
Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. DNA Preparation and Genome Sequencing, Assembly, and Annotations
2.3. Comparative Genome and COG Analyses
2.4. Antimicrobial Susceptibility and Biofilm Formation Assay
2.5. Infection Experiments for S. zooepidemicus Strain ZHZ 211
2.6. Statistical Analysis
3. Results
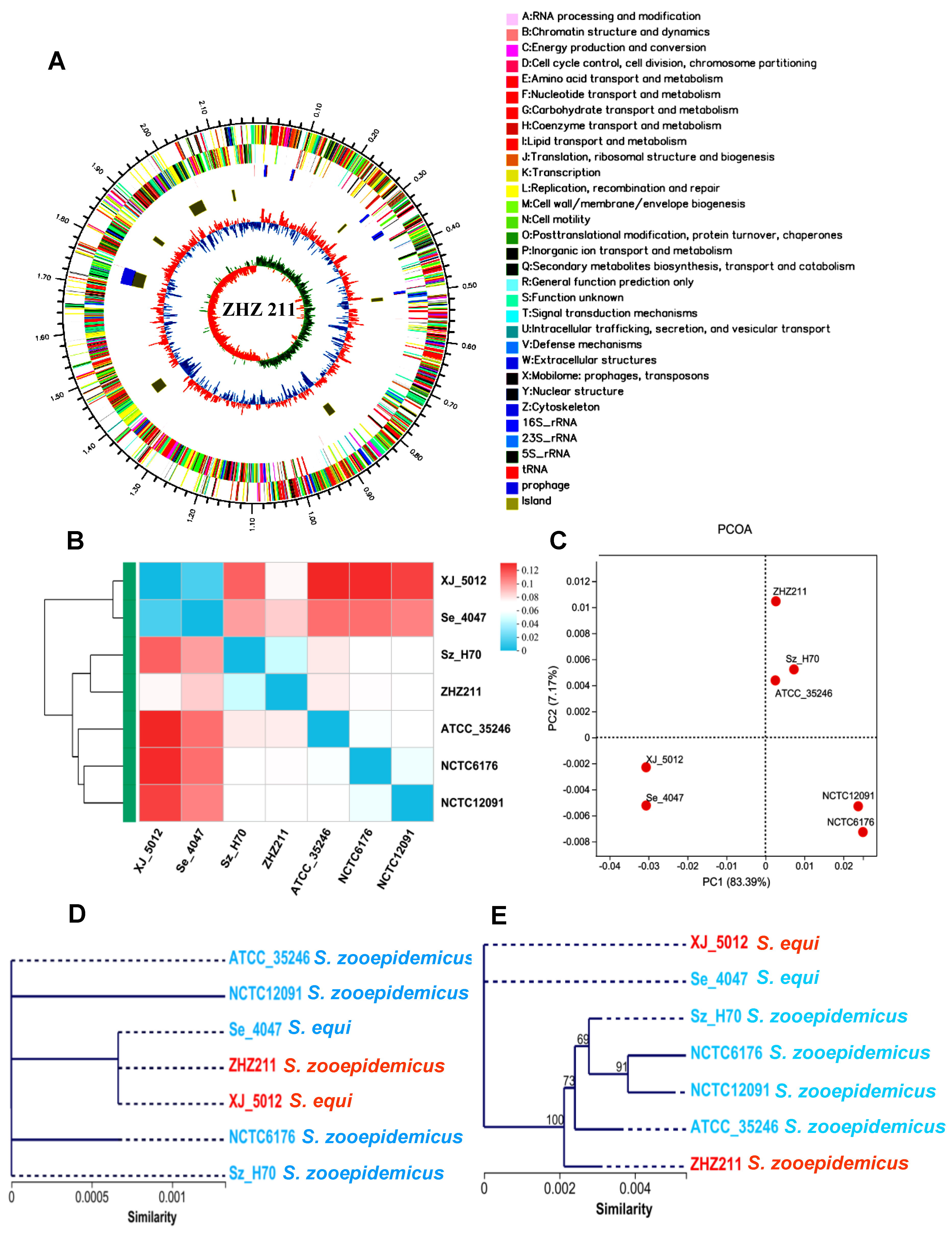
3.1. Genome Sequencing and Genomic Features of ZHZ 211 Strain
3.2. Genomic Synteny and Core Genome Analyses
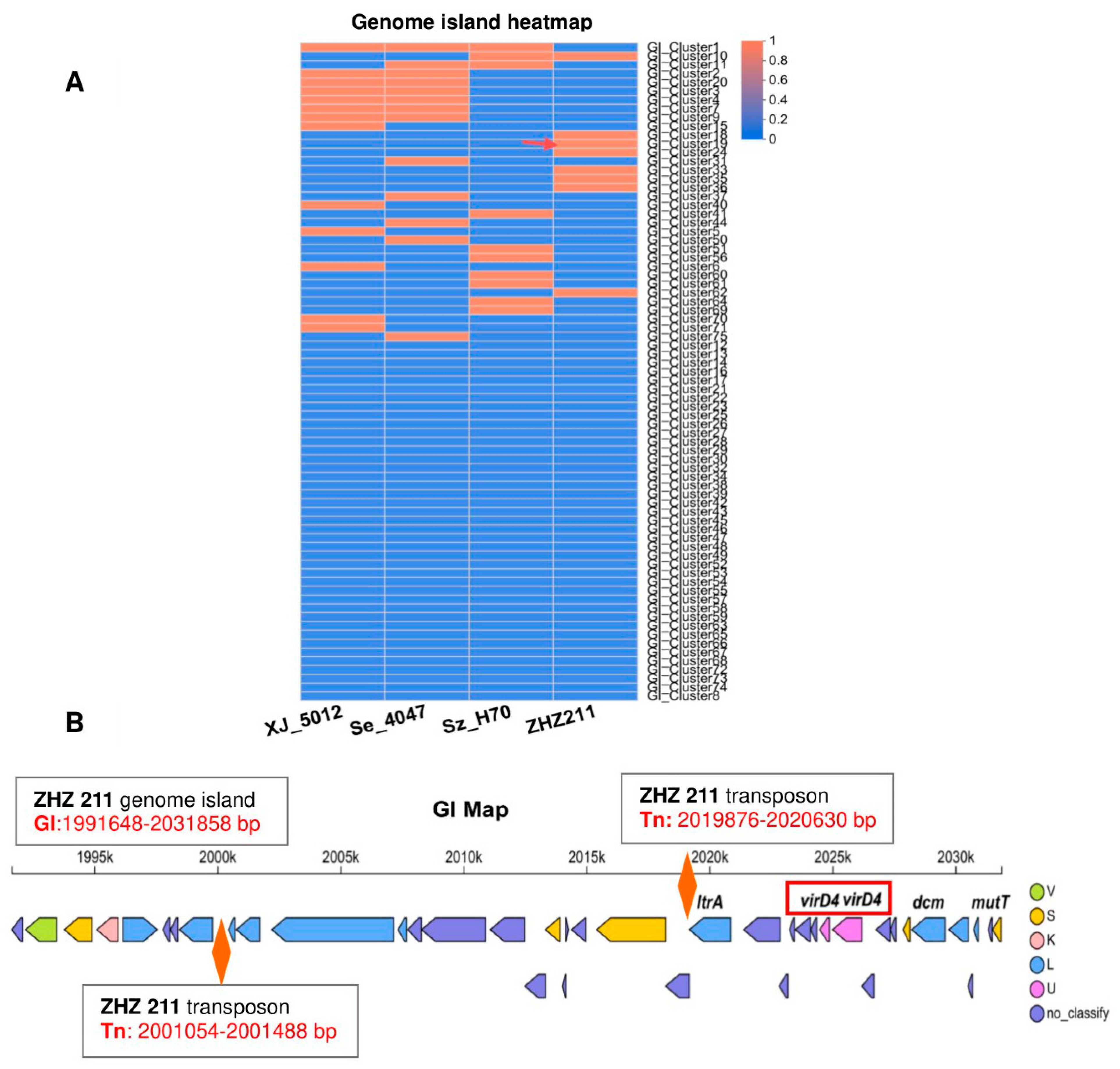
3.3. Comparative Genome Analysis and Prophages of ZHZ 211
3.4. Growth, Antimicrobial Resistance Gene, Phenotype, and Biofilm Formation of ZHZ 211
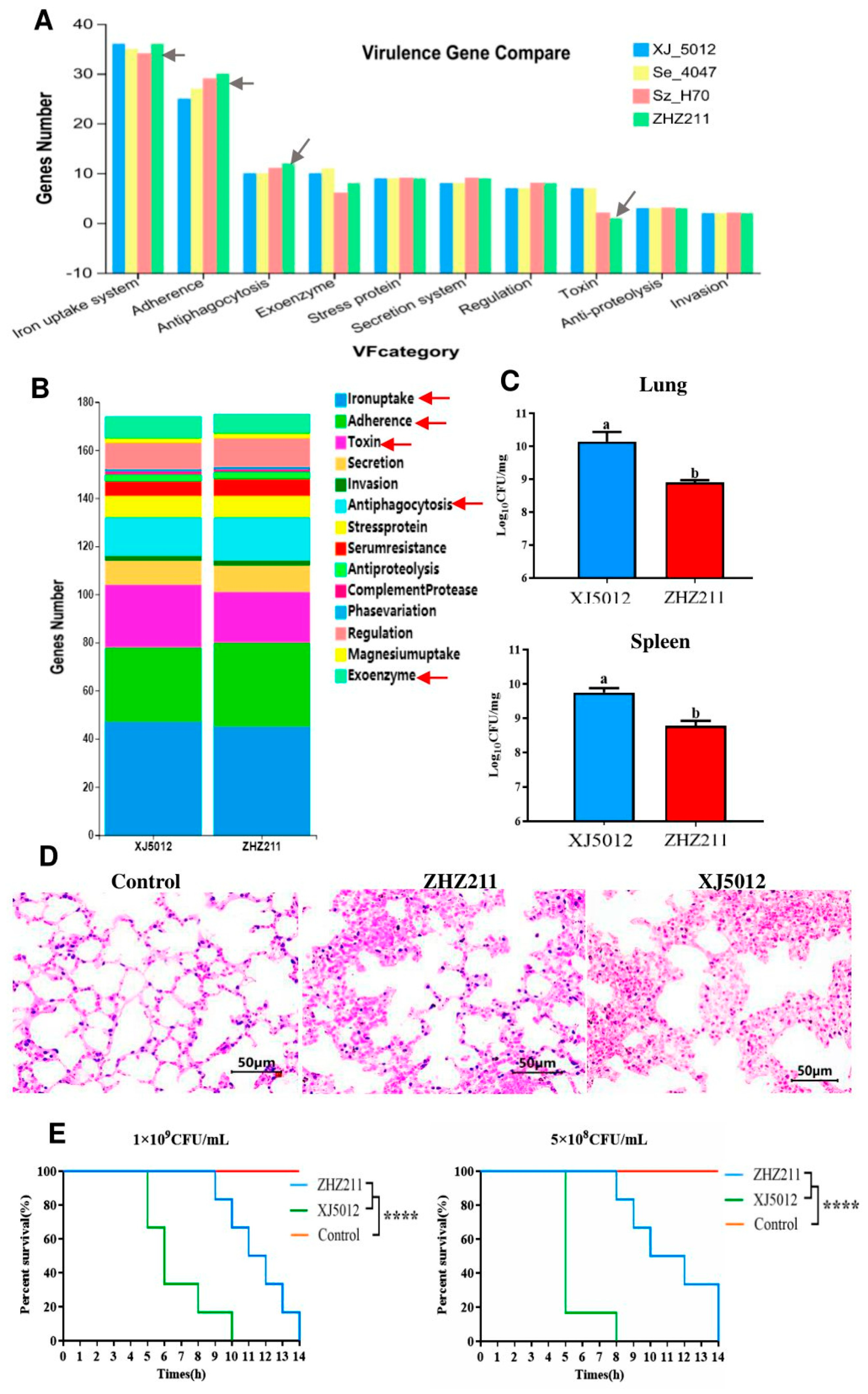
3.5. Virulence Genes and Virulence-Associated Phenotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Waller, A.S.; Robinson, C. Streptococcus zooepidemicus and Streptococcus equi evolution: The Role of CRISPRs. Biochem. Soc. Trans. 2013, 41, 1437–1443. [Google Scholar] [CrossRef] [PubMed]
- Bjornsdottir, S.; Harris, S.R.; Svansson, V.; Gunnarsson, E.; Sigurethardottir, O.G.; Gammeljord, K.; Steward, K.F.; Newton, J.R.; Robinson, C.; Charbonneau, A.R.L.; et al. Genomic dissection of an Icelandic epidemic of respiratory disease in horses and associated zoonotic cases. mBio 2017, 8, e00826-17. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, S.; Båverud, V.; Egenvall, A.; Aspán, A.; Pringle, J. Comparison of sampling sites and laboratory diagnostic tests for S. equi subsp. equi in horses from confirmed strangles outbreaks. J. Vet. Intern. Med. 2013, 27, 542–547. [Google Scholar] [CrossRef] [PubMed]
- Acke, E.; Midwinter, A.C.; Lawrence, K.; Gordon, S.J.; Moore, S.; Rasiah, I.; Steward, K.; French, N.; Waller, A. Prevalence of Streptococcus dysgalactiae subsp. equisimilis and S. equi subsp. zooepidemicus in a sample of healthy dogs, cats and horses. N. Z. Vet. J. 2015, 63, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, M.; Bojesen, A.M.; Petersen, M.R.; Christensen, H. A major outbreak of Streptococcus equi subsp. zooepidemicus infections in free-range chickens is linked to horses. Avian Dis. 2012, 56, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Geiping, L.; Detlefsen, H.; Trittmacher, S.; Baums, C.G.; Bergmann, R.; Hennig-Pauka, I. Streptococcus equi subspecies zooepidemicus—A case report of sudden death in a German sow farm. Porcine. Health. Manag. 2023, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Priestnall, S.; Erles, K. Streptococcus zooepidemicus: An emerging canine pathogen. Vet. J. 2011, 188, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Abbott, Y.; Acke, E.; Khan, S.; Muldoon, E.G.; Markey, B.K.; Pinilla, M.; Leonard, F.C.; Steward, K.; Waller, A. Zoonotic transmission of Streptococcus equi subsp. zooepidemicus from a dog to a handler. J. Med. Microbiol. 2010, 59, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.W.; Kenkre, J.S.; Bowler, I.C.; McBride, S.J. Streptococcus equi subspecies zooepidemicus meningitis: A case report and review of the literature. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 1459–1463. [Google Scholar] [CrossRef]
- Bosica, S.; Chiaverini, A.; Angelis, M.E.D.; Petrini, A.; Averaimo, D.; Martino, M.; Rulli, M.; Saletti, M.A.; Cantelmi, M.C.; Ruggeri, F.; et al. Early Release—Severe Streptococcus equi subspecies zooepidemicus outbreak from unpasteurized dairy product consumption, Italy. Emerg. Infect. Dis. J. 2023, 29, 5. [Google Scholar] [CrossRef]
- Hoyer-Nielsen, A.K.; Gaini, S.; Kjerulf, A.; Kollslieth, R.; Steig, T.A.; Stegger, M.; Jóanesarson, J. Sepsis, endocarditis, and purulent arthritis due to a rare zoonotic infection with Streptococcus equi subspecies zooepidemicus. Case Rep. Infect. Dis. 2018, 2018, 3265701. [Google Scholar] [CrossRef]
- Held, J.; Schmitz, R.; van der Linden, M.; Nuhrenberg, T.; Hacker, G.; Neumann, F.J. Purulent pericarditis and pneumoniacaused by Streptococcus equi subspecies zooepidemicus. J. Med. Microbiol. 2014, 63 Pt 2, 313–316. [Google Scholar] [CrossRef]
- Holden, M.T.; Heather, Z.; Paillot, R.; Steward, K.F.; Webb, K.; Ainslie, F.; Jourdan, T.; Bason, N.C.; Holroyd, N.E.; Mungall, K.; et al. Genomic evidence for the evolution of Streptococcus equi: Host restriction, increased virulence, and genetic exchange with human pathogens. PLoS Pathog. 2009, 5, e1000346. [Google Scholar] [CrossRef]
- Ma, Z.; Geng, J.; Zhang, H.; Yu, H.; Yi, L.; Lei, M.; Lu, C.P.; Fan, H.J.; Hu, S. Complete genome sequence of Streptococcus equi subsp. zooepidemicus strain ATCC 35246. J. Bacteriol. 2011, 193, 5583–5584. [Google Scholar] [CrossRef]
- Beres, S.B.; Sesso, R.; Pinto, S.W.; Hoe, N.P.; Porcella, S.F.; Deleo, F.R.; Musser, J.M. Genome sequence of a Lancefield group C Streptococcus zooepidemicus strain causing epidemic nephritis: New information about an old disease. PLoS ONE 2008, 3, e3026. [Google Scholar] [CrossRef]
- Paillot, R.; Darby, A.C.; Robinson, C.; Wright, N.L.; Steward, K.F.; Anderson, E.; Webb, K.; Holden, M.T.; Efstratiou, A.; Broughton, K.; et al. Identification of three novel superantigen-encoding genes in Streptococcus equi subsp. zooepidemicus, szeF, szeN, and szeP. Infect. Immun. 2010, 78, 4817–4827. [Google Scholar] [CrossRef]
- Lefebure, T.; Richards, V.P.; Lang, P.; Pavinski-Bitar, P.; Stanhope, M.J. Gene repertoire evolution of Streptococcus pyogenes inferred from phylogenomic analysis with Streptococcus canis and Streptococcus dysgalactiae. PLoS ONE 2012, 7, e37607. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alber, J.; El-Sayed, A.; Estoepangestie, S.; Lämmler, C.; Zschöck, M. Dissemination of the superantigen encoding genes seeL, seeM, szeL and szeM in Streptococcus equi subsp. equi and Streptococcus equi subsp. zooepidemicus. Vet. Microbiol. 2005, 109, 135–141. [Google Scholar] [CrossRef]
- Liu, W.; Fang, L.; Li, M.; Li, S.; Guo, S.; Luo, R.; Feng, Z.; Li, B.; Zhou, Z.; Shao, G.; et al. Comparative genomics of Mycoplasma: Analysis of conserved essential genes and diversity of the pan-genome. PLoS ONE 2012, 7, e35698. [Google Scholar] [CrossRef]
- Liu, W.; Xiao, S.; Li, M.; Guo, S.; Li, S.; Luo, R.; Feng, Z.; Li, B.; Zhou, Z.; Shao, G.; et al. Comparative genomic analyses of Mycoplasma hyopneumoniae pathogenic 168 strain and its high-passaged attenuated strain. BMC Genom. 2013, 14, 80. [Google Scholar] [CrossRef]
- Szczepanek, S.M.; Tulman, E.R.; Gorton, T.S.; Liao, X.; Lu, Z.; Zinski, J.; Aziz, F.; Frasca, S., Jr.; Kutish, G.F.; Geary, S.J. Comparative genomic analyses of attenuated strains of Mycoplasma gallisepticum. Infect. Immun. 2010, 78, 1760–1771. [Google Scholar] [CrossRef]
- Bertelli, C.; Greub, G. Rapid bacterial genome sequencing: Methods and applications in clinical microbiology. Clin. Microbiol. Infect. 2013, 19, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Fulde, M.; Valentin-Weigand, P. Epidemiology and pathogenicity of zoonotic streptococci. Curr. Top. Microbiol. Immunol. 2013, 368, 49–81. [Google Scholar] [CrossRef]
- Laus, F.; Preziuso, S.; Spaterna, A.; Beribe, F.; Tesei, B.; Cuteri, V. Clinical and epidemiological investigation of chronic upper respiratory diseases caused by beta-haemolytic Streptococci in horses. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Newton, J.R.; Laxton, R.; Wood, J.L.; Chanter, N. Molecular epidemiology of Streptococcus zooepidemicus infection in naturally occurring equine respiratory disease. Vet. J. 2008, 175, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.; Steward, K.F.; Charbonneau, A.R.L.; Walsh, S.; Wilson, H.; Timoney, J.F.; Wernery, U.; Joseph, M.; Craig, D.; van Maanen, K.; et al. Globetrotting strangles: The unbridled national and international transmission of Streptococcus equi between horses. Microb. Genom. 2021, 7, 000528. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Tang, H. ISEScan: Automated identification of insertion sequence elements in prokaryotic genomes. Bioinformatics 2017, 33, 3340–3347. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucl. Acids Res. 2016, 44, 16–21. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucl. Acids Res. 2017, 45, 30–35. [Google Scholar] [CrossRef]
- Bland, C.; Ramsey, T.L.; Sabree, F.; Lowe, M.; Brown, K.; Kyrpides, N.C.; Hugenholtz, P. CRISPR recognition tool (CRT): A tool for automatic detection of clustered regularly interspaced palindromic repeats. BMC Bioinform. 2007, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic islands: Tools of bacterial horizontal gene transfer and evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.; Petty, N.K.; Zakou, N.L.B.; Beatson, S.A.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.E.; Mau, B.; Perna, N.T. ProgressiveMauve: Multiple genome alignment with gene gain, loss and rearrangement. PLoS ONE 2010, 5, e11147. [Google Scholar] [CrossRef] [PubMed]
- Lam, F.; Lalansingh, C.M.; Babaran, H.E.; Wang, Z.; Prokopec, S.D.; Fox, N.S.; Boutros, P.C. VennDiagramWeb: A web application for the generation of highly customizable Venn and Euler diagrams. BMC Bioinform. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- M100-S23; Performance Standards for Antimicrobial Susceptibility Testing. 23rd informational supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2013.
- Haney, E.F.; Trimble, M.J.; Hancock, R.E.W. Microtiter plate assays to assess antibiofilm activity against bacteria. Nat. Protoc. 2021, 16, 2615–2632. [Google Scholar] [CrossRef] [PubMed]
- Timoney, J.F. The pathogenic equine streptococci. Vet. Res. 2004, 35, 397–409. [Google Scholar] [CrossRef]
- Costa, M.O.; Lage, B. Streptococcus equi subspecies zooepidemicus and sudden deaths in swine, Canada. Emerg. Infect. Dis. 2020, 26, 2522–2524. [Google Scholar] [CrossRef]
- Bower, C.; Daeschel, M.A. Resistance responses of microorganisms in food environments. Int. J. Food Microbiol. 1999, 50, 33–44. [Google Scholar] [CrossRef]
- Stepanovic, S.; Cirkovic, I.; Ranin, L.; Svabic-Vlahovic, M. Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 2004, 38, 428–432. [Google Scholar] [CrossRef]
- Hanning, I.; Donoghue, D.J.; Jarquin, R.; Kumar, G.S.; Aguiar, V.F.; Metcalf, J.H.; Reyes-Herrera, I.; Slavik, M. Campylobacter biofilm phenotype exhibits reduced colonization potential in young chickens and altered in vitro virulence. Poult. Sci. 2009, 88, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Holmberg, A.; Lood, R.; Morgelin, M.; Holst, E.; Collin, M.; Christensson, B.; Rasmussen, M. Biofilm formation by Propioni bacterium acnes is a characteristic of invasive isolates. Clin. Microbiol. Infect. 2009, 15, 787–795. [Google Scholar] [CrossRef]
- Jain, A.; Agarwal, A. Biofilm production, a marker of pathogenic potential of colonizing and commensal staphylococci. J. Micro Biol. Meth. 2009, 76, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.; Feng, Y.; Wang, C.; Zheng, F.; Li, M.; Liao, H.; Mao, Y.; Pan, X.; Wang, J.; Hu, D.; et al. Functional definition of LuxS, an autoinducer-2 (AI-2) synthase and its role in full virulence of Streptococcus suis serotype 2. J. Microbiol. 2011, 49, 1000–1011. [Google Scholar] [CrossRef]
- Grenier, D.; Grignon, L.; Gottschalk, M. Characterisation of biofilm formation by a Streptococcus suis meningitis isolate. Vet. J. 2009, 179, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Furukawa, T.; Matsumoto-Mashimo, C.; Yamane, K.; Sugimori, C.; Nambu, T.; Mori, N.; Nishikawa, H.; Walker, C.B.; Leung, K.P.; et al. Gene expression profile and pathogenicity of biofilm-forming prevotella intermedia strain 17. BMC Microbiol. 2009, 9, 11. [Google Scholar] [CrossRef]
- Karslake, J.; Maltas, J.; Brumm, P.; Wood, K.B. Population density modulates drug inhibition and gives rise to potential bista bility of treatment outcomes for bacterial infections. PLoS Comput. Biol. 2016, 12, e1005098. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Ahmed, S.; Wang, F.; Gu, Y.; Zhang, C. Antimicrobial activity and resistance: Influencing factors. Front. Pharmacol. 2017, 8, 364. [Google Scholar] [CrossRef]
- Vulin, C.; Leimer, N.; Huemer, M.; Ackermann, M.; Zinkernagel, A.S. Prolonged bacterial lag time results in small colony varants that represent a sub-population of persisters. Nat. Commun. 2018, 9, 4074. [Google Scholar] [CrossRef]
- Novais, J.S.; Carvalho, M.F.; Ramundo, M.S.; Beltrame, C.O.; Geraldo, R.B.; Jordão, A.K.; Ferreira, V.F.; Castro, H.C.; Figueiredo, A.M.S. Antibiofilm effects of N,O-acetals derived from 2-amino-1,4-naphthoquinone are associated with downregulation of important global virulence regulators in methicillin-resistant Staphylococcus aureus. Sci. Rep. 2020, 10, 19631. [Google Scholar] [CrossRef]
- Thulin, P.; Johansson, L.; Low, D.E.; Gan, B.S.; Kotb, M.; McGeer, A.; Norrby-Teglund, A. Viable group A Streptococci in macro phages during acute soft tissue infection. PLoS Med. 2006, 3, e53. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.R.; Dong, S.; Pritchard, D.G. The hyaluronan lyase of Streptococcus pyogenes bacteriophage H4489A. Biochem. J. 2002, 365 Pt 1, 317–322. [Google Scholar] [CrossRef]
- Brussow, H.; Canchaya, C.; Hardt, W.D. Phages and the evolution of bacterial pathogens: From genomic rearrangements to lysogenic conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Domingues, R.; Evans, B.; Sutton, J.M.; Adriaenssens, E.M.; Turner, D. Genomic diversity of bacteriophages infect ing the genus acinetobacter. Viruses 2022, 14, 181. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Park, J.H. Characterization of prophages in Leuconostoc derived from Kimchi and genomic analysis of the induced prophage in Leuconostoc lactis. J. Microbiol. Biotechnol. 2022, 32, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Nepal, R.; Houtak, G.; Shaghayegh, G.; Bouras, G.; Shearwin, K.; Psaltis, A.J. Prophages encoding human immune evasion clus ter genes are enriched in Staphylococcus aureus isolated from chronic rhinosinusitis patients with nasal polyps. Microb. Genom. 2021, 7, 000726. [Google Scholar] [CrossRef] [PubMed]
- Artiushin, S.C.; Timoney, J.F.; Sheoran, A.S.; Muthupalani, S.K. Character-ization and immunogenicity of pyrogenic mitogens SePE-H and SePE-I of Streptococcus equi. Microb. Pathog. 2022, 32, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Igwe, E.I.; Shewmaker, P.L.; Facklam, R.R.; Farley, M.M.; van Beneden, C.; Beall, B. Identification of superantigen genes speM, ssa, and smeZ in invasive strains of beta-hemolytic group C and G streptococci recovered from humans. FEMS Microbiol. Lett. 2003, 229, 259–264. [Google Scholar] [CrossRef]
- Proft, T.; Webb, P.D.; Handley, V.; Fraser, J.D. Two novel superantigens found in both group A and group C Streptococcus. Infect. Immun. 2003, 71, 1361–1369. [Google Scholar] [CrossRef]
- Sheng, J.; Ling, P.; Wang, F. Constructing a recombinant hyaluronic acid biosynthesis operon and producing food-grade hyalu ronic acid in Lactococcus lactis. J. Ind. Microbiol. Biotechnol. 2015, 42, 197–206. [Google Scholar] [CrossRef]
- Wei, Z.; Fu, Q.; Chen, Y.; Cong, P.; Xiao, S.; Mo, D.; He, Z.; Liu, X. The capsule of Streptococcus equi ssp. zooepidemicus is a target for attenuation in vaccine development. Vaccine 2012, 30, 4670–4675. [Google Scholar] [CrossRef] [PubMed]
- Hynes, W.L.; Walton, S.L. Hyaluronidases of gram-positive bacteria. FEMS Microbiol. Lett. 2000, 183, 201–207. [Google Scholar] [CrossRef]
- Moncalián, G.; Cabezón, E.; Alkorta, I.; Valle, M.; Moro, F.; Valpuesta, J.M.; Goñi, F.M.; de La Cruz, F. Characterization of ATP and DNA binding activities of TrwB, the coupling protein essential in plasmid R388 conjugation. J. Biol. Chem. 1999, 274, 36117–36124. [Google Scholar] [CrossRef][Green Version]
- Beranek, A.; Zettl, M.; Lorenzoni, K.; Schauer, A.; Manhart, M.; Koraimann, G.J. Thirty-eight C-terminal amino acids of the coupling protein TraD of the F-like conjugative resistance plasmid R1 are required and sufficient to confer binding to the substrate selector protein TraM. J. Bacteriol. 2004, 186, 6999–7006. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schröder, G.; Krause, S.; Zechner, E.L.; Traxler, B.; Yeo, H.J.; Lurz, R.; Waksman, G.; Lanka, E. TraG-like proteins of DNA transfer systems and of the Helicobacter pylori type IV secretion system: Inner membrane gate for exported substrates? J. Bacteriol. 2002, 184, 2767–2779. [Google Scholar] [CrossRef] [PubMed]
- Grohmann, E.; Christie, P.J.; Waksman, G.; Backert, S. Type IV secretion in gram-negative and gram-positive bacteria. Mol. Microbiol. 2018, 107, 455–471. [Google Scholar] [CrossRef]
- Costa, T.R.D.; Harb, L.; Khara, P.; Zeng, L.; Hu, B.; Christie, P.J. Type IV secretion systems: Advances in structure, function, and activation. Mol. Microbiol. 2021, 115, 436–452. [Google Scholar] [CrossRef]
- Selbach, M. Functional analysis of the Helicobacter pylori cag pathogenicity island reveals both virD4-cagA-dependent and virD4-cagA-independent mechanisms. Infect. Immun. 2002, 70, 665–671. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, Y.; Zhou, J.; Zhu, L.; Gu, Y.; Zhang, X.; Li, X.; Fang, W. Roles of the putative type IV-like secretion system key component VirD4 and PrsA in pathogenesis of Streptococcus suis type 2. Front. Cell Infect. Microbiol. 2016, 6, 00172. [Google Scholar] [CrossRef]
- Zhang, W.; Rong, C.; Chen, C.; Gao, G.F. Type-IVC secretion system: A novel subclass of type IV secretion system (T4SS) common existing in gram-positive genus Streptococcus. PLoS ONE 2012, 7, e46390. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | ZHZ 211 | Genes | XJ 5012 | Genes |
|---|---|---|---|---|
| Amoxicilin | S | PBP2x, PBP1a, soxS, PBP2b | S | PBP2x, PBP1a, soxS, PBP2b |
| Ampicillin | R | PBP2x, PBP1a, soxS, PBP2b | S | PBP2x, PBP1a, soxS, PBP2b |
| Cefuroxime | R | PBP2x, PBP1a, soxS, PBP2b | R | PBP2x, PBP1a, soxS, PBP2b |
| Ceftiofur | S | PBP2x, PBP1a, soxS, PBP2b | S | PBP2x, PBP1a, soxS, PBP2b S |
| Cefoxitin | S | PBP2x, PBP1a, soxS, PBP2b | S | PBP2x, PBP1a, soxS, PBP2b |
| Penicillin | R | PBP2x, PBP1a, soxS, PBP2b | S | PBP2x, PBP1a, soxS, PBP2b |
| Gentamicin | S | vgaE, lmrP, rpsL, cfrA, baeS, lsaC, salA, baeS, gidB | R | vgaE, lmrP, gidB, rpsL |
| Streptomycin | S | vgaE, lmrP, rpsL, cfrA, baeS, lsaC, salA, baeS, gidB | S | vgaElmrP, gidB, rpsL |
| Erythromycin | S | macBlmrP, cfrA, mefE, efrA, efrB, mtrA, | S | macBlmrP, efrA, mtrA, mefE, efrB |
| Clarithromycin | R | macBlmrP, cfrA, mefE, efrA, efrB, mtrA | R | macBlmrP, efrA, mtrA, mefE efrB, |
| Doxycycline | S | macBlmrP, cfrA, mefE, efrA, efrB, mtrA | R | macBlmrP, efrA, mtrA, mefE efrB, |
| Oxytetracycline | S | lmrP, tetB(P), adeR, soxS, rpsJ | S | lmrP, soxS, adeR, tetB(P), tetT, rpsJ |
| Tetracycline | S | lmrP, tetB(P), adeR, soxS, rpsJ | R | lmrP, soxS, adeR, tetB(P), tetT, rpsJ |
| Levofloxacin | S | patB, arlR, arlS, gyrA, efrA, efrB, soxS | S | efrA, arlR, arlS, soxS, efrB, gyrA, patB |
| Norfloxacin | S | patB, arlR, arlS, gyrA, efrA, efrB, soxS | S | efrA, arlR, arlS, soxS, efrB, gyrA, patB |
| Enrofloxacin | R | patB, arlR, arlS, gyrA, efrA, efrB, soxS | R | efrA, arlR, arlS, soxS, efrB, gyrA, patB, |
| Ciprofloxacin | S | patB, arlR, arlS, gyrA, efrA, efrB, soxS | S | efrA, arlR, arlS, soxS, efrB, gyrA, patB |
| Sulfafurazole | R | / | S | / |
| Sulfadiazine Sodium | R | / | S | / |
| Rifampin | S | efrA, efrB, soxS, rpoB | S | efrA, soxS, efrB, rpoB |
| Clindymycin | S | lmrP, cfrA, lsaC, salA, lmrB | S | lmrP, lmrB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Zhang, Z.; Wang, L.; Zhang, B.; Su, L. Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China. Microorganisms 2024, 12, 824. https://doi.org/10.3390/microorganisms12040824
Su Y, Zhang Z, Wang L, Zhang B, Su L. Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China. Microorganisms. 2024; 12(4):824. https://doi.org/10.3390/microorganisms12040824
Chicago/Turabian StyleSu, Yan, Zehua Zhang, Li Wang, Baojiang Zhang, and Lingling Su. 2024. "Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China" Microorganisms 12, no. 4: 824. https://doi.org/10.3390/microorganisms12040824
APA StyleSu, Y., Zhang, Z., Wang, L., Zhang, B., & Su, L. (2024). Whole-Genome Sequencing and Phenotypic Analysis of Streptococcus equi subsp. zooepidemicus Sequence Type 147 Isolated from China. Microorganisms, 12(4), 824. https://doi.org/10.3390/microorganisms12040824