
Expanding the CRISPR Toolbox for Engineering Lycopene Biosynthesis in Corynebacterium glutamicum

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Plasmid Construction
2.3. Plasmid Curing for Iterative Gene Editing
2.4. Determination of Cell Growth and Glucose Concentration
2.5. Extraction and Quantification of Lycopene from the C. glutamicum
3. Results
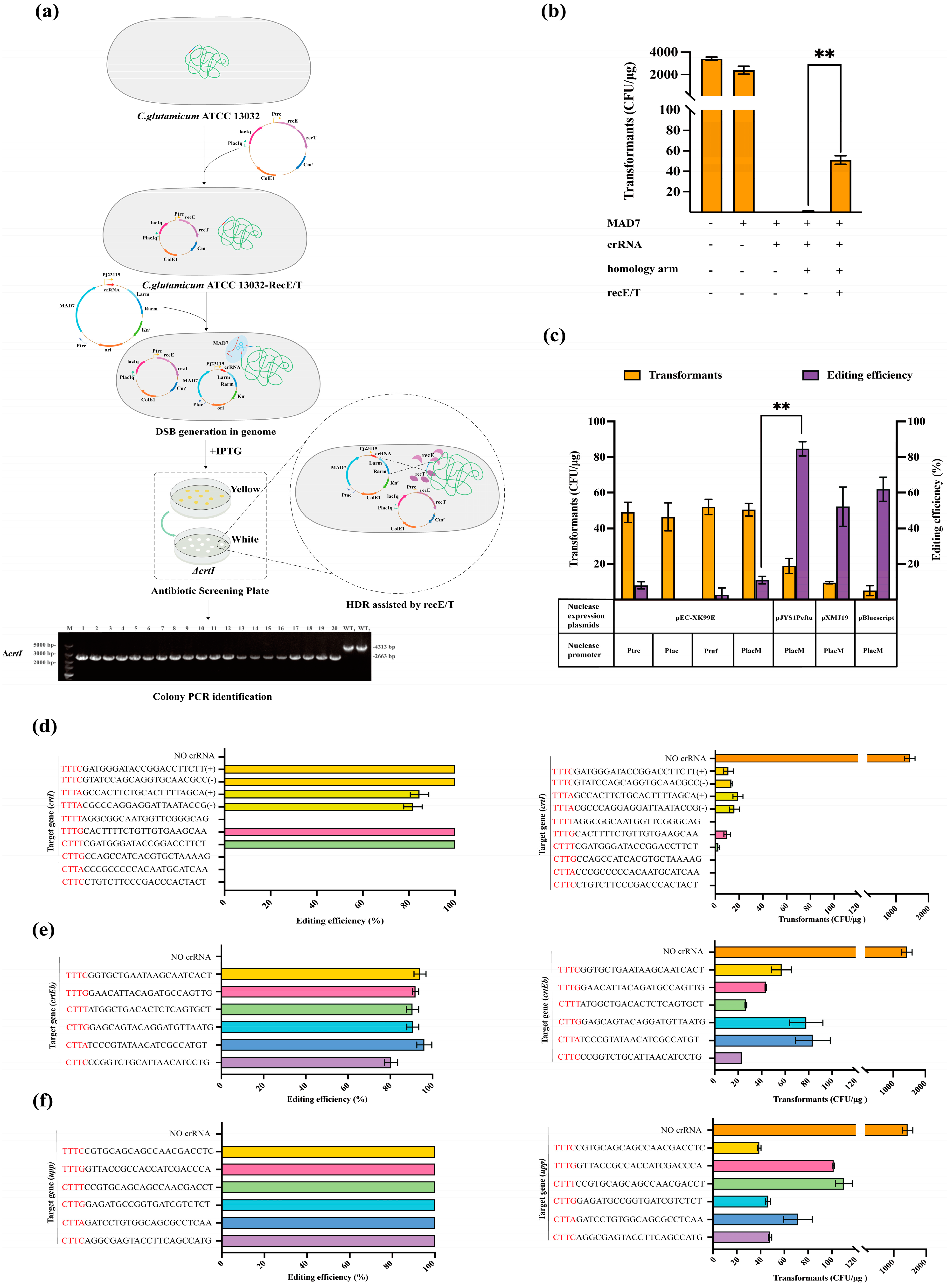
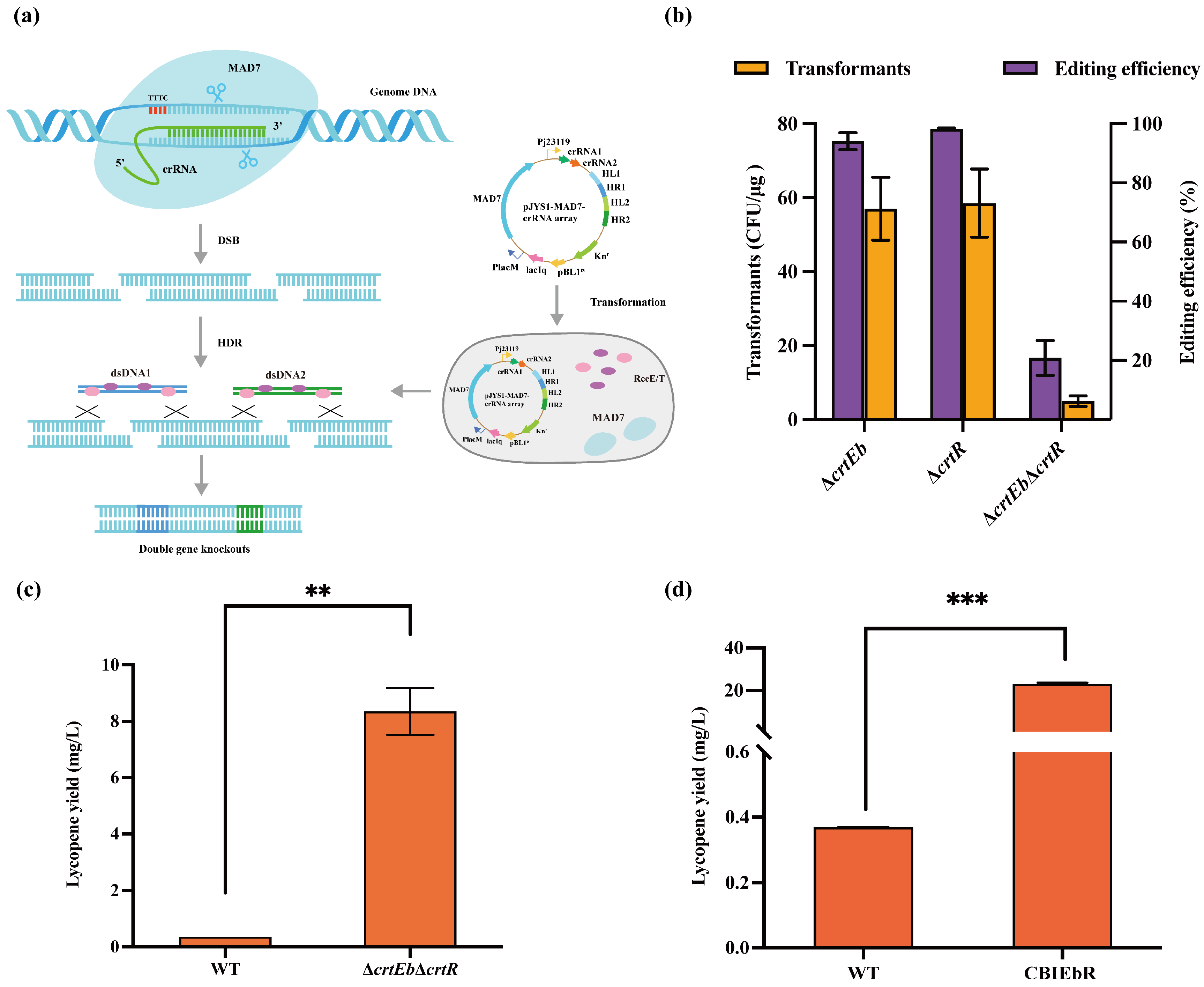
3.1. Establishment and Optimization of CRISPR System Based on MAD7 Nucleases
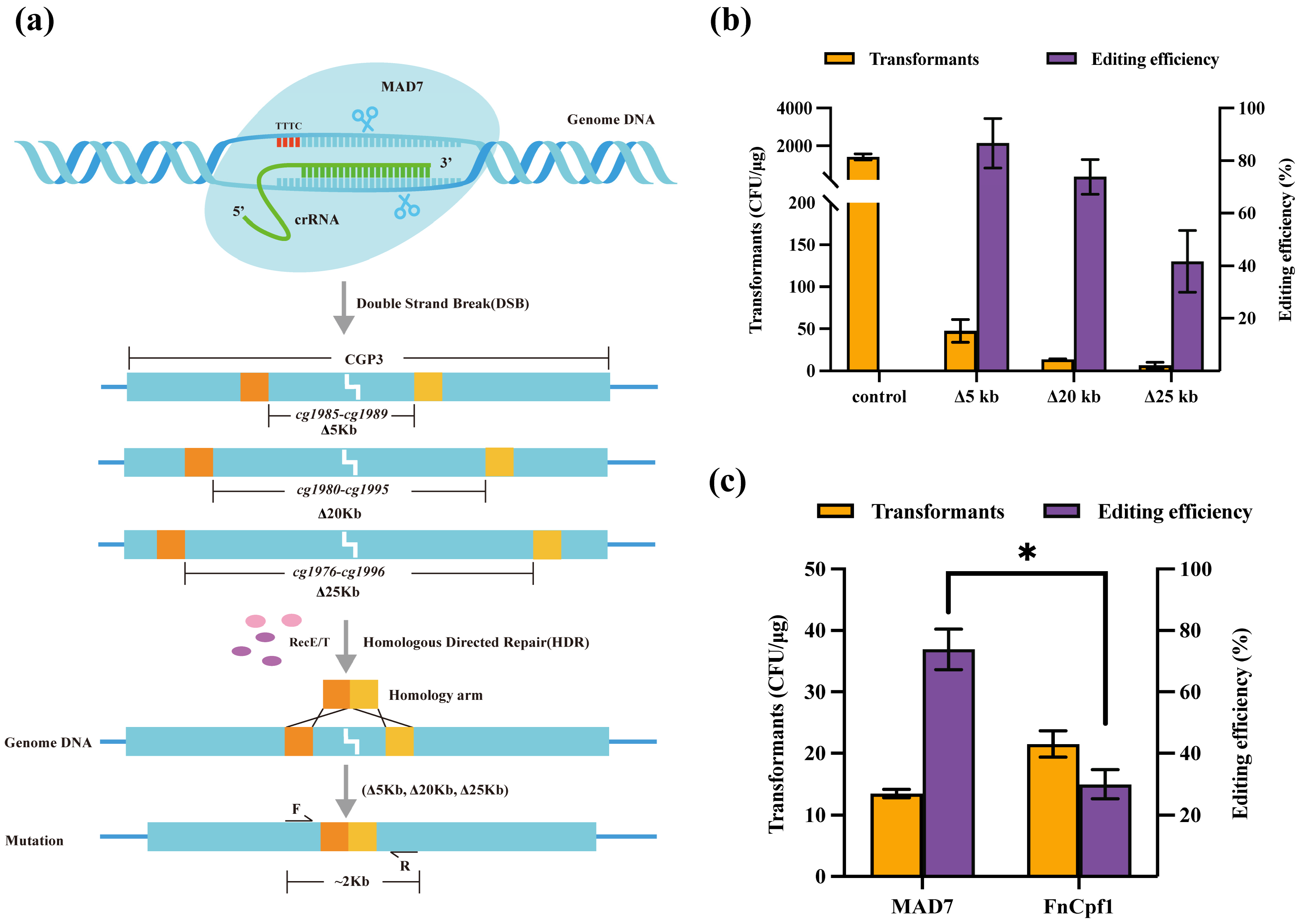
3.2. CRISPR/MAD7 System in Large DNA Fragment Deletion
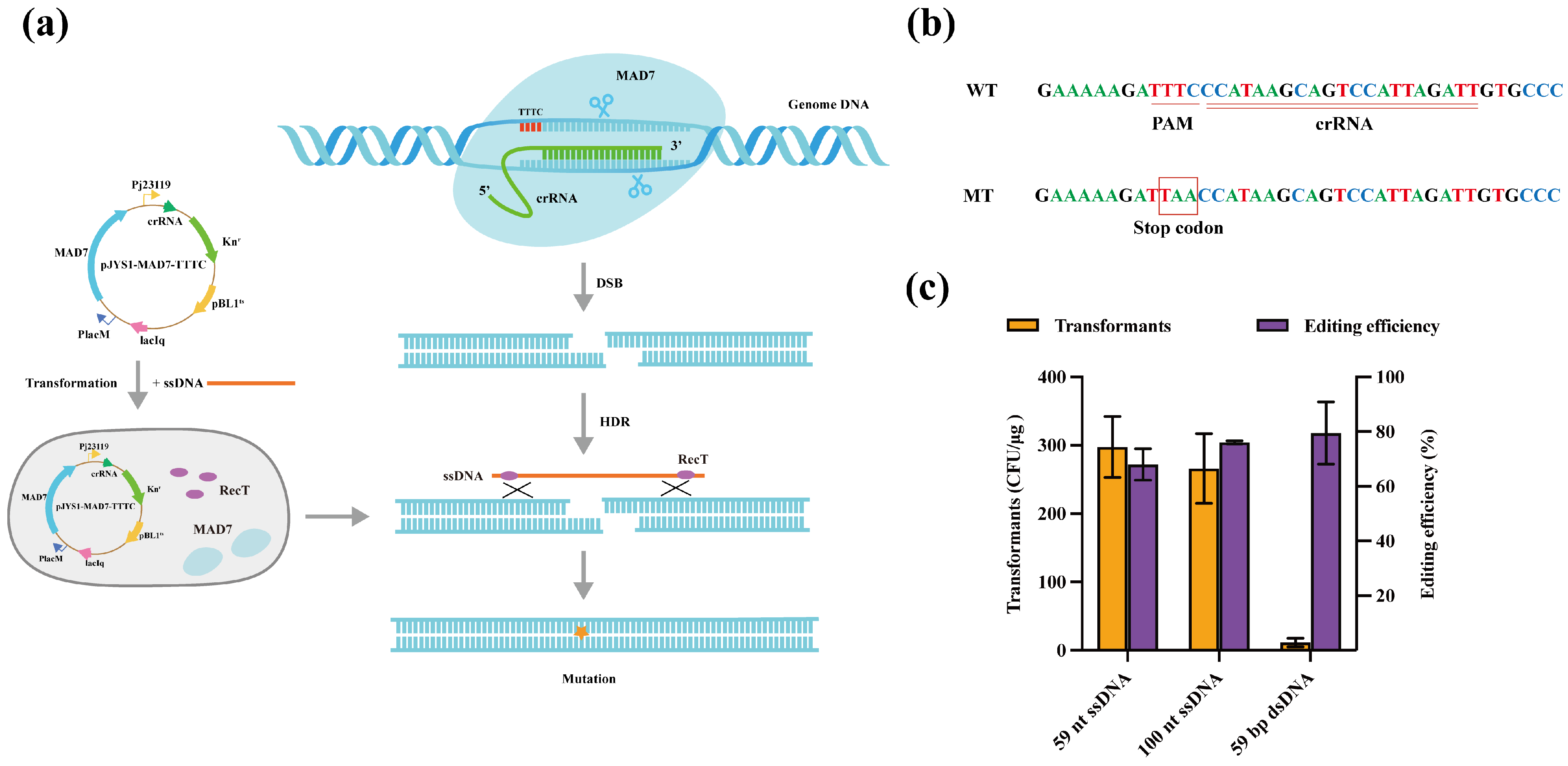
3.3. Editing Site Range by the CRISPR/MAD7 System
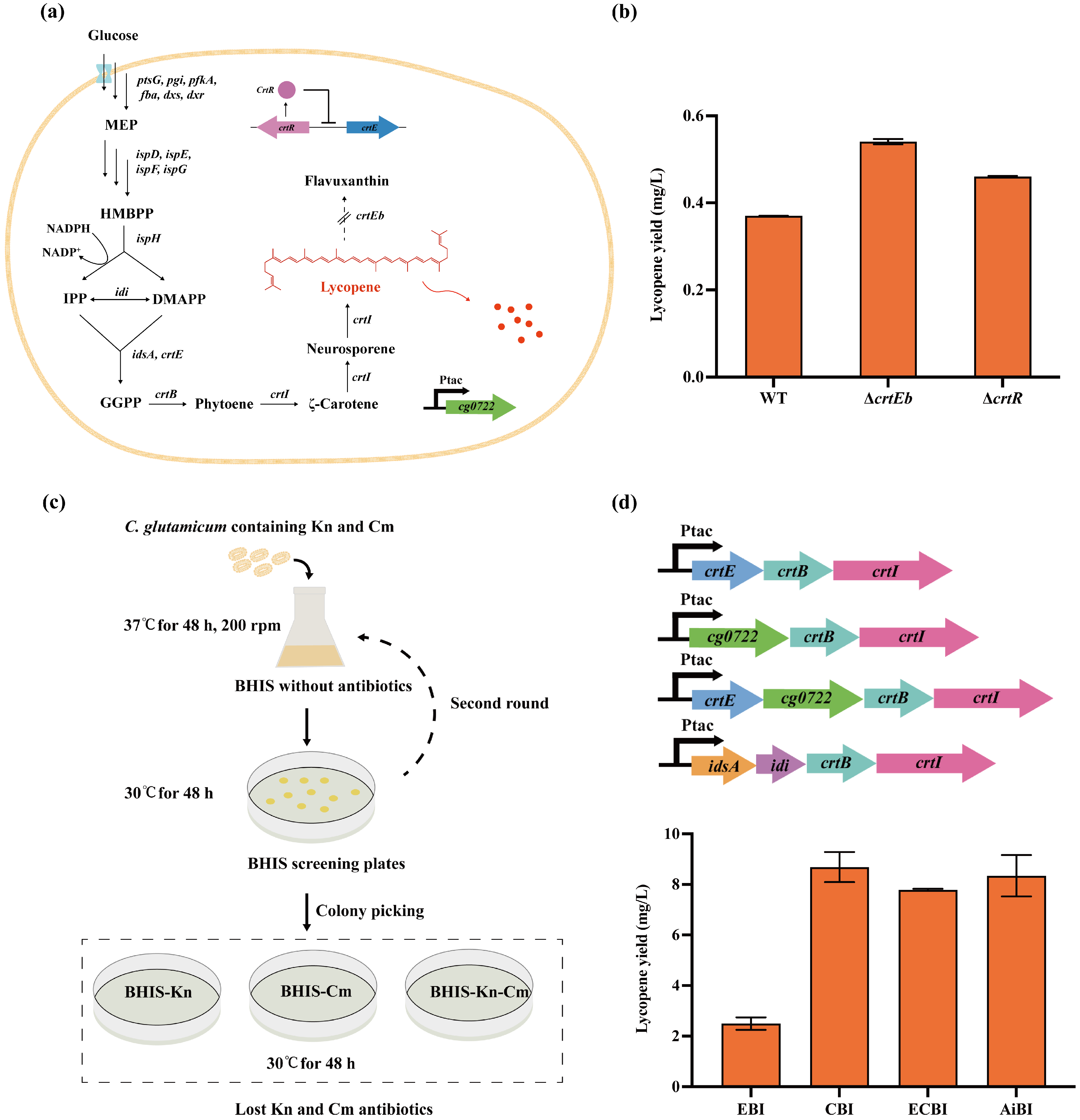
3.4. Metabolic Engineering of Lycopene Pathway Assisted by CRISPR/MAD7 System
3.5. High-Density Fermentation for Lycopene Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Cui, Z.; Hu, L. Recent technological strategies for enhancing the stability of lycopene in processing and production. Food Chem. 2023, 405, 134799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, D.; Duan, Y.; Zheng, X.; Lin, Y.; Liang, S. Production of lycopene by metabolically engineered Pichia pastoris. Biosci. Biotechnol. Biochem. 2019, 84, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Zhang, R.-R.; Yin, Y.; Tan, G.-F.; Wang, G.-L.; Liu, H.; Zhuang, J.; Zhang, J.; Zhuang, F.-Y.; Xiong, A.-S. Advances in engineering the production of the natural red pigment lycopene: A systematic review from a biotechnology perspective. J. Adv. Res. 2023, 46, 31–47. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Fleurat-Lessard, P.; Cruz, R.G.; Lafarge, C.; Grangeteau, C.; Yahou, F.; Gerbeau-Pissot, P.; Júnior, O.A.; Gervais, P.; Simon-Plas, F.; et al. Antioxidant properties of ergosterol and its role in yeast resistance to oxidation. Antioxidants 2021, 10, 1024. [Google Scholar] [CrossRef] [PubMed]
- Gottl, V.L.; Pucker, B.; Wendisch, V.F.; Henke, N.A. Screening of structurally distinct lycopene beta-cyclases for production of the cyclic c40 carotenoids beta-carotene and astaxanthin by Corynebacterium glutamicum. J. Agric. Food Chem. 2023, 71, 7765–7776. [Google Scholar] [CrossRef] [PubMed]
- Becker, J.; Rohles, C.M.; Wittmann, C. Metabolically engineered Corynebacterium glutamicum for bio-based production of chemicals, fuels, materials, and healthcare products. Metab. Eng. 2018, 50, 122–141. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Wang, T.; Bo, T.; Cai, N.; Yuan, M.; Wu, C.; Jiang, H.; Peng, H.; Chen, N.; Li, Y. Enhanced production of d-pantothenic acid in Corynebacterium glutamicum using an efficient CRISPR–Cpf1 genome editing method. Microb. Cell Factories 2023, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Matano, C.; Uhde, A.; Youn, J.-W.; Maeda, T.; Clermont, L.; Marin, K.; Krämer, R.; Wendisch, V.F.; Seibold, G.M. Engineering of Corynebacterium glutamicum for growth and l-lysine and lycopene production from n-acetyl-glucosamine. Appl. Microbiol. Biotechnol. 2014, 98, 5633–5643. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.; Peters-Wendisch, P.; Wendisch, V.F. Carotenoid biosynthesis and overproduction in Corynebacterium glutamicum. BMC Microbiol. 2012, 12, 198. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Swofford, C.A.; Rückert, C.; Chatzivasileiou, A.O.; Ou, R.W.; Opdensteinen, P.; Luttermann, T.; Zhou, K.; Stephanopoulos, G.; Prather, K.L.; et al. Heterologous production of alpha-carotene in Corynebacterium glutamicum using a multicopy chromosomal integration method. Bioresour. Technol. 2021, 341, 125782. [Google Scholar]
- Yamaryo-Botte, Y.; Rainczuk, A.K.; Lea-Smith, D.J.; Brammananth, R.; van der Peet, P.L.; Meikle, P.; Ralton, J.E.; Rupasinghe, T.W.; Williams, S.J.; Coppel, R.L. Acetylation of trehalose mycolates is required for efficient MmpL-mediated mem-brane transport in Corynebacterineae. ACS Chem. Biol. 2015, 10, 734–746. [Google Scholar] [CrossRef]
- Taniguchi, H.; Henke, N.A.; Heider, S.A.; Wendisch, V.F. Overexpression of the primary sigma factor gene sigA improved carotenoid production by Corynebacterium glutamicum: Application to production of beta-carotene and the non-native linear C50 carotenoid bisanhydrobacterioruberin. Metab. Eng. Commun. 2017, 4, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.E.; Wolf, N.; Hofemeier, A.; Peters-Wendisch, P.; Wendisch, V.F. Optimization of the IPP Precursor Supply for the Production of Lycopene, Decaprenoxanthin and Astaxanthin by Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2014, 2, 28. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and glucosylation of C50 and C40 carotenoids by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Henke, N.A.; Heider, S.A.; Peters-Wendisch, P.; Wendisch, V.F. Production of the Marine Carotenoid Astaxanthin by Metabolically Engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Lv, X.; Li, Y.; Xiu, X.; Liao, C.; Xu, Y.; Liu, Y.; Li, J.; Du, G.; Liu, L. CRISPR genetic toolkits of classical food microorganisms: Current state and future prospects. Biotechnol. Adv. 2023, 69, 108261. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Qian, F.; Yang, J.; Liu, Y.; Dong, F.; Xu, C.; Sun, B.; Chen, B.; Xu, X.; Li, Y.; et al. CRISPR-Cpf1 assisted genome editing of Corynebacterium glutamicum. Nat. Commun. 2017, 8, 15179. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Schiel, J.A.; Maksimova, E.; Strezoska, Ž.; Zhao, G.; Anderson, E.M.; Wu, Y.; Warren, J.; Bartels, A.; Smith, A.v.B.; et al. ErCas12a CRISPR-MAD7 for model generation in human cells, mice, and rats. CRISPR J. 2020, 3, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zhu, Z.; Liu, G.; Sun, C.; Lin, D.; Xue, C.; Li, S.; Zhang, D.; Gao, C.; Wang, Y.; et al. Genome editing in plants with MAD7 nuclease. J. Genet. Genom. 2021, 48, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, K.G.; Rendsvig, J.K.H.; Jarczynska, Z.D.; Cortes, M.V.d.C.B.; van Esch, A.P.; Morera-Gómez, M.; Contesini, F.J.; Mortensen, U.H. A Mad7 system for genetic engineering of Filamentous fungi. J. Fungi 2023, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- Mund, M.; Weber, W.; Degreif, D.; Schiklenk, C. A MAD7-based genome editing system for E. coli. Microb. Biotechnol. 2023, 16, 5. [Google Scholar] [CrossRef] [PubMed]
