Associations of Fecal Microbiota with Ectopic Fat in African Caribbean Men

 , ,
, ,
Abstract
:1. Introduction
2. Methods
2.1. Study Population
2.2. Fecal Sample Collection and Processing
2.3. Computed Tomography Assessments
2.4. Other Variables
2.5. Statistical Methods
2.5.1. Diversity Analyses
- Model 1: unadjusted (with the exception that models with observed OTUs as the independent variable additionally adjusted for unrarefied sequencing depth);
- Model 2: Model 1 + adjustment for age (years), education status (categorical), current smoking status (yes/no), drinking 4+ alcoholic beverages per week (yes/no), hours walked per week for exercise (hours), and the time difference between CT scan and fecal sample collection (years).
2.5.2. Compositional Analyses
2.5.3. Sensitivity Analyses
3. Results
3.1. Sample Characteristics
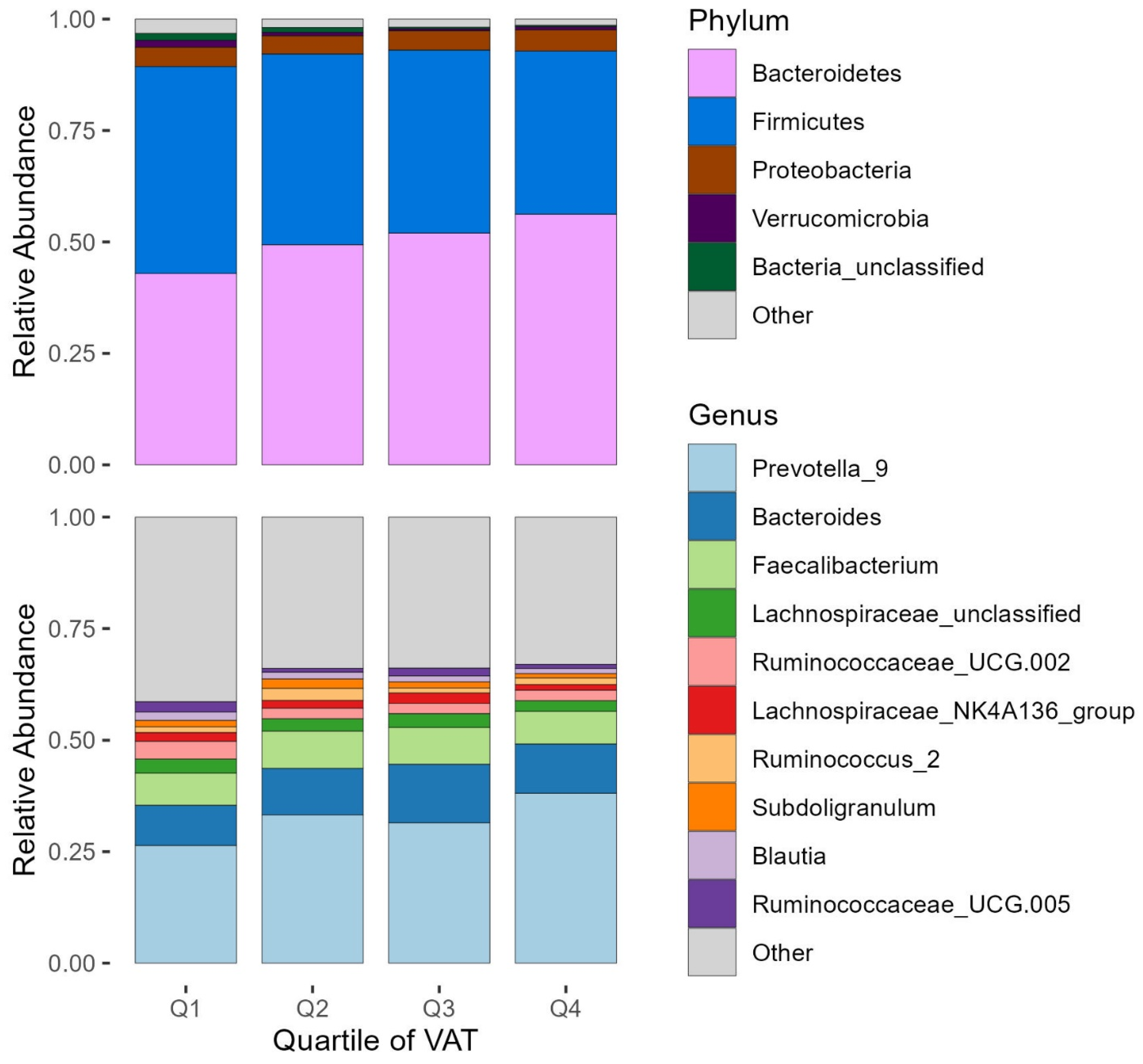
3.2. Microbiota Characteristics
3.3. Diversity Analyses
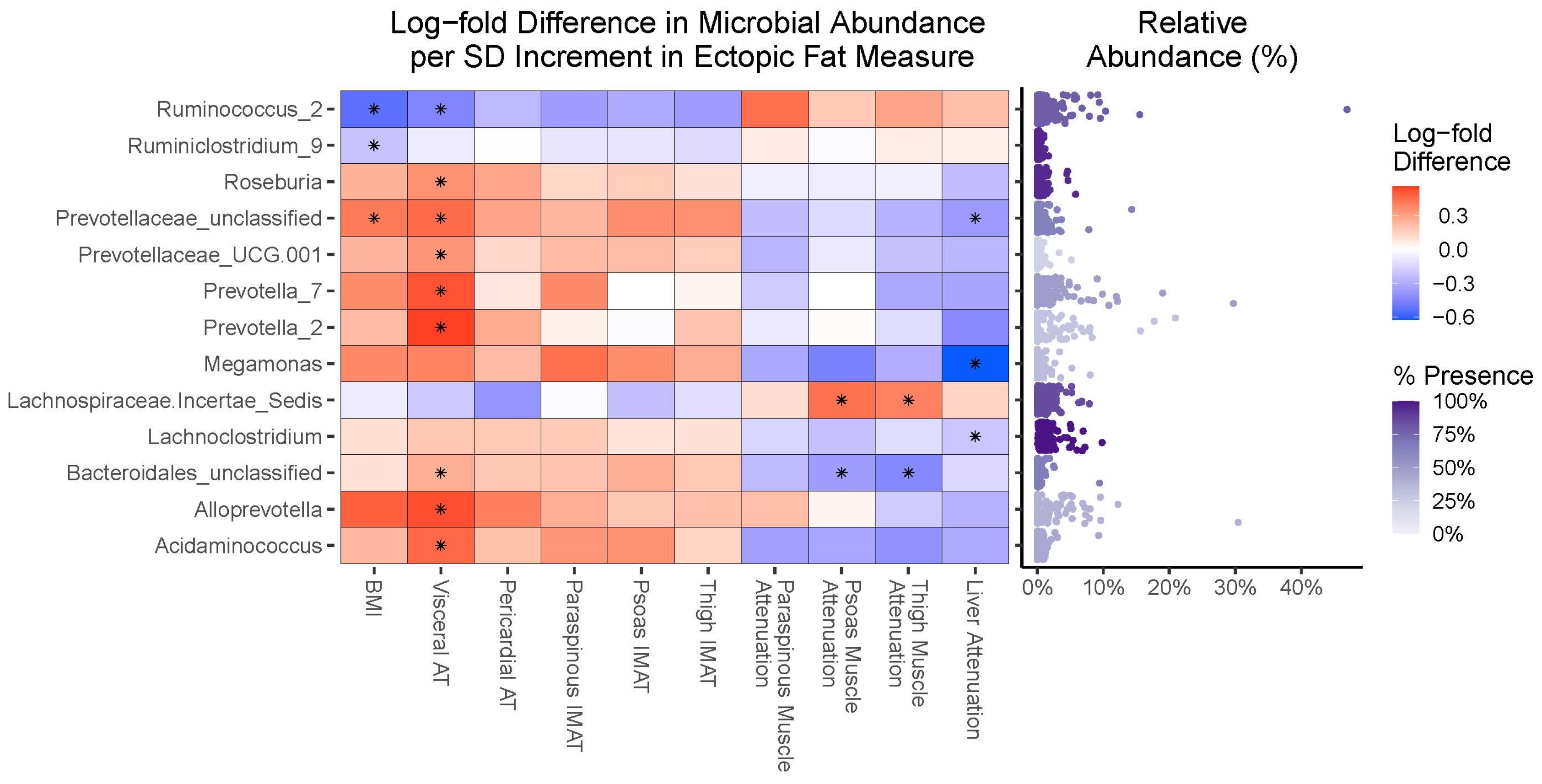
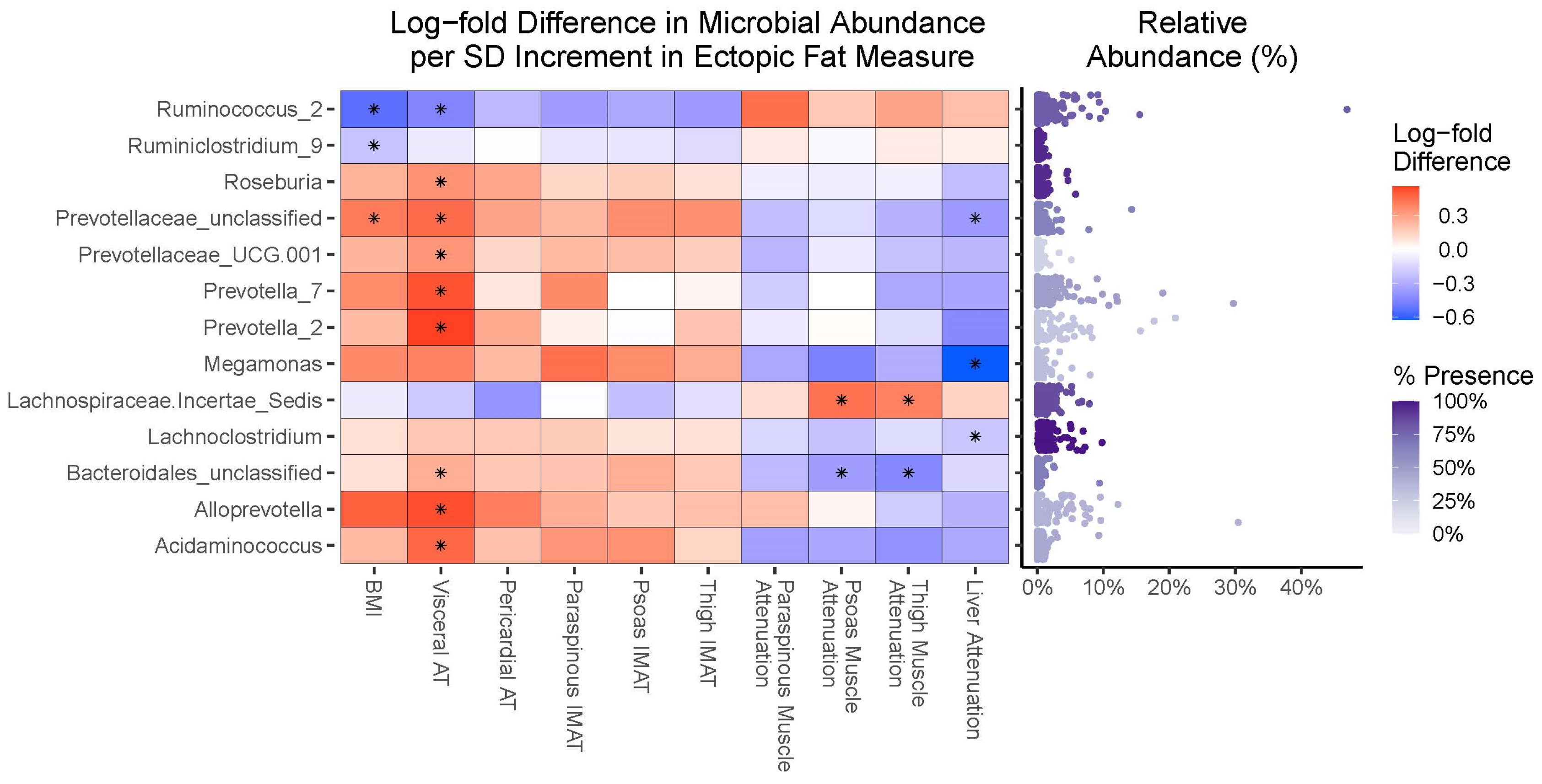
3.4. Compositional Analyses
3.5. Sensitivity Analyses
4. Discussion
4.1. The Importance of Race/Ethnicity and Geography in Microbiota–Obesity Relationships
4.2. Fecal Microbiota Diversity Is Associated with Some, but Not All, Ectopic Fats
4.3. Fecal Microbiota Taxa Are Similarly Associated with Overall Obesity and Ectopic Fat Accumulation
4.4. Perspectives and Implications for Future Research
4.5. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neeland, I.J.; Ross, R.; Després, J.P.; Matsuzawa, Y.; Yamashita, S.; Shai, I.; Seidell, J.; Magni, P.; Santos, R.D.; Arsenault, B.; et al. Visceral and ectopic fat, atherosclerosis, and cardiometabolic disease: A position statement. Lancet Diabetes Endocrinol. 2019, 7, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Chartrand, D.J.; Murphy-Després, A.; Alméras, N.; Lemieux, I.; Larose, E.; Després, J.-P. Overweight, Obesity, and CVD Risk: A Focus on Visceral/Ectopic Fat. Curr. Atheroscler. Rep. 2022, 24, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.; Bellettiere, J.; Allison, M.; McClelland, R.L.; Miljkovic, I.; Vella, C.A.; Ouyang, P.; De-Guzman, K.R.; Criqui, M.; Unkart, J. Muscle area and density and risk of all-cause mortality: The Multi-Ethnic Study of Atherosclerosis. Metabolism 2020, 111, 154321. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.O.; Sidney, S.; Lewis, C.E.; Tolan, K.; Scherzinger, A.L.; Stamm, E.R. Racial differences in amounts of visceral adipose tissue in young adults: The CARDIA (Coronary Artery Risk Development in Young Adults) Study. Am. J. Clin. Nutr. 1999, 69, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.J.; Wang, Z.; Gallagher, D.; Heymsfield, S.B. Comparison of visceral adipose tissue mass in adult African Americans and whites. Obes. Res. 2005, 13, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, R.; Vega, G.L.; Grundy, S.M.; Browning, J.D. Ethnic differences in hepatic steatosis: An insulin resistance paradox? J. Hepatol. 2008, 49, 791–801. [Google Scholar] [CrossRef]
