
The Identification of the Mitochondrial DNA Polymerase γ (Mip1) of the Entomopathogenic Fungus Metarhizium brunneum




Abstract
1. Introduction
2. Materials and Methods
2.1. Media, Strains and Growth Conditions
2.2. Nucleic Acid Manipulations
2.3. Transformation
2.4. Microscopy
2.5. Protein Molecular Modeling
2.6. RNA Extraction, First-Strand cDNA Synthesis and Quantitative Real-Time PCR
2.7. Statistical Analysis
3. Results
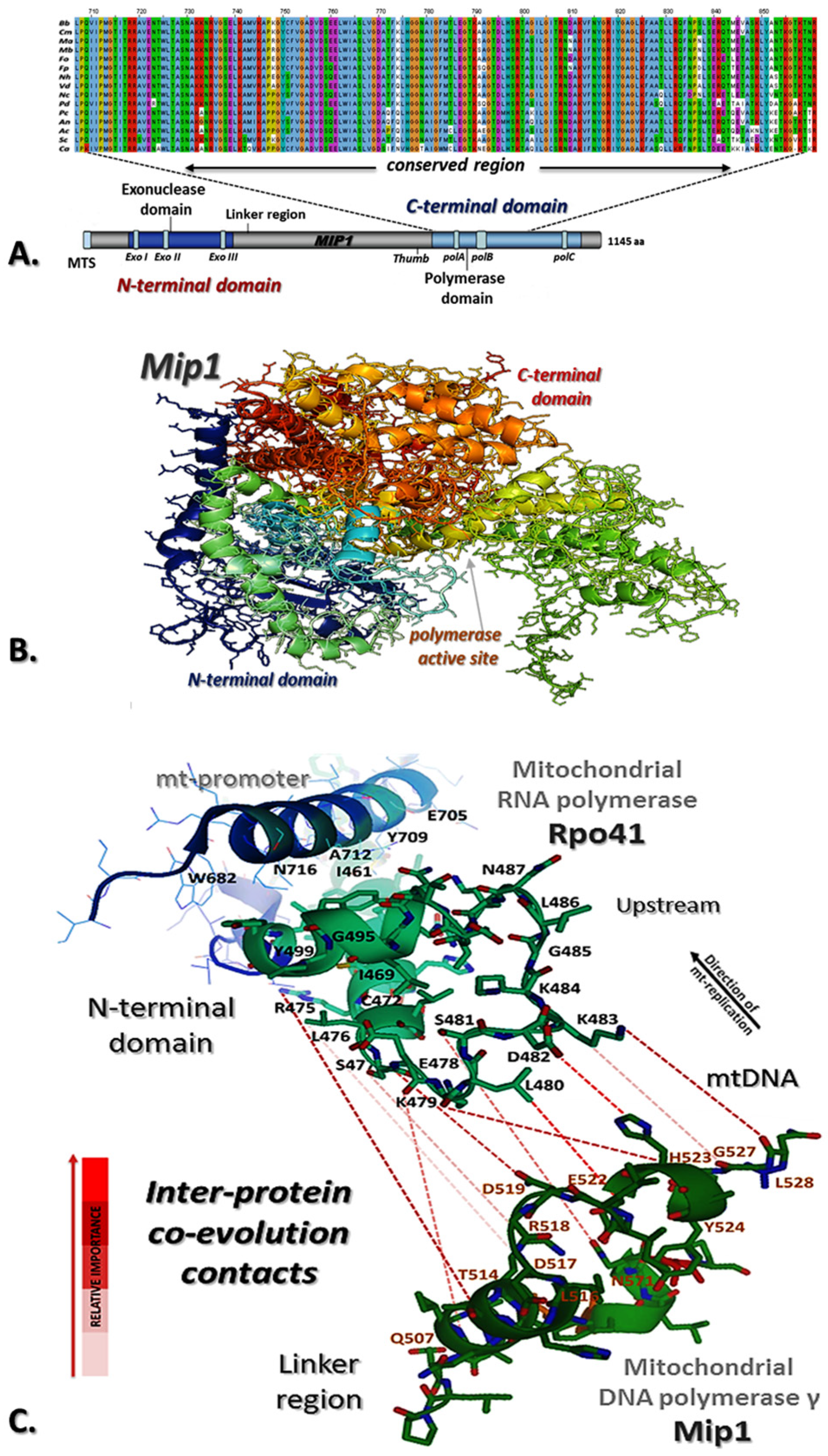
3.1. Identification of M. brunneum Mip1
3.2. Mip1 Has an Important Role in Fungal Cell Growth, Morphogenesis, Conidiation and Stress Tolerance
3.3. In Silico Analysis of M. brunneum Mip1 and Interactions with Rpo41-Mtf1 Complex
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foury, F. Cloning and sequencing of the nuclear gene MIP1 encoding the catalytic subunit of the yeast mitochondrial DNA polymerase. J. Biol. Chem. 1989, 264, 20552–20560. [Google Scholar] [CrossRef]
- Viikov, K.; Väljamäe, P.; Sedman, J. Yeast mitochondrial DNA polymerase is a highly processive single-subunit enzyme. Mitochondrion 2011, 11, 119–126. [Google Scholar] [CrossRef]
- Lucas, P.; Lasserre, J.P.; Plissonneau, J.; Castroviejo, M. Absence of accessory subunit in the DNA polymerase γ purified from yeast mitochondria. Mitochondrion 2004, 4, 13–20. [Google Scholar] [CrossRef]
- Greenleaf, A.L.; Kelly, J.L.; Lehman, I.R. Yeast RPO41 gene product is required for transcription and maintenance of the mitochondrial genome. Proc. Natl. Acad. Sci. USA 1986, 83, 3391–3394. [Google Scholar] [CrossRef]
- Schinkel, A.H.; Koerkamp, M.G.; Tabak, H.F. Mitochondrial RNA polymerase of Saccharomyces cerevisiae: Composition and mechanism of promoter recognition. EMBO J. 1988, 7, 3255. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Sun, W.; Wang, Z.; Zhang, J.; Chen, D.; Murchie, A.I. Identification and characterization of the mitochondrial RNA polymerase and transcription factor in the fission yeast Schizosaccharomyces pombe. Nucleic Acids Res. 2011, 39, 5119–5130. [Google Scholar] [CrossRef]
- Lisowsky, T.; Wilkens, D.; Stein, T.; Hedtke, B.; Börner, T.; Weihe, A. The C-terminal region of mitochondrial single-subunit RNA polymerases contains species-specific determinants for maintenance of intact mitochondrial genomes. Mol. Biol. Cell 2002, 13, 2245–2255. [Google Scholar] [CrossRef]
- Cermakian, N.; Ikeda, T.M.; Miramontes, P.; Lang, B.F.; Gray, M.W.; Cedergren, R. On the evolution of the single-subunit RNA polymerases. J. Mol. Evol. 1997, 45, 671–681. [Google Scholar] [CrossRef]