- Rojek, J.B.; Basavaraju, Y.; Nallapareddy, S.; Bulté, D.B.; Baumgartner, R.; Schoffelen, S.; Grav, L.M.; Goletz, S.; Pedersen, L. Expanding the CRISPR toolbox for Chinese Hamster Ovary cells with comprehensive tools for MAD7 genome editing. Biotechnol. Bioeng. 2023, 120, 1478–1491. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.; Peters-Wendisch, P.; Beekwilder, J.; Wendisch, V.F. IdsA is the major geranylgeranyl pyrophosphate synthase involved in carotenogenesis in Corynebacterium glutamicum. FEBS J. 2014, 281, 4906–4920. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Zhong, M.; Zhang, Z.; Chen, T.; Shen, Y.; Lin, Z.; Wang, Y. Genomic iterative replacements of large synthetic DNA fragments in Corynebacterium glutamicum. ACS Synth. Biol. 2022, 11, 1588–1599. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Cui, Y.; Du, L.; Liu, X.; Xie, X.; Chen, N. Identification and application of a growth-regulated promoter for improving l-valine production in Corynebacterium glutamicum. Microb. Cell Factories 2018, 17, 185. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Xu, D.; Li, Y.; Wang, X. Construction of a novel SacB-based system for marker-free gene deletion in Corynebacterium glutamicum. Plasmid 2012, 67, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Yuen, G.; Khan, F.J.; Gao, S.; Stommel, J.M.; Batchelor, E.; Wu, X.; Luo, J. CRISPR/Cas9-mediated gene knockout is insensitive to target copy number but is dependent on guide RNA potency and Cas9/sgRNA threshold expression level. Nucleic Acids Res. 2017, 45, 12039–12053. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Rittmann, D.; Singh, A.; Krumbach, K.; Bhatt, K.; Eggeling, L.; Besra, G.S.; Bhatt, A. Mmpl genes are associated with mycolic acid metabolism in Mycobacteria and Corynebacteria. Chem. Biol. 2012, 19, 498–506. [Google Scholar] [CrossRef]
