilis. J. Invertebr. Pathol. 2018, 153, 51–56. [Google Scholar] [CrossRef]
- Thompson, S.R.; Brandenburg, R.L.; Roberson, G.T. Entomopathogenic fungi detection and avoidance by mole crickets (Orthoptera: Gryllotalpidae). Environ. Entomol. 2007, 36, 165–172. [Google Scholar] [CrossRef] [PubMed]
- George, J.; Jenkins, N.E.; Blanford, S.; Thomas, M.B.; Baker, T.C. Malaria mosquitoes attracted by fatal fungus. PloS ONE 2013, 8, e62632. [Google Scholar] [CrossRef] [PubMed]
- Geedi, R.; Canas, L.; Reding, M.E.; Ranger, C.M. Attraction of Myzus persicae (Hemiptera: Aphididae) to volatiles emitted from the entomopathogenic fungus Beauveria bassiana. Environ. Entomol. 2023, 52, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Bissinger, B.W.; Roe, R.M. Tick Repellent Research, Methods, and Development. In Biology of Ticks; Sonenshine, D.E., Roe, R.M., Eds.; Oxford University Press: New York, NY, USA, 2014; Volume 2, pp. 382–408. [Google Scholar]
- Ritz, K.; Young, I.M. Interactions between soil structure and fungi. Mycologist 2004, 18, 52–59. [Google Scholar] [CrossRef]
- Albariño, R.; Villanueva, V.D.; Canhoto, C. The effect of sunlight on leaf litter quality reduces growth of the shredder Klapopteryx kuscheli. Freshw. Biol. 2008, 53, 1881–1889. [Google Scholar] [CrossRef]
- Pieristè, M.; Forey, E.; Lounès-Hadj Sahraoui, A.; Meglouli, H.; Laruelle, F.; Delporte, P.; Robson, T.M.; Chauvat, M. Spectral composition of sunlight affects the microbial functional structure of beech leaf litter during the initial phase of decomposition. Plant Soil 2020, 451, 515–530. [Google Scholar] [CrossRef]
- Baldrian, P.; Merhautová, V.; Petránková, M.; Cajthaml, T.; Šnajdr, J. Distribution of microbial biomass and activity of extracellular enzymes in a hardwood forest soil reflect soil moisture content. Appl. Soil Ecol. 2010, 46, 177–182. [Google Scholar] [CrossRef]
- Samanta, A.K.; Basu, G.; Mishra, L. Role of major constituents of coconut fibres on absorption of ionic dyes. Ind. Crops Prod. 2018, 117, 20–27. [Google Scholar] [CrossRef]
- Eggins, H.O.W.; Pugh, G.J.F. Isolation of cellulose-decomposing fungi from the soil. Nature 1962, 193, 94–95. [Google Scholar] [CrossRef]
- Conrady, M.W.; Bauer, M.; Jo, K.D.; Cropek, D.M.; Busby, R.R. Solid-phase microextraction (SPME) for determination of geosmin and 2-methylisoborneol in volatile emissions from soil disturbance. Chemosphere 2021, 284, 131333. [Google Scholar] [CrossRef]
- Wickham, H.; Seidel, D. Scales: Scale Functions for Visualization 2022. Available online: https://cran.r-project.org/package=scales (accessed on 11 December 2023).
- Dantas-Torres, F. Biology and ecology of the brown dog tick, Rhipicephalus sanguineus. Parasit. Vectors 2010, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Knülle, W.; Rudolph, D. Humidity relationships and water balance of ticks. In Physiology of Ticks; Obenchain, F.D., Galun, R., Eds.; Pergamon Press: Oxford, UK, 1982; pp. 43–70. [Google Scholar]
- Baldrian, P.; Valášková, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [PubMed]
- Carlile, M.J.; Watkinson, S.C.; Gooday, G.W. Fungal Cells and Vegetative Growth. In The Fungi; Carlile, M.J., Watkinson, S.C., Gooday, G.W., Eds.; Academic Press: London, UK, 2001; pp. 85–184. [Google Scholar]
- Trabalon, M. Chemical Communication and Contact Cuticular Compounds in Spiders. In Spider Ecophysiology; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 125–140. [Google Scholar]
- Stensmyr, M.C.; Dweck, H.K.M.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [PubMed]
- Zaroubi, L.; Ozugergin, I.; Mastronardi, K.; Imfeld, A.; Law, C.; Gélinas, Y.; Piekny, A.; Findlay, B.L. The ubiquitous soil terpene geosmin acts as a warning chemical. Appl. Environ. Microbiol. 2022, 88, e0009322. [Google Scholar] [CrossRef] [PubMed]
- Busby, R.R.; Conrady, M.W.; Jo, K.D.; Cropek, D.M. Characterising earth scent. Environ. Chem. 2023, 20, 226–234. [Google Scholar] [CrossRef]
- Carroll, J.F.; Benante, J.P.; Kramer, M.; Lohmeyer, K.H.; Lawrence, K. Formulations of DEET, picaridin, and IR3535 applied to skin repel nymphs of the lone star tick (Acari: Ixodidae) for 12 hours. J. Med. Entomol. 2010, 47, 699–704. [Google Scholar] [CrossRef]
- Benelli, G.; Pavela, R. Repellence of essential oils and selected compounds against ticks—A systematic review. Acta Trop. 2018, 179, 47–54. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gooding, C.E.; Gould, L.; Gries, G. Harmful and Harmless Soil-Dwelling Fungi Indicate Microhabitat Suitability for Off-Host Ixodid Ticks. Microorganisms 2024, 12, 609. https://doi.org/10.3390/microorganisms12030609
Gooding CE, Gould L, Gries G. Harmful and Harmless Soil-Dwelling Fungi Indicate Microhabitat Suitability for Off-Host Ixodid Ticks. Microorganisms. 2024; 12(3):609. https://doi.org/10.3390/microorganisms12030609
Chicago/Turabian StyleGooding, Claire E., Layla Gould, and Gerhard Gries. 2024. "Harmful and Harmless Soil-Dwelling Fungi Indicate Microhabitat Suitability for Off-Host Ixodid Ticks" Microorganisms 12, no. 3: 609. https://doi.org/10.3390/microorganisms12030609
APA StyleGooding, C. E., Gould, L., & Gries, G. (2024). Harmful and Harmless Soil-Dwelling Fungi Indicate Microhabitat Suitability for Off-Host Ixodid Ticks. Microorganisms, 12(3), 609. https://doi.org/10.3390/microorganisms12030609