- Gallagher, D.; Kuznia, P.; Heshka, S.; Albu, J.; Heymsfield, S.B.; Goodpaster, B.; Visser, M.; Harris, T.B. Adipose tissue in muscle: A novel depot similar in size to visceral adipose tissue. Am. J. Clin. Nutr. 2005, 81, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Albu, J.B.; Kovera, A.J.; Allen, L.; Wainwright, M.; Berk, E.; Raja-Khan, N.; Janumala, I.; Burkey, B.; Heshka, S.; Gallagher, D. Independent association of insulin resistance with larger amounts of intermuscular adipose tissue and a greater acute insulin response to glucose in African American than in white nondiabetic women. Am. J. Clin. Nutr. 2005, 82, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Maruvada, P.; Leone, V.; Kaplan, L.M.; Chang, E.B. The Human Microbiome and Obesity: Moving beyond Associations. Cell Host Microbe 2017, 22, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.D. The Gut Microbiome and Its Role in Obesity. Nutr. Today 2016, 51, 167–174. [Google Scholar] [CrossRef]
- Tilves, C.; Tanaka, T.; Differding, M.K.; Spira, A.P.; Chia, C.W.; Ferrucci, L.; Mueller, N.T. The gut microbiome and regional fat distribution: Findings from the Baltimore Longitudinal Study of Aging. Obesity 2023, 31, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Al Khatib, H.; et al. Microbiome connections with host metabolism and habitual diet from 1,098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Goodrich, J.K.; Jackson, M.A.; Yet, I.; Davenport, E.R.; Vieira-Silva, S.; Debelius, J.; Pallister, T.; Mangino, M.; Raes, J.; et al. Heritable components of the human fecal microbiome are associated with visceral fat. Genome Biol. 2016, 17, 189. [Google Scholar] [CrossRef] [PubMed]
- Mallott, E.K.; Sitarik, A.R.; Leve, L.D.; Cioffi, C.; Camargo, C.A.; Hasegawa, K.; Bordenstein, S.R. Human microbiome variation associated with race and ethnicity emerges as early as 3 months of age. PLoS Biol. 2023, 21, e3002230. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.W.; Priya, S.; Blekhman, R.; Bordenstein, S.R. Gut microbiota diversity across ethnicities in the United States. PLoS Biol. 2018, 16, e2006842. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; George Markowitz, R.H.; Brooks, A.W.; Mallott, E.K.; Leigh, B.A.; Olszewski, T.; Zare, H.; Bagheri, M.; Smith, H.M.; Friese, K.A.; et al. Individuality and ethnicity eclipse a short-term dietary intervention in shaping microbiomes and viromes. PLoS Biol. 2022, 20, e3001758. [Google Scholar] [CrossRef] [PubMed]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef] [PubMed]
- Ecklu-Mensah, G.; Choo-Kang, C.; Maseng, M.G.; Donato, S.; Bovet, P.; Viswanathan, B.; Bedu-Addo, K.; Plange-Rhule, J.; Boateng, P.O.; Forrester, T.E.; et al. Gut microbiota and fecal short chain fatty acids differ with adiposity and country of origin: The METS-microbiome study. Nat. Commun. 2023, 14, 5160. [Google Scholar] [CrossRef] [PubMed]