- Zhao, N.; Li, L.; Luo, G.; Xie, S.; Lin, Y.; Han, S.; Huang, Y.; Zheng, S. Multiplex gene editing and large DNA fragment deletion by the CRISPR/Cpf1-RecE/T system in Corynebacterium glutamicum. J. Ind. Microbiol. Biotechnol. 2020, 47, 599–608. [Google Scholar] [CrossRef]
- Wang, B.; Hu, Q.; Zhang, Y.; Shi, R.; Chai, X.; Liu, Z.; Shang, X.; Zhang, Y.; Wen, T. A RecET-assisted CRISPR-Cas9 genome editing in Corynebacterium glutamicum. Microb. Cell Factories 2018, 17, 63. [Google Scholar] [CrossRef] [PubMed]
- Binder, S.; Siedler, S.; Marienhagen, J.; Bott, M.; Eggeling, L. Recombineering in Corynebacterium glutamicum combined with optical nanosensors: A general strategy for fast producer strain generation. Nucleic Acids Res. 2013, 41, 6360–6369. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Cheng, Y.; Ji, J.; Wang, C.; Wei, Y.; Wang, Y. Large chromosomal segment deletions by CRISPR/LbCpf1-mediated multiplex gene editing in soybean. J. Integr. Plant Biol. 2021, 63, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lin, Y.; Dai, Z.; Wang, Q. Modulating DNA repair pathways to diversify genomic alterations in saccharomyces cerevisiae. Microbiol. Spectr. 2022, 10, e0232621. [Google Scholar] [CrossRef] [PubMed]
- Henke, N.A.; Austermeier, S.; Grothaus, I.L.; Götker, S.; Persicke, M.; Peters-Wendisch, P.; Wendisch, V.F. Corynebacterium glutamicum CrtR and its orthologs in Actinobacteria: Conserved function and application as genetically encoded biosensor for detection of geranylgeranyl pyrophosphate. Int. J. Mol. Sci. 2020, 21, 5482. [Google Scholar] [CrossRef] [PubMed]