- Filée, J.; Forterre, P.; Sen-Lin, T.; Laurent, J. Evolution of DNA polymerase families: Evidences for multiple gene exchange between cellular and viral proteins. J. Mol. Evol. 2002, 54, 763–773. [Google Scholar] [CrossRef]
- Varassas, S.P.; Kouvelis, V.N. Mitochondrial Transcription of Entomopathogenic Fungi Reveals Evolutionary Aspects of Mitogenomes. Front. Microbiol. 2022, 13, 821638. [Google Scholar] [CrossRef]
- Trasviña-Arenas, C.H.; Hoyos-Gonzalez, N.; Castro-Lara, A.Y.; Rodriguez-Hernandez, A.; Sanchez-Sandoval, M.E.; Jimenez-Sandoval, P.; Ayala-García, V.M.; Díaz-Quezada, C.; Lodi, T.; Baruffini, E.; et al. Amino and carboxy-terminal extensions of yeast mitochondrial DNA polymerase assemble both the polymerization and exonuclease active sites. Mitochondrion 2019, 49, 166–177. [Google Scholar] [CrossRef]
- Young, M.J.; Imperial, R.J.; Lakhi, S.; Court, D.A. A non-radioactive DNA synthesis assay demonstrates that elements of the Sigma 1278b Mip1 mitochondrial DNA polymerase domain and C-terminal extension facilitate robust enzyme activity. Yeast 2021, 38, 262–275. [Google Scholar] [CrossRef]
- Freel, K.C.; Friedrich, A.; Schacherer, J. Mitochondrial genome evolution in yeasts: An all-encompassing view. FEMS Yeast Res. 2015, 15, fov023. [Google Scholar] [CrossRef]
- Kolondra, A.; Labedzka-Dmoch, K.; Wenda, J.M.; Drzewicka, K.; Golik, P. The transcriptome of Candida albicans mitochondria and the evolution of organellar transcription units in yeasts. BMC Genom. 2015, 16, 827. [Google Scholar] [CrossRef]
- Christinaki, A.C.; Kanellopoulos, S.G.; Kortsinoglou, A.M.; Andrikopoulos, M.A.; Theelen, B.; Boekhout, T.; Kouvelis, V.N. Mitogenomics and mitochondrial gene phylogeny decipher the evolution of Saccharomycotina yeasts. Genome Biol. Evol. 2022, 14, evac073. [Google Scholar] [CrossRef]
- Kortsinoglou, A.M.; Saud, Z.; Eastwood, D.C.; Butt, T.M.; Kouvelis, V.N. The mitochondrial genome contribution to the phylogeny and identification of Metarhizium species and strains. Fungal Biol. 2020, 124, 845–853. [Google Scholar] [CrossRef]
- Saud, Z.; Kortsinoglou, A.M.; Kouvelis, V.N.; Butt, T.M. Telomere length de novo assembly of all 7 chromosomes and mitogenome sequencing of the model entomopathogenic fungus, Metarhizium brunneum, by means of a novel assembly pipeline. BMC Genomics 2021, 22, 87. [Google Scholar] [CrossRef]
- Butt, T.M.; Jackson, C.; Magan, N. Fungi as Biocontrol Agents: Progress Problems and Potential; CABI: Egham, UK, 2001. [Google Scholar] [CrossRef]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control 2008, 46, 72–82. [Google Scholar] [CrossRef]
- Kimberly, M.S.A.; Seow, M.H. Mode of infection of Metarhizium spp. fungus and their potential as biological control agent’s fungi. J. Fungi 2017, 3, 30. [Google Scholar] [CrossRef]