- Hullar, M.A.J.; Jenkins, I.C.; Randolph, T.W.; Curtis, K.R.; Monroe, K.R.; Ernst, T.; Shepherd, J.A.; Stram, D.O.; Cheng, I.; Kristal, B.S.; et al. Associations of the gut microbiome with hepatic adiposity in the Multiethnic Cohort Adiposity Phenotype Study. Gut Microbes 2021, 13, 1965463. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, I.; Cauley, J.A.; Petit, M.A.; Ensrud, K.E.; Strotmeyer, E.; Sheu, Y.; Gordon, C.L.; Goodpaster, B.H.; Bunker, C.H.; Patrick, A.L.; et al. Greater adipose tissue infiltration in skeletal muscle among older men of African ancestry. J. Clin. Endocrinol. Metab. 2009, 94, 2735–2742. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic-Gacic, I.; Ferrell, R.E.; Patrick, A.L.; Kammerer, C.M.; Bunker, C.H. Estimates of African, European and Native American ancestry in Afro-Caribbean men on the island of Tobago. Hum. Hered. 2005, 60, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Bunker, C.H.; Patrick, A.L.; Konety, B.R.; Dhir, R.; Brufsky, A.M.; Vivas, C.A.; Becich, M.J.; Trump, D.L.; Kuller, L.H. High Prevalence of Screening-detected Prostate Cancer among Afro-Caribbeans: The Tobago Prostate Cancer Survey. Cancer Epidemiol. Biomark. Prev. 2002, 11, 726–729. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Terry, J.G.; Shay, C.M.; Schreiner, P.J.; Jacobs, D.R.; Sanchez, O.A.; Reis, J.P.; Goff, D.C.; Gidding, S.S.; Steffen, L.M.; Carr, J.J. Intermuscular Adipose Tissue and Subclinical Coronary Artery Calcification in Midlife: The CARDIA Study (Coronary Artery Risk Development in Young Adults). Arter. Thromb. Vasc. Biol. 2017, 37, 2370–2378. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.E.; Carr, J.J.; Nair, S.; Terry, J.G.; Reed, R.D.; Smith, G.D.; Segev, D.L.; Kumar, V.; Lewis, C.E. Abdominal lean muscle is associated with lower mortality among kidney waitlist candidates. Clin. Transplant. 2017, 31, e12911. [Google Scholar] [CrossRef] [PubMed]
- Granados, A.; Gebremariam, A.; Gidding, S.S.; Terry, J.G.; Carr, J.J.; Steffen, L.M.; Jacobs, D.R.; Lee, J.M. Association of abdominal muscle composition with prediabetes and diabetes: The CARDIA study. Diabetes Obes. Metab. 2019, 21, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Miljkovic, I.; Kuipers, A.L.; Cvejkus, R.K.; Carr, J.J.; Terry, J.G.; Thyagarajan, B.; Wheeler, V.W.; Nair, S.; Zmuda, J.M. Hepatic and Skeletal Muscle Adiposity Are Associated with Diabetes Independent of Visceral Adiposity in Nonobese African-Caribbean Men. Metab. Syndr. Relat. Disord. 2020, 18, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Ramdath, D.D.; Hilaire, D.G.; Cheong, K.D.; Sharma, S. Dietary intake among adults in Trinidad and Tobago and development of a quantitative food frequency questionnaire to highlight nutritional needs for lifestyle interventions. Int. J. Food Sci. Nutr. 2011, 62, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.T.; McCullough, M.L.; Newby, P.; Manson, J.E.; Meigs, J.B.; Rifai, N.; Willett, W.C.; Hu, F.B. Diet-quality scores and plasma concentrations of markers of inflammation and endothelial dysfunction. Am. J. Clin. Nutr. 2005, 82, 163–173. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package Version 2.5-7; Scientific Research Publishing: Wuhan, China, 2020. [Google Scholar]
- Lin, H.; Peddada, S.D. Analysis of compositions of microbiomes with bias correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Satten, G.A.; Hu, Y.-J. LOCOM: A logistic regression model for testing differential abundance in compositional microbiome data with false discovery rate control. Proc. Natl. Acad. Sci. USA 2022, 119, e2122788119. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the analysis of high-throughput sequencing datasets: Characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, C.; Monteil, M.; Davis, M. Overweight and Obesity in Children Are Associated with an Abundance of Firmicutes and Reduction of Bifidobacterium in Their Gastrointestinal Microbiota. Child. Obes. 2020, 16, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Sze, M.A.; Schloss, P.D. Looking for a Signal in the Noise: Revisiting Obesity and the Microbiome. mBio 2016, 7, e01018-16. [Google Scholar] [CrossRef] [PubMed]
- Pinart, M.; Dötsch, A.; Schlicht, K.; Laudes, M.; Bouwman, J.; Forslund, S.K.; Pischon, T.; Nimptsch, K. Gut Microbiome Composition in Obese and Non-Obese Persons: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.; Ma, X.; Sankaran, K.; Ru, Y.; Chen, L.; Baiocchi, M.; Zhu, S. Sex-specific association between gut microbiome and fat distribution. Nat. Commun. 2019, 10, 2408. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered Fecal Microbiota Correlates with Liver Biochemistry in Nonobese Patients with Non-alcoholic Fatty Liver Disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef] [PubMed]
- Ruuskanen, M.O.; Åberg, F.; Männistö, V.; Havulinna, A.S.; Méric, G.; Liu, Y.; Loomba, R.; Vázquez-Baeza, Y.; Tripathi, A.; Valsta, L.M.; et al. Links between gut microbiome composition and fatty liver disease in a large population sample. Gut Microbes 2021, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Correa-De-Araujo, R.; Addison, O.; Miljkovic, I.; Goodpaster, B.H.; Bergman, B.C.; Clark, R.V.; Elena, J.W.; Esser, K.A.; Ferrucci, L.; Harris-Love, M.O.; et al. Myosteatosis in the Context of Skeletal Muscle Function Deficit: An Interdisciplinary Workshop at the National Institute on Aging. Front. Physiol. 2020, 11, 963. [Google Scholar] [CrossRef] [PubMed]
- Lanthier, N.; Rodriguez, J.; Nachit, M.; Hiel, S.; Trefois, P.; Neyrinck, A.M.; Cani, P.D.; Bindels, L.B.; Thissen, J.-P.; Delzenne, N.M. Microbiota analysis and transient elastography reveal new extra-hepatic components of liver steatosis and fibrosis in obese patients. Sci. Rep. 2021, 11, 659. [Google Scholar] [CrossRef] [PubMed]