- Sumi, S.; Suzuki, Y.; Matsuki, T.; Yamamoto, T.; Tsuruta, Y.; Mise, K.; Kawamura, T.; Ito, Y.; Shimada, Y.; Watanabe, E.; et al. Light-inducible carotenoid production controlled by a marr-type regulator in Corynebacterium glutamicum. Sci. Rep. 2019, 9, 13136. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.; Zhang, C.; Zhang, W.; Xu, H.; Li, Y.; Wei, M.; Meng, J.; Meng, Y.; Wang, J.; Chen, N. A Myo-Inositol-Inducible expression system for Corynebacterium glutamicum and its application. Front. Bioeng. Biotechnol. 2021, 9, 746322. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, J.; Al Makishah, N.H.; Sun, X.; Wen, Z.; Jiang, Y.; Yang, S. Advances and perspectives for genome editing tools of Corynebacterium glutamicum. Front. Microbiol. 2021, 12, 654058. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, L.; Xie, S.; Zhao, N.; Han, S.; Lin, Y.; Zheng, S. Recombineering using RecET in Corynebacterium glutamicum ATCC 14067 via a self-excisable cassette. Sci. Rep. 2017, 7, 7916. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Lu, Y.; Zheng, P.; Sun, J.; Ma, Y. Development of a CRISPR/Cas9 genome editing toolbox for Corynebacterium glutamicum. Microb. Cell Factories 2017, 16, 205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, F.; Yang, Y.; Jiang, Y.; Huo, Y.X. Optimizing a CRISPR-Cpf1-based genome engineering system for Corynebacterium glutamicum. Microb. Cell Factories 2019, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Collias, D.; Vialetto, E.; Yu, J.; Co, K.; Almási, D.H.; Rüttiger, A.-S.; Achmedov, T.; Strowig, T.; Beisel, C.L. Systematically attenuating DNA targeting enables CRISPR-driven editing in bacteria. Nat. Commun. 2023, 14, 680. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strategy | Titer (mg/g DCW) | Reference |