- Apostolaki, A.; Harispe, L.; Calcagno-Pizarelli, A.M.; Vangelatos, I.; Sophianopoulou, V.; Arst, H.N., Jr.; Penalva, M.A.; Amillis, S.; Scazzocchio, C. Aspergillus nidulans CkiA is an essential casein kinase I required for delivery of amino acid transporters to the plasma membrane. Mol. Microbiol. 2012, 84, 530–549. [Google Scholar] [CrossRef]
- Goldstein, A.L.; McCusker, J.H. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 1999, 15, 1541–1553. [Google Scholar] [CrossRef]
- Koukaki, M.; Giannoutsou, E.; Karagouni, A.; Diallinas, G. A novel improved method for Aspergillus nidulans transformation. J. Microbiol. Methods 2003, 55, 687–695. [Google Scholar] [CrossRef]
- Aqeel, A.M.; Pasche, J.S.; Gudmestad, N.C. Variability in morphology and aggressiveness among North American vegetative compatibility groups of Colletotrichum coccodes. Phytopathology 2008, 98, 901–909. [Google Scholar] [CrossRef]
- Buchan, D.W.; Jones, D.T. The PSIPRED protein analysis workbench: 20 years on. Nucleic Acids Res. 2019, 47, W402–W407. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A completely reimplemented MPI bioinformatics toolkit with a new HHpred server at its core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Webb, B.; Sali, A. Comparative protein structure modeling using MODELLER. Curr. Protoc. Bioinform. 2016, 54, 5–6. [Google Scholar] [CrossRef]
- Doublie, S.; Tabor, S.; Long, A.M.; Richardson, C.C.; Ellenberger, T. Crystal structure of a bacteriophage T7 DNA replication complex at 2.2 Å resolution. Nature 1998, 391, 251–258. [Google Scholar] [CrossRef]
- Zhang, W.; Bell, E.W.; Yin, M.; Zhang, Y. EDock: Blind protein–ligand docking by replica-exchange monte carlo simulation. J. Cheminform. 2020, 12, 37. [Google Scholar] [CrossRef]
- De Wijngaert, B.; Sultana, S.; Singh, A.; Dharia, C.; Vanbuel, H.; Shen, J.; Vasilchuk, D.; Martinez, S.E.; Kandiah, E.; Patel, S.S.; et al. Cryo-EM structures reveal transcription initiation steps by yeast mitochondrial RNA polymerase. Mol. Cell 2021, 81, 268–280. [Google Scholar] [CrossRef]
- Felenbok, B. The ethanol utilization regulon of Aspergillus nidulans: The alcA-alcR system as a tool for the expression of recombinant proteins. J. Biotechnol. 1991, 17, 11–17. [Google Scholar] [CrossRef]
- Lee, S.; Lee, Y.J.; Choi, S.; Park, S.B.; Tran, Q.G.; Heo, J.; Kim, H.S. Development of an alcohol-inducible gene expression system for recombinant protein expression in Chlamydomonas reinhardtii. J. Appl. Phycol. 2018, 30, 2297–2304. [Google Scholar] [CrossRef]