- Rosito, G.A.; Massaro, J.M.; Hoffmann, U.; Ruberg, F.L.; Mahabadi, A.A.; Vasan, R.S.; O’Donnell, C.J.; Fox, C.S. Pericardial Fat, Visceral Abdominal Fat, Cardiovascular Disease Risk Factors, and Vascular Calcification in a Community-Based Sample. Circulation 2008, 117, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Ching, Y.H.; Li, Y.P.; Liu, J.Y.; Huang, Y.T.; Huang, Y.W.; Yang, S.S.; Huang, W.C.; Chuang, H.L. Nonalcoholic Fatty Liver Disease Is Exacerbated in High-Fat Diet-Fed Gnotobiotic Mice by Colonization with the Gut Microbiota from Patients with Nonalcoholic Steatohepatitis. Nutrients 2017, 9, 1220. [Google Scholar] [CrossRef] [PubMed]
- Hoyles, L.; Fernández-Real, J.M.; Federici, M.; Serino, M.; Abbott, J.; Charpentier, J.; Heymes, C.; Luque, J.L.; Anthony, E.; Barton, R.H.; et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 2018, 24, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Frost, F.; Kacprowski, T.; Rühlemann, M.; Pietzner, M.; Bang, C.; Franke, A.; Nauck, M.; Völker, U.; Völzke, H.; Dörr, M.; et al. Long-term instability of the intestinal microbiome is associated with metabolic liver disease, low microbiota diversity, diabetes mellitus and impaired exocrine pancreatic function. Gut 2021, 70, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, S.R.; Ajmera, V.; Loomba, R. Emerging Role of the Gut Microbiome in Nonalcoholic Fatty Liver Disease: From Composition to Function. Clin. Gastroenterol. Hepatol. 2019, 17, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.O.; Bäckhed, F. Signals from the gut microbiota to distant organs in physiology and disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Usyk, M.; Vázquez-Baeza, Y.; Chen, G.-C.; Isasi, C.R.; Williams-Nguyen, J.S.; Hua, S.; McDonald, D.; Thyagarajan, B.; Daviglus, M.L.; et al. Microbial co-occurrence complicates associations of gut microbiome with US immigration, dietary intake and obesity. Genome Biol. 2021, 22, 336. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pasolli, E.; Tett, A.; Tarallo, S.; Naccarati, A.; De Angelis, M.; Neviani, E.; Cocolin, L.; Gobbetti, M.; Segata, N.; et al. Distinct Genetic and Functional Traits of Human Intestinal Prevotella copri Strains Are Associated with Different Habitual Diets. Cell Host Microbe 2019, 25, 444–453.e3. [Google Scholar] [CrossRef] [PubMed]
- Palmas, V.; Pisanu, S.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Loviselli, A.; Manzin, A.; et al. Gut microbiota markers associated with obesity and overweight in Italian adults. Sci. Rep. 2021, 11, 5532. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.W.F.; Kersten, S. The role of the gut microbiota in metabolic health. FASEB J. 2015, 29, 3111–3123. [Google Scholar] [CrossRef] [PubMed]
- Tilves, C.M.; Zmuda, J.M.; Kuipers, A.L.; Nestlerode, C.S.; Evans, R.W.; Bunker, C.H.; Patrick, A.L.; Miljkovic, I. Association of Lipopolysaccharide-Binding Protein with Aging-Related Adiposity Change and Prediabetes Among African Ancestry Men. Diabetes Care 2016, 39, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Fusco, W.; Lorenzo, M.B.; Cintoni, M.; Porcari, S.; Rinninella, E.; Kaitsas, F.; Lener, E.; Mele, M.C.; Gasbarrini, A.; Collado, M.C.; et al. Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients 2023, 15, 2211. [Google Scholar] [CrossRef] [PubMed]
- Lozano, C.P.; Wilkens, L.R.; Shvetsov, Y.B.; Maskarinec, G.; Park, S.-Y.; Shepherd, J.A.; Boushey, C.J.; Hebert, J.R.; Wirth, M.D.; Ernst, T.; et al. Associations of the Dietary Inflammatory Index with total adiposity and ectopic fat through the gut microbiota, LPS, and C-reactive protein in the Multiethnic Cohort–Adiposity Phenotype Study. Am. J. Clin. Nutr. 2021, 115, 1344–1356. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.-Y.; Huang, F.-Q.; Lao, X.; Lu, Y.; Gao, X.; Alolga, R.N.; Yin, K.; Zhou, X.; Wang, Y.; Liu, B.; et al. Integrated metagenomics identifies a crucial role for trimethylamine-producing Lachnoclostridium in promoting atherosclerosis. NPJ Biofilms Microbiomes 2022, 8, 11. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; DuGar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Masi, C.; Manson, J.E.; Rexrode, K.M.; Qi, L. Gut Microbiota Metabolites and Risk of Major Adverse Cardiovascular Disease Events and Death: A Systematic Review and Meta-Analysis of Prospective Studies. J. Am. Hear. Assoc. 2017, 6, e004947. [Google Scholar] [CrossRef] [PubMed]
- Schiattarella, G.G.; Sannino, A.; Toscano, E.; Giugliano, G.; Gargiulo, G.; Franzone, A.; Trimarco, B.; Esposito, G.; Perrino, C. Gut microbe-generated metabolite trimethylamine-N-oxide as cardiovascular risk biomarker: A systematic review and dose-response meta-analysis. Eur. Heart J. 2017, 38, 2948–2956. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Liu, Y.; Zhou, R.F.; Chen, X.-L.; Wang, C.; Tan, X.-Y.; Wang, L.-J.; Zheng, R.-D.; Zhang, H.-W.; Ling, W.-H.; et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci. Rep. 2016, 6, 19076. [Google Scholar] [CrossRef] [PubMed]
- De Wolfe, T.J.; Arefin, M.R.; Benezra, A.; Gómez, M.R. Chasing Ghosts: Race, Racism, and the Future of Microbiome Research. mSystems 2021, 6, e0060421. [Google Scholar] [CrossRef] [PubMed]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R.; Arrieta, M.-C.; Azad, M.B.; Bailey, M.T.; Broussard, J.L.; Bruggeling, C.E.; Claud, E.C.; Costello, E.K.; Davenport, E.R.; Dutilh, B.E.; et al. The human gut microbiome and health inequities. Proc. Natl. Acad. Sci. USA 2021, 118, e2017947118. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.R.; Davenport, E.R.; Gautam, Y.; Bhandari, D.; Tandukar, S.; Ng, K.M.; Fragiadakis, G.K.; Holmes, S.; Gautam, G.P.; Leach, J.; et al. Gut microbiome transition across a lifestyle gradient in Himalaya. PLoS Biol. 2018, 16, e2005396. [Google Scholar] [CrossRef] [PubMed]
- Bowyer, R.C.E.; Jackson, M.A.; Le Roy, C.I.; Ni Lochlainn, M.; Spector, T.D.; Dowd, J.B.; Steves, C.J. Socioeconomic Status and the Gut Microbiome: A TwinsUK Cohort Study. Microorganisms 2019, 7, 17. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Overall (n = 193) | VAT Q1 (n = 49) | VAT Q2 (n = 48) | VAT Q3 (n = 48) | VAT Q4 (n = 48) | Linear Trend p-Value | |
|---|---|---|---|---|---|---|
| Age (years) | 60.0 [56.0, 68.0] | 60.0 [56.0, 68.0] | 58.5 [55.0, 63.3] | 60.0 [55.0, 70.0] | 61.0 [57.8, 68.3] | 0.280 |
| Education | ||||||
| Primary | 147 (76.2%) | 41 (83.7%) | 32 (66.7%) | 39 (81.3%) | 35 (72.9%) | 0.072 |
| Secondary | 28 (14.5%) | 2 (4.1%) | 9 (18.8%) | 6 (12.5%) | 11 (22.9%) | |
| Post-Secondary | 18 (9.3%) | 6 (12.2%) | 7 (14.6%) | 3 (6.3%) | 2 (4.2%) | |
| Hours Walked/Week | 2.5 [0.0, 6.0] | 2.0 [0.0, 5.3] | 2.3 [0.0, 6.3] | 2.5 [0.0, 5.3] | 2.8 [0.0, 6.0] | 0.858 |
| BMI (kg/m2) | 28.3 (4.9) | 24.2 (3.1) | 27.7 (3.2) | 29.1 (4.0) | 32.2 (5.2) | <0.001 |
| Current Smoking Status | 18 (9.3%) | 9 (18.4%) | 5 (10.4%) | 2 (4.2%) | 2 (4.2%) | 0.051 |
| Has 4+ Alcoholic Drinks/Week | 25 (13.0%) | 4 (8.2%) | 7 (14.6%) | 7 (14.6%) | 7 (14.6%) | 0.722 |
| Time Difference between Measures (years) | 2.5 [1.6, 2.6] | 2.5 [1.6, 2.8] | 2.5 [1.7, 2.7] | 2.5 [1.7, 2.6] | 1.7 [1.6, 2.5] | 0.137 |
| Fat Measures | ||||||
| Abdominal VAT (cm3) | 93.0 [55.7, 124.6] | 39.2 [22.9, 48.7] | 70.6 [63.4, 80.6] | 111.2 [105.7, 118.1] | 150.9 [136.1, 172.3] | <0.001 |
| Pericardial Fat (cm3) | 30.0 [19.3, 46.4] | 18.3 [12.7, 25.7] | 26.6 [18.4, 37.5] | 34.0 [22.3, 44.6] | 48.3 [40.5, 62.7] | <0.001 |
| Psoas IMAT (cm3) | 0.6 [0.4, 0.9] | 0.4 [0.2, 0.6] | 0.6 [0.5, 0.8] | 0.7 [0.5, 0.9] | 0.9 [0.6, 1.2] | <0.001 |
| Paraspinous IMAT (cm3) | 2.4 [1.7, 3.4] | 1.7 [1.0, 2.3] | 2.4 [1.8, 2.9] | 2.5 [2.1, 3.9] | 3.2 [2.4, 4.2] | <0.001 |
| Thigh IMAT (cm3) | 105.6 [83.8, 134.6] | 69.7 [42.3, 95.8] | 103.7 [88.2, 128.7] | 111.4 [95.6, 131.3] | 139.9 [106.9, 182.8] | <0.001 |
| Psoas Muscle Attenuation (HU) | 49.2 [46.1, 51.3] | 50.9 [49.1, 52.4] | 49.6 [47.9, 51.4] | 48.5 [45.5, 51.0] | 47.1 [44.7, 50.0] | <0.001 |
| Paraspinous Muscle Attenuation (HU) | 44.8 [39.9, 48.6] | 47.9 [44.8, 50.3] | 45.3 [41.9, 48.5] | 44.1 [37.4, 48.2] | 42.9 [37.3, 45.3] | <0.001 |
| Thigh Muscle Attenuation (HU) | 43.9 [41.1, 45.9] | 45.4 [43.1, 46.8] | 44.8 [42.5, 46.4] | 42.8 [40.9, 45.1] | 41.3 [39.7, 44.0] | <0.001 |
| Liver Attenuation (HU) | 57.5 [53.4, 61.5] | 61.3 [58.6, 63.3] | 59.1 [55.3, 61.2] | 56.5 [53.6, 60.3] | 52.3 [45.2, 56.5] | <0.001 |
| Alpha Diversity Measures | ||||||
| Observed OTUs | 60.3 (13.9) | 61.3 (13.5) | 59.4 (12.6) | 61.2 (14.9) | 59.1 (14.7) | 0.589 |
| Pielou’s Evenness | 0.6 [0.5, 0.7] | 0.7 [0.6, 0.7] | 0.6 [0.5, 0.7] | 0.6 [0.5, 0.7] | 0.6 [0.5, 0.7] | 0.019 |
| Shannon Diversity | 2.5 (0.7) | 2.6 (0.6) | 2.5 (0.7) | 2.5 (0.6) | 2.3 (0.7) | 0.037 |
| BMI or Fat Measure | Model | Observed OTUs | Pielou’s Evenness | Shannon Diversity Index |
|---|---|---|---|---|
| BMI | 1 | −0.06 (−0.21, 0.08) | −0.19 (−0.33, −0.04) | −0.17 (−0.31, −0.03) |
| 2 | −0.02 (−0.16, 0.12) | −0.16 (−0.30, −0.02) | −0.14 (−0.27, 0.00) | |
| VAT | 1 | −0.04 (−0.18, 0.11) | −0.16 (−0.30, −0.02) | −0.14 (−0.28, 0.00) |
| 2 | −0.06 (−0.21, 0.09) | −0.17 (−0.31, −0.04) | −0.15 (−0.29, −0.02) | |
| Pericardial fat | 1 | −0.01 (−0.16, 0.13) | −0.11 (−0.25, 0.03) | −0.10 (−0.24, 0.05) |
| 2 | −0.03 (−0.18, 0.11) | −0.12 (−0.27, 0.02) | −0.11 (−0.25, 0.03) | |
| Paraspinous IMAT | 1 | 0.04 (−0.11, 0.18) | −0.04 (−0.19, 0.10) | −0.03 (−0.17, 0.12) |
| 2 | −0.03 (−0.17, 0.12) | −0.09 (−0.23, 0.05) | −0.08 (−0.22, 0.06) | |
| Psoas IMAT | 1 | −0.01 (−0.15, 0.14) | −0.07 (−0.22, 0.07) | −0.06 (−0.20, 0.08) |
| 2 | −0.03 (−0.17, 0.11) | −0.08 (−0.22, 0.06) | −0.07 (−0.21, 0.07) | |
| Thigh IMAT | 1 | −0.04 (−0.18, 0.10) | −0.09 (−0.23, 0.05) | −0.08 (−0.22, 0.06) |
| 2 | −0.04 (−0.19, 0.10) | −0.09 (−0.24, 0.05) | −0.08 (−0.23, 0.06) | |
| Paraspinous muscle attenuation | 1 | −0.06 (−0.20, 0.08) | −0.03 (−0.17, 0.11) | −0.04 (−0.18, 0.10) |
| 2 | 0.04 (−0.09, 0.16) | 0.02 (−0.11, 0.15) | 0.02 (−0.10, 0.15) | |
| Psoas muscle attenuation | 1 | 0.01 (−0.14, 0.15) | 0.00 (−0.15, 0.14) | 0.00 (−0.14, 0.14) |
| 2 | 0.07 (−0.08, 0.21) | 0.03 (−0.11, 0.16) | 0.04 (−0.10, 0.17) | |
| Thigh muscle attenuation | 1 | −0.05 (−0.19, 0.09) | 0.00 (−0.15, 0.14) | −0.01 (−0.16, 0.13) |
| 2 | 0.02 (−0.12, 0.17) | 0.03 (−0.10, 0.17) | 0.03 (−0.10, 0.16) | |
| Liver attenuation | 1 | 0.12 (−0.02, 0.26) | 0.13 (−0.02, 0.27) | 0.13 (−0.01, 0.27) |
| 2 | 0.09 (−0.05, 0.24) | 0.10 (−0.04, 0.24) | 0.10 (−0.04, 0.25) |
| BMI or Fat Measures | Adjusted R2 | p-Value |
|---|---|---|
| BMI | 1.93% | 0.0064 |
| VAT | 1.85% | 0.0085 |
| Pericardial fat | 1.14% | 0.0519 |
| Paraspinous IMAT | 0.69% | 0.2030 |
| Psoas IMAT | 0.75% | 0.1649 |
| Thigh IMAT | 0.57% | 0.2968 |
| Paraspinous muscle attenuation | 0.33% | 0.7050 |
| Psoas muscle attenuation | 0.54% | 0.3388 |
| Thigh muscle attenuation | 0.31% | 0.7520 |
| Liver attenuation | 0.67% | 0.2146 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tilves, C.; Mueller, N.T.; Zmuda, J.M.; Kuipers, A.L.; Methé, B.; Li, K.; Carr, J.J.; Terry, J.G.; Wheeler, V.; Nair, S.; et al. Associations of Fecal Microbiota with Ectopic Fat in African Caribbean Men. Microorganisms 2024, 12, 812. https://doi.org/10.3390/microorganisms12040812
Tilves C, Mueller NT, Zmuda JM, Kuipers AL, Methé B, Li K, Carr JJ, Terry JG, Wheeler V, Nair S, et al. Associations of Fecal Microbiota with Ectopic Fat in African Caribbean Men. Microorganisms. 2024; 12(4):812. https://doi.org/10.3390/microorganisms12040812
Chicago/Turabian StyleTilves, Curtis, Noel T. Mueller, Joseph M. Zmuda, Allison L. Kuipers, Barbara Methé, Kelvin Li, John Jeffrey Carr, James G. Terry, Victor Wheeler, Sangeeta Nair, and et al. 2024. "Associations of Fecal Microbiota with Ectopic Fat in African Caribbean Men" Microorganisms 12, no. 4: 812. https://doi.org/10.3390/microorganisms12040812