|---|---|---|
| Deletion of crtEb and crtYe, expression of crtE, crtB, crtI from C. glutamicum by changing native promoter into the Ptac promoter | 4.56 | [10] |
| Deletion of crtEb, overexpression of crtE, crtB, crtI | 2.4 | [9] |
| Overexpression of sigA | 0.82 | [12] |
| Overexpression of idi | 0.08 | [13] |
| Deletion of crtEb and crtYe, expression of crtE, crtB, crtI | 0.79 | [14] |
| Deletion of crtEb, crtYe, and crtYf, expression of crtE, crtB, crtI, dxs from C. glutamicum using a Ptuf promoter | 0.5 | [15] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, Z.; Chen, X.; Ye, Z.; Zhao, M.; Li, C.; Gao, S.; Sinskey, A.J.; Yao, L.; Dai, J.; Jiang, Y.; et al. Expanding the CRISPR Toolbox for Engineering Lycopene Biosynthesis in Corynebacterium glutamicum. Microorganisms 2024, 12, 803. https://doi.org/10.3390/microorganisms12040803
Zhan Z, Chen X, Ye Z, Zhao M, Li C, Gao S, Sinskey AJ, Yao L, Dai J, Jiang Y, et al. Expanding the CRISPR Toolbox for Engineering Lycopene Biosynthesis in Corynebacterium glutamicum. Microorganisms. 2024; 12(4):803. https://doi.org/10.3390/microorganisms12040803
Chicago/Turabian StyleZhan, Zhimin, Xiong Chen, Zhifang Ye, Ming Zhao, Cheng Li, Shipeng Gao, Anthony J. Sinskey, Lan Yao, Jun Dai, Yiming Jiang, and et al. 2024. "Expanding the CRISPR Toolbox for Engineering Lycopene Biosynthesis in Corynebacterium glutamicum" Microorganisms 12, no. 4: 803. https://doi.org/10.3390/microorganisms12040803