- Yang, X.; Chang, H.R.; Yin, Y.W. Yeast mitochondrial transcription factor Mtf1 determines the precision of promoter-directed initiation of RNA polymerase Rpo41. PLoS ONE 2015, 10, e0136879. [Google Scholar] [CrossRef]
- Basu, U.; Bostwick, A.M.; Das, K.; Dittenhafer-Reed, K.E.; Patel, S.S. Structure, mechanism, and regulation of mitochondrial DNA transcription initiation. J. Biol. Chem. 2020, 295, 18406–18425. [Google Scholar] [CrossRef]
- Goovaerts, Q.; Shen, J.; De Wijngaert, B.; Basu, U.; Patel, S.S.; Das, K. Structures illustrate step-by-step mitochondrial transcription initiation. Nature 2023, 622, 872–879. [Google Scholar] [CrossRef]
- Sanchez-Sandoval, E.; Diaz-Quezada, C.; Velazquez, G.; Arroyo-Navarro, L.F.; Almanza-Martinez, N.; Trasviña-Arenas, C.H.; Brieba, L.G. Yeast mitochondrial RNA polymerase primes mitochondrial DNA polymerase at origins of replication and promoter sequences. Mitochondrion 2015, 24, 22–31. [Google Scholar] [CrossRef]
- Stuart, G.R.; Santos, J.H.; Strand, M.K.; Van Houten, B.; Copeland, W.C. Mitochondrial and nuclear DNA defects in Saccharomyces cerevisiae with mutations in DNA polymerase γ associated with progressive external ophthalmoplegia. Hum. Mol. Genet. 2006, 15, 363–374. [Google Scholar] [CrossRef]
- Chu, Z.; Li, J.; Eshaghi, M.; Karuturi, R.K.M.; Lin, K.; Liu, J. Adaptive expression responses in the Pol-γ null strain of S. pombe depleted of mitochondrial genome. BMC Genomics 2007, 8, 323. [Google Scholar] [CrossRef]
- Dujon, B. Mitochondrial genetics revisited. Yeast 2020, 37, 191–205. [Google Scholar] [CrossRef]
- Kalifa, L.; Sia, E.A. Analysis of Rev1p and Pol ζ in mitochondrial mutagenesis suggests an alternative pathway of damage tolerance. DNA Repair 2007, 6, 1732–1739. [Google Scholar] [CrossRef]
- Baldacci, G.; Bernardi, G. Replication origins are associated with transcription initiation sequences in the mitochondrial genome of yeast. EMBO J. 1982, 1, 987. [Google Scholar] [CrossRef]
- de Zamaroczy, M.; Faugeron-Fonty, G.; Baldacci, G.; Goursot, R.; Bernardi, G. The ori sequences of the mitochondrial genome of a wild-type yeast strain: Number, location, orientation and structure. Gene 1984, 32, 439–457. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Nandakumar, D.; Deshpande, A.P.; Lucas, T.P.; Ramanagouda, R.; Tang, G.Q.; Raney, K.; Yin, Y.W.; Patel, S.S. The yeast mitochondrial RNA polymerase and transcription factor complex catalyzes efficient priming of DNA synthesis on single-stranded DNA. J. Biol. Chem. 2016, 291, 16828–16839. [Google Scholar] [CrossRef] [PubMed]
- Baldacci, G.; Cherif-Zahar, B.; Bernardi, G. The initiation of DNA replication in the mitochondrial genome of yeast. EMBO J. 1984, 3, 2115. [Google Scholar] [CrossRef]
- Graves, T.; Dante, M.; Eisenhour, L.; Christianson, T.W. Precise mapping and characterization of the RNA primers of DNA replication for a yeast hypersuppressive petite by in vitro capping with guanylyltransferase. Nucleic Acids Res. 1998, 26, 1309–1316. [Google Scholar] [CrossRef] [PubMed]
- Paratkar, S.; Patel, S.S. Mitochondrial Transcription Factor Mtf1 Traps the Unwound Non-template Strand to Facilitate Open Complex Formation. J. Biol. Chem. 2010, 285, 3949–3956. [Google Scholar] [CrossRef]
- Gerhold, J.M.; Aun, A.; Sedman, T.; Jõers, P.; Sedman, J. Strand invasion structures in the inverted repeat of Candida albicans mitochondrial DNA reveal a role for homologous recombination in replication. Mol. Cell 2010, 39, 851–861. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Name | Sequence |
|---|---|
| 5UTR-MIP1[ApaI]-F | CGCGGGGCCCTTCACATTGACATGTCATTAATGACTGCC |
| 5UTR-MIP1[SpeI]-R | CGCGACTAGTCAAGAGTTGAACATCAAGTTTGAGCTGTC |
| HYGrB-pAG32-[XbaI]-F | CGCGTCTAGAGACATGGAGGCCCAGAATACCCTC |
| HYGrB-pAG32-[SpeI]-R | CGCGACTAGTCAGTATAGCGACCAGCATTCACATAC |
| alc-prom-[SpeI]-F | CGCGACTAGTCAATGAAGCCCATTCATCTTCTTGTCGACGAGC |
| alc-prom-[SpeI]-R | CGCGACTAGTTTGTCGGCTGTTTCGTGGCAAGTCGTG |
| MTS-MIP1[SpeI]-[5]F | CGCGACTAGTATGAATACGCTTTGTCCTGCTGCTGGTCACG |
| MTS-MIP1[NotI]-[3]R | CGCGGCGGCCGCCATACTCGTATTCCGACGACAGTGTGCC |
| β-actin-F | CATACATGGTCGAGAACAAGTCC |
| β-actin-R | AGTCCAGCGCCCCAAATAAC |
| mip1-F | AGGAGCGGGTGGTTGTTG |
| mip1-R | TCGCGATTCTTCTTGTGCC |
| atp9-F | ATGTTACAATCTTCAAAAATAATAGGAGC |
| atp9-R | TTAAGCAACATTTAATAATAATAATGACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varassas, S.P.; Amillis, S.; Pappas, K.M.; Kouvelis, V.N. The Identification of the Mitochondrial DNA Polymerase γ (Mip1) of the Entomopathogenic Fungus Metarhizium brunneum. Microorganisms 2024, 12, 1052. https://doi.org/10.3390/microorganisms12061052
Varassas SP, Amillis S, Pappas KM, Kouvelis VN. The Identification of the Mitochondrial DNA Polymerase γ (Mip1) of the Entomopathogenic Fungus Metarhizium brunneum. Microorganisms. 2024; 12(6):1052. https://doi.org/10.3390/microorganisms12061052
Chicago/Turabian StyleVarassas, Stylianos P., Sotiris Amillis, Katherine M. Pappas, and Vassili N. Kouvelis. 2024. "The Identification of the Mitochondrial DNA Polymerase γ (Mip1) of the Entomopathogenic Fungus Metarhizium brunneum" Microorganisms 12, no. 6: 1052. https://doi.org/10.3390/microorganisms12061052
APA StyleVarassas, S. P., Amillis, S., Pappas, K. M., & Kouvelis, V. N. (2024). The Identification of the Mitochondrial DNA Polymerase γ (Mip1) of the Entomopathogenic Fungus Metarhizium brunneum. Microorganisms, 12(6), 1052. https://doi.org/10.3390/microorganisms